1. Introduction
Mycotoxins are toxic fungal metabolites that can contaminate a wide array of food and feed [1]. Mycotoxin-producing fungi can be classified into either field or storage fungi. Field fungi, such as the Fusarium species, produce mycotoxins on the crops in the field, whereas storage fungi, such as the Aspergillus and Penicillium species, produce mycotoxins on the crops after harvesting [2]. Fusarium fungi have traditionally been associated with temperate climatic conditions, since they require somewhat lower temperature for growth and mycotoxin production than, for example, the Aspergillus species [3]. The most toxicologically important Fusarium mycotoxins are trichothecenes (including deoxynivalenol (DON) and T-2 toxin (T-2)), zearalenone (ZEN) and fumonisin B1 (FB1).
Fusarium mycotoxins are capable of inducing both acute and chronic toxic effects. These effects are dependent on the mycotoxin type, the level and duration of exposure, the animal species that is exposed and the age of the animal [4]. Intake of high doses of mycotoxins may lead to acute mycotoxicoses, which are characterized by well-described clinical signs [5,6]. Exposure of pigs to high concentrations of DON causes abdominal distress, malaise, diarrhea, emesis and even shock or death. Exposure of pigs to fumonisins can lead to pulmonary edema due to cardiac insufficiency. In horses fumonisins can cause equine leukoencephalomalacia (ELEM) and target the brain [7]. Since these high contamination levels are rare in modern agricultural practice [8], this review will not discuss extensively their effect on animal or human health. Indeed, although the results of a global survey indicate that the Fusarium mycotoxins DON, fumonisins, and ZEN respectively contaminated 55%, 54% and 36% of feed and feed ingredients in the period 2004–2011, the majority of samples was found to comply with even the most stringent European Union regulations or recommendations on the maximal tolerable concentration (Table A3) [8]. Therefore, this review will focus on the effect of low to moderate doses of the major Fusarium mycotoxins.
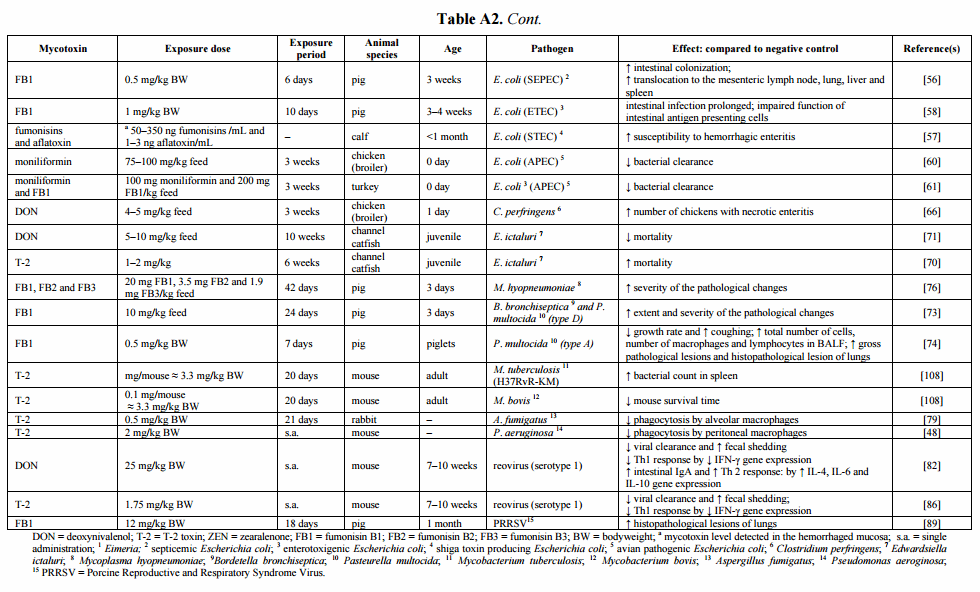
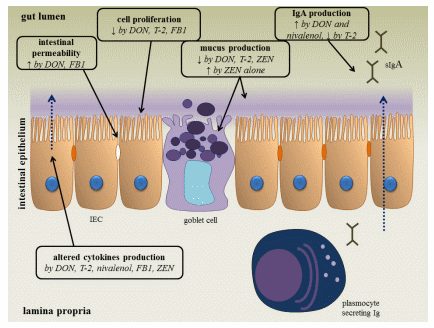
Following oral intake of low to moderate amounts of these mycotoxins, the gastro-intestinal epithelial cell layer will be exposed first [9]. The intestinal mucosa acts as a barrier, preventing the entry of foreign antigens including food proteins, xenobiotics (such as drugs and toxins), commensal microbiota and pathogens into the underlying tissues [9,10]. The mucosal immunity, which consists of an innate and adaptive immune system, can be affected by Fusarium mycotoxins (Figure 1) [9,10]. An important component of the innate immune system are the intestinal epithelial cells, which are interconnected by tight junctions, and covered with mucus, produced by goblet cells [11]. By measuring the transepithelial electrical resistance (TEER), several in vitro and ex vivo studies indicate that DON and FB1 are able to increase the permeability of the intestinal epithelial layer of human, porcine and avian origin [12–14]. Also the viability and proliferation of animal and human intestinal epithelial cells can be negatively affected by Fusarium mycotoxins [9,15–20]. Their effect on mucus production is variable: co-exposure of low doses of DON, T-2 and ZEN reduces the number of goblet cells in pigs [21], but ZEN given alone at higher doses increases the activity of goblet cells [22]. Several mycotoxins are also able to modulate the production of cytokines in vitro and in vivo [9,23]. For example, DON increases the expression of TGF-β and IFN-γ in mice and fumonisins decrease the expression of IL-8 in an intestinal porcine epithelial cell line (IPEC-1) [9].
Figure 1. The effect of Fusarium mycotoxins on the intestinal epithelium. A variety of Fusarium mycotoxins alter the different intestinal defense mechanisms including epithelial integrity, cell proliferation, mucus layer, immunoglobulins (Ig) and cytokine production. (IEC: intestinal epithelial cell) (based on [9]).
Fusarium mycotoxins can cross the intestinal epithelium and reach the systemic compartment [20,24], affecting the immune system. Exposure to these toxins can either result in immunostimulatory or immunosuppressive effects depending on the age of the host and exposure dose and duration [20,25]. Mycotoxin-induced immunomodulation may affect innate and adaptive immunity by an impaired function of macrophages and neutrophils, a decreased T- and B-lymphocyte activity and antibody production [23,25,26]. In addition to the effect of Fusarium mycotoxins on the animal or human host, these mycotoxins may alter the metabolism of the pathogen, which may alter the outcome of the infectious disease [27,28].
A wealth of research papers clearly indicate a negative influence of Fusarium mycotoxins on the intestinal function and immune system. Since the intestinal tract is also a major portal of entry to many enteric pathogens and their toxins, mycotoxin exposure could increase the animal susceptibility to these pathogens. Furthermore, mycotoxin-induced immunosuppression may also result in decreased animal or human host resistance to infectious diseases.
This review attempts to summarize the impact of Fusarium mycotoxin exposure on the animal and human host susceptibility to infectious diseases. More specifically, the effect of Fusarium mycotoxins on enteric, systemic and respiratory infectious diseases in livestock animals and animal models for human diseases are highlighted.
2. Effect of Fusarium Toxins on Parasitic Diseases
Coccidiosis
Intestinal protozoa, including the coccidia (Eimeria, Isospora, Cryptosporidium and Sarcosporidia) and flagellates, are important infectious agents. Coccidiosis in poultry generally refers to the disease caused by the Eimeria species, and is still considered one of the most important enteric diseases affecting performance. These obligate intracellular parasites have an oral-fecal life cycle with developmental stages alternating between the external environment and the host [29].
Seven species of Eimeria (E. acervulina, E. brunetti, E. maxima, E. mitis, E. necatrix, E. praecox and E. tenella) are found in chickens [29]. The physical and biological characteristics, pathogenicity and immunogenicity depend on the species. Immunity to Eimeria is complex, multifactorial and influenced by both host and parasite [30].
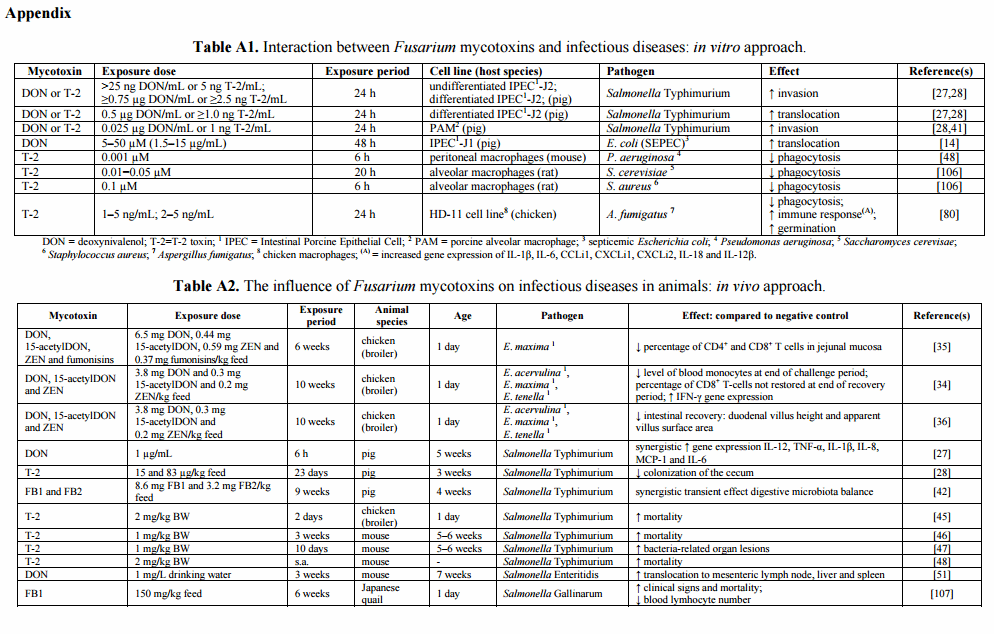
Cell-mediated immunity, mainly evoked by the intraepithelial lymphocytes (IEL) and lymphocytes of the lamina propria, is the major protective immune component against avian coccidiosis [31,32]. The CD4+ T-lymphocytes, IEL and macrophages are involved in the response against primary exposure to Eimeria [31], while CD8+ T-lymphocytes and IFN-γ are important in the protective immune response against Eimeria infection [33]. Girgis et al. [34,35] showed a negative impact of diets naturally contaminated with Fusarium mycotoxins on the cell-mediated immune response against coccidiosis in broilers (Table A2). Following primary infection of broilers with Eimeria, Fusarium mycotoxins decreased the percentage of CD4+ and CD8+ T-cells in the jejunal mucosa [35]. In addition, feeding on a mycotoxin-contaminated diet lowered the blood levels of CD8+ T-cells and monocytes, which could suggest an increased recruitment at the intestinal site of coccidial infection or a delayed replication necessary to replenish these subsets in the circulation [34,35]. Additionally, feeding on a Fusarium mycotoxin-contaminated diet increased IFN-γ gene expression in the cecal tonsils of Eimeria-challenged birds, however, without being linked to the apparent resistance to coccidial infection in terms of changes in oocyst yield [34]. The cecal tonsils constitute a lymphoid tissue in the cecum belonging to the gut-associated lymphoid tissue (GALT). Resistance to Eimeria infection is related to the expression of a set of interleukins rather than only IFN-γ and the up-regulation of the gene may not necessarily be associated with functional secretion [34]. Furthermore, it was shown that moderate levels of Fusarium mycotoxins negatively affect intestinal morphology and interfere with intestinal recovery from an enteric coccidial infection, indicated by a lower villus height and apparent villus area (Table A2) [36]. Although Girgis et al. [34,35] demonstrated that Fusarium mycotoxins impair the Eimeria-induced immune response, no effect was seen on fecal oocyst counts. Similarly, Békési et al. [37] showed no impact of a T-2 and ZEN-contaminated diet on Cryptosporidium baileyi oocyst excretion in broilers.
Research investigating the influence of mycotoxins on the animal susceptibility to infectious diseases focuses mainly on exposure to single major mycotoxins. Limited information about the impact of mycotoxin co-occurrence and plant metabolites of mycotoxins on this interaction is available. Nevertheless, Girgis et al. [34,35] showed that the combination of DON, 15-acetylDON (15-AcDON), ZEN and fumonisins alters the Eimeria-induced immune response. Interestingly, mycotoxin contamination of broiler feed may reduce the efficacy of the anti-coccidial treatment with lasalocid [38].
To conclude, Fusarium mycotoxins negatively affect the innate and adaptive cellular immune response against Eimeria, though without changing the oocyst yield. Further data of clinical coccidiosis lesion scoring is still needed in order to evaluate the effect of Fusarium mycotoxins on the severity of the disease.
3. Effect of Fusarium Toxins on Bacterial Diseases
3.1. Salmonellosis
Salmonellosis is an infection with the Gram-negative Salmonella bacterium, a facultative anaerobic, facultative intracellular microorganism of the Enterobacteriaceae family. The host—Salmonella interaction is complex, with a broad array of mechanisms used by the bacteria to overcome host defenses. Two important disease manifestations are differentiated, i.e., gastroenteritis and enteric fever, caused by nontyphoidal and typhoidal Salmonella serovars, respectively [39].
Nontyphoidal Salmonella strains, such as Salmonella serovar Typhimurium and Salmonella serovar Enteritidis strains, infect a wide range of animal hosts, including pigs and poultry, without causing clinical symptoms in these animals. Infection in slaughter pigs and poultry can cause meat and egg contamination [39,40].
An infection with Salmonella generally occurs in three stages: the adhesion to the intestinal wall, the invasion of the gut wall and the dissemination to mesenteric lymph nodes and other organs. Via bacterial-mediated endocytosis, Salmonella invades the intestinal epithelial cells, after which the bacterium becomes enclosed within an intracellular phagosomal compartment (the Salmonella-containing vacuole (SCV)). After crossing the epithelial barrier, the bacterium is located predominantly in macrophages in the underlying tissue [39].
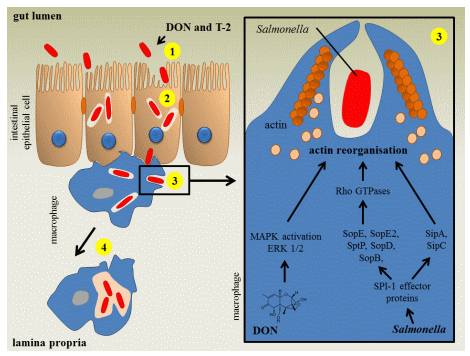
Feeding pigs a Fusarium mycotoxin-contaminated diet influences the intestinal phase of the pathogenesis of Salmonella Typhimurium infections as illustrated in Figure 2. Non-cytotoxic concentrations of DON and T-2 enhance intestinal Salmonella invasion and increase the passage of Salmonella Typhimurium across the epithelium (Table A1) [28,41]. Chronic exposure of specific pathogen-free pigs to naturally fumonisin-contaminated feed had no impact on Salmonella Typhimurium translocation [42]. Once Salmonella has invaded the intestinal epithelium, the innate immune system is triggered and the porcine gut will start to produce several cytokines [28,43]. Both Fusarium mycotoxins and Salmonella affect the innate immune system. Vandenbroucke et al. [27] showed that low concentrations of DON could potentiate the early intestinal immune response induced by Salmonella Typhimurium infection. Co-exposure of the intestine to DON and Salmonella Typhimurium resulted in increased expression of several cytokines, for instance, those responsible for the stimulation of the inflammatory response (TNF-α) and T-lymphocyte stimulation (IL-12) (Table A2). The authors suggested that the enhanced intestinal inflammation could be due to a DON-induced stimulation of Salmonella Typhimurium invasion in and translocation across the intestinal epithelium [27].
Fusarium mycotoxins also affect the systemic part of the Salmonella Typhimurium infection in pigs. After the intestinal phase of the pathogenesis, Salmonella can spread to the bloodstream using the host macrophage to establish the systemic infection. However, in pigs the systemic part of Salmonella Typhimurium is poorly documented and colonization is mostly limited to the gastrointestinal tract [44]. After bacterial uptake by the macrophage, Salmonella can survive and even proliferate in this cell. Exposure of macrophages to non-cytotoxic concentrations of DON and T-2 promotes the uptake of Salmonella Typhimurium (Figure 2, Table A1). Salmonella entry in host cells involves a complex series of actin cytoskeletal changes. Macrophage invasion coincides with membrane ruffling, followed by bacterium uptake and formation of Salmonella-containing vacuole [41]. Vandenbroucke et al. [41] showed in vitro that DON enhances Salmonella Typhimurium engulfment, since low concentrations of DON modulate the cytoskeleton of macrophages through ERK1/2 F-actin reorganization resulting in an enhanced uptake of Salmonella Typhimurium in porcine alveolar macrophages (PAM) (Figure 2, Table A1). Non-cytotoxic concentrations of the Fusarium mycotoxins DON and T-2 did not affect the intracellular proliferation of Salmonella Typhimurium in porcine macrophages (Figure 2) [28,41].
Figure 2. The impact of deoxynivalenol and T-2 toxin on a Salmonella Typhimurium infection in pigs. In vitro, deoxynivalenol (DON) and T-2 toxin (T-2) promote Salmonella invasion (1) and transepithelial passage (2) of IPEC-J2 cell layer. Subsequently, the bacterium can spread to the bloodstream using the host macrophage to establish the systemic infection. In vitro, DON and T-2 enhance Salmonella uptake (3) in porcine alveolar macrophages. The Salmonella invasion of macrophages coincides with membrane ruffling, caused by actin cytoskeletal changes. Activation of host Rho GTPases by the Salmonella pathogenicity island (SPI)-1 type 3 secretion system (T3SS) effector proteins SopB, SopE, SopE2 and SopD leads to actin cytoskeleton reorganization. After Salmonella internalization has occurred, the bacterium injects the effector protein SptP which promotes the inactivation of Rho GTPases. The bacterium can also modulate the actin dynamics of the host cell in a direct manner through the bacterial effector proteins SipA and SipC. The mycotoxin DON enhances the uptake of Salmonella in macrophages through activation of the mitogen-activated protein kinases (MAPK) extracellular signal-regulated kinases (ERK1/2) pathway, which induces actin reorganizations and membrane ruffles. DON and T-2 do not affect intracellular bacterial proliferation (4) (based on [41,44]).
In addition to the effects of Fusarium mycotoxins on the host susceptibility to a Salmonella Typhimurium infection, these mycotoxins also modulate the bacterial metabolism. Although no effect of DON or T-2 on the growth of Salmonella Typhimurium is detected, DON and T-2 modulate the Salmonella gene expression [28,41]. The enhanced inflammatory effect following exposure to DON is more likely a result of the toxic effect of the mycotoxin on the intestine than on the bacterium [27].
Only high concentrations of DON increase the bacterial expression of regulators of Salmonella pathogenicity island (SPI)-1 and SPI-2, respectively hilA and ssrA. SPI-1 consists of genes coding for bacterial secretion systems necessary for invasion, while SPI-2 genes encode essential intracellular replication mechanisms [41]. For T-2 the toxic effects on the bacterium itself are probably more pronounced than the host cell-mediated effects resulting in a reduced in vivo colonization in pigs. Low concentrations of T-2 cause a reduced motility of Salmonella and a general down regulation of genes involved in Salmonella metabolism, genes encoding ribosomal proteins and SPI-1 genes [28].
Only limited information is available concerning the interaction between Fusarium mycotoxins and Salmonella Typhimurium infection in other animals. The currently available publications mainly focus on the interaction of T-2 and the systemic phase of a Salmonella Typhimurium infection. In T-2-challenged broiler chickens and mice an increased level of Salmonella Typhimurium-related organ lesions or mortality was seen (Table A2) [45–48]. Infection of mice with Salmonella Typhimurium results in systemic infection and a disease similar to that seen in humans after infection with Salmonella Typhi [49]. Increased mortality might be explained partly by the synergistic effects of bacterial lipopolysaccharide (LPS) and T-2 during the late phase of murine salmonellosis [50]. In addition to Salmonella Typhimurium, DON reduces the resistance to oral infection with Salmonella Enteritidis in mice by promoting translocation of Salmonella to mesenteric lymph node (MLN), liver and spleen (Table A2) [51].
Mouse and pig models are important animal models to investigate the impact of mycotoxins, infectious diseases and their combination on animal health [52,53]. Infection of mice with Salmonella Typhimurium is an important host–pathogen interaction model to investigate typhoid fever in humans. Moderate to high concentrations of T-2 have shown to increase Salmonella-induced mortality [46,47,50]. The pig is very similar to humans in terms of anatomic and physiologic characteristics such as size, digestive physiology, kidney structure and function, pulmonary vascular bed structure, coronary artery distribution, respiratory rates, cardiovascular anatomy and physiology, and immune response, and has been used to study various intestinal pathogens, including Salmonella and Escherichia coli [53]. The interaction between mycotoxins and Salmonella Typhimurium studied in a porcine model of infection, gives us relevant information concerning the impact of this interaction on human intestinal inflammation and immune response [27].
In conclusion, the exact outcome of co-exposure to Fusarium mycotoxins and Salmonella Typhimurium is difficult to predict. Published data show an influence of mycotoxin exposure on the bacterium, the host cells and the host–pathogen interaction. Depending on the characteristics of the mycotoxin exposure, one of these effects will determine the outcome of the interaction between Fusarium mycotoxins and Salmonella Typhimurium.
3.2. Colibacillosis
Escherichia coli is a Gram-negative, non-sporulating rod-shaped bacterium of the family Enterobacteriaceae. Although this bacterium is considered to be a normal component of the intestinal microbiota, it is frequently associated with both intestinal and extra-intestinal infections in humans and animals. A certain number of these strains possess particular combinations of virulence factors which enables them to cause disease. Clinical syndromes resulting from infection with these pathotypes include enteric/diarrheal disease, urinary tract infections and sepsis/meningitis.
The pathogenesis of E. coli infections depends on the pathotype involved and may include colonizing the intestinal mucosa, evasion of host defenses, multiplication, and induction of host damage [54,55].
Fusarium mycotoxins may influence the pathogenesis of E. coli infections in different animal species by stimulating intestinal colonization and translocation and negatively affecting the immune response. Feeding a diet contaminated with a moderate level of FB1 to pigs enhanced intestinal colonization and translocation of a septicemic E. coli (SEPEC) strain from the intestine to the systemic compartment. FB1-treatment resulted in a higher bacterial translocation to the mesenteric lymph nodes and lungs, and to a lesser extent to liver and spleen (Table A2) [56]. It was shown in vitro that DON increased the translocation of SEPEC over the intestinal epithelial cell monolayer (IPEC-1) (Table A1) [14].
Mycotoxins increase the calf susceptibility to shiga toxin or verotoxin-producing E. coli (STEC)-associated hemorrhagic enteritis. Recently, Baines et al. [57] showed that exposing calves of less than one month old to the combination of aflatoxin and fumonisins promoted STEC-associated hemorrhagic enteritis (Table A2) [57].
Feeding a FB1-contaminated diet to pigs negatively affects the mucosal immune response against an infection with enterotoxigenic E. coli (ETEC). Devriendt et al. [58] showed a prolonged intestinal infection of E. coli in pigs administered fumonisins for 10 consecutive days and subsequently challenged with E. coli (F4+ ETEC) (Table A2). Antigen-presenting cells (APCs) have an important role in the mucosal immune system by connecting the innate and adaptive immune response, through uptake of antigen in lamina propria, maturation and migration to GALT, and interaction with T cells. FB1 negatively affected the function of intestinal APCs by a reduced up-regulation of the major histocompatibility complex class II (MHC-II), cluster of differentiation (CD) 80/6 and IL-12p40 cytokine gene expression [58]. This altered function of APCs could therefore influence the E. coli-induced adaptive immune response [58,59]. Additionally, moniliformin and FB1 delayed systemic E. coli (avian pathogenic E. coli, APEC) clearance in broilers and turkeys after intravenous administration (Table A2) [60,61].
The results of these studies may also be valid for human infections since the gastro-intestinal tract of pigs and humans are very similar [58]. Infant diarrhea caused by enteropathogenic E. coli (EPEC) is known to be of major concern in developing countries and, for instance, enterohemorrhagic E. coli (EHEC) infections are a major worldwide public health hazard.
3.3. Necrotic Enteritis in Broilers
Necrotic enteritis (NE) is a disease in broilers caused by Clostridium perfringens. This Gram-positive spore-forming bacterium occurs naturally in the environment, feed and gastrointestinal tract of chickens and other animals [62,63]. NE is a complex, multifactorial enteric disease with many known and unknown factors influencing its occurrence and the severity of the outbreaks. The best-known predisposing factor is mucosal damage caused by coccidial pathogens [64]. Only C. perfringens strains expressing the NetB toxin are capable of inducing NE in broilers [65]. C. perfringens is auxotrophic for several amino acids, thus availability of these amino acids would allow extensive bacterial proliferation [63].
The intake of DON-contaminated feed is a predisposing factor for the development of necrotic enteritis in broiler chickens due to the negative influence on the epithelial barrier, and to an increased intestinal nutrient availability for clostridial proliferation. Recently, we [66] showed in an experimental subclinical NE infection model that chickens fed a diet contaminated with DON for three weeks were more prone to develop NE lesions compared to chickens on a control diet (Table A2). The negative effects of DON on the small intestinal barrier can lead to an impaired nutrient digestion and leakage of plasma amino acids into the intestinal lumen, providing the necessary growth substrate for extensive proliferation of C. perfringens [66].
3.4. Edwardsiella ictaluri Infection in Catfish
Edwardsiella ictaluri is a Gram-negative bacterium of the Enterobacteriaceae family. Bacillary Necrosis of Pangasianodon (BNP) caused by E. ictaluri is the most frequently occurring infectious disease in catfish [67]. Besides the Vietnamese freshwater production, also the American channel catfish (Ictalurus punctatus) industry suffers massively from E. icatluri infections which have been termed Enteric Septicemia of Catfish (ESC). BNP is characterized by multifocal irregular white spots of varying sizes on several organs including liver, spleen and kidney [68]. ESC in channel catfish may occur in an acute form characterized by enteritis and septicemia with rapid mortality, or in a chronic form, which is characterized by meningoencephalitis, open lesions on the cranial region and exophthalmia [69]. Mortality associated with the co-occurrence of Fusarium mycotoxins and E. ictaluri is difficult to predict in juvenile channel catfish. T-2 increased E. ictaluri-associated mortality [70], while moderate contamination of DON improved the survival of the channel catfish [71] (Table A2). Mycotoxin sensitivity differs between fish species. Rainbow trout, for example, are extremely sensitive to DON, while channel catfish are rather resistant [71,72]. Important data concerning the toxicity of the mycotoxin on the bacterium are lacking. Further investigation of the interaction between Fusarium mycotoxins and E. ictaluri will be necessary to evaluate the outcome.
3.5. Swine Respiratory Disease
Respiratory disease in pigs is often caused by the combined effects of multiple pathogens and predisposing factors [73]. Primary infections with bacteria such as Actinobacillus pleuropneumoniae, Mycoplasma hyopneumoniae, Bordetella bronchiseptica or viruses such as influenza virus and Porcine Reproductive and Respiratory Syndrome Virus (PRRSV), can predispose pigs to secondary pathogens such as Pasteurella multocida and Trueperella pyogenes [74]. Respiratory symptoms can vary depending on the pathogens involved. M. hyopneumoniae is the principal etiological agent responsible for enzootic pneumonia in pigs [75]. M. hyopneumoniae is an obligate symbiotic and host-specific bacterium, which is lacking a cell wall. This pathogen affects the respiratory mucosal clearance system by disrupting the celia on the epithelial surface and modulates the immune system of the respiratory tract. Consequently, M. hyopneumoniae predisposes animals to concurrent infections with other respiratory pathogens [75]. Dietary exposure to fumonisins induces pulmonary edema and may facilitate M. hyopneumoniae infection (Table A2) [76].
The progressive form of porcine atrophic rhinitis is often due to a combined infection with B. bronchiseptica and toxigenic P. multocida [73,77]. Dietary exposure to FB1 of piglets infected with both bacteria increases the risk of pneumonia and the severity of the pathological changes [73]. P. multocida type A is the most frequently occurring secondary pathogen that can cause pneumonic pasteurellosis [78]. Halloy et al. [74] showed that inoculation of piglets with P. multocida combined with an oral bolus of FB1 induced a cough and a lung inflammatory process characterized by an increased number of total cells, macrophages and lymphocytes in broncheo-alveolar lavage fluid (BALF). Lung lesions were more severe in these animals and consisted of subacute interstitial pneumonia [74].
4. Effect of Fusarium Toxins on Fungal Diseases
Aspergillosis
Aspergillus fumigatus is an ubiquitous saprophytic fungus found in soil, plant debris, and the indoor environment, including hospitals. This fungus is also an opportunistic pathogen. Inhalation of its conidia can cause life-threatening infections in the respiratory system of immunocompromised animals and humans. Respiratory macrophages are the first line of defense against inhaled Aspergillus conidia. T-2 impaired the phagocytotic activities of macrophages against A. fumigatus conidia in chickens and rabbits (Tables A1 and A2) [79,80]. However, the pro-inflammatory response of A. fumigatus infected chicken macrophages was increased by T-2 (Table A1) [80]. The effect of T-2 on the innate immune response against Aspergillus conidia is dual, which suggests that depending on the characteristics of the mycotoxin exposure and the animal, one of these effects will determine the outcome of this interaction.
5. Effect of Fusarium Toxins on Viral Diseases
5.1. Reovirus
Reovirus is a non-enveloped double-stranded RNA virus that has been isolated from the gastro-intestinal tract and respiratory tract of both humans and animals [81,82]. Enteric reoviruses cause mostly a mild and self-limiting infection [82]. Nevertheless, reovirus infections can be more severe, affecting, for example, the central nervous system in mice and rats [81]. Viral arthritis is the most frequent reovirus-associated disease in poultry, which is characterized by lameness and swellings affecting primarily tarsometatarsal joints and the feet [83–85].
Fusarium mycotoxins negatively affect the intestinal virus clearance in mice. Li et al. [82,86] showed that high concentrations of DON and T-2 suppress the host immune response to reovirus as evidenced by the inability to clear the virus from the intestine as well as by increased fecal shedding of the virus (Table A2). Trichothecene exposure increased the intestinal viral load, which could increase inflammation and discomfort to the host during the infection process. The increased fecal shedding could enhance virus dissemination among individuals [86]. Both mycotoxins decreased the cell-mediated viral clearance by suppressing the gene expression of IFN-γ in Peyer’s Patches (PP) [82,86]. DON enhanced Th2 cytokine expression prior to and after reovirus infection, which potentiates the IgA and IgG responses to reovirus [82]. In contrast, T-2 suppressed reovirus-induced immunoglobulin responses [86]. The lack of a similar effect of Th2 cytokines by T-2 suggests inherent differences between both mycotoxins in their capacity to modulate cytokines during viral infection, although both mycotoxins belong to the class of trichothecenes [86]. Nevertheless, the intestinal clearance of reovirus was less efficient after T-2 exposure compared to DON [86]. Since reovirus infection in mice is used as a model for several enteric and respiratory viral infections in humans and other animals [81], these results could assume an impact of mycotoxins on host susceptibility to more virulent viruses.
5.2. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV)
Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) is an enveloped single-stranded RNA virus belonging to the family Arteriviridae, within the order Nidovirales [87]. Currently, PRRS is one of the most economic significant diseases in swine production [87,88]. The clinical symptoms, respiratory or reproductive, vary with the viral strain, the immune status of the herd, and management factors [88,89]. PRRSV is a highly infectious virus that replicates within the monocytes or macrophages with the lung being a predominant site of viral multiplication [89]. Exposure of piglets to FB1 increased the risk for PRRSV disease [90]. More severe histopathological lesions were observed when pigs were exposed to FB1 and subsequently inoculated with PRRSV. The authors suggest that FB1 causes immunosuppression, facilitating PRRSV to induce more severe lesions [89]. Given the importance of PRRSV in worldwide swine production and the frequent occurrence of fumonisins, research should be performed investigating this interaction also at lower doses of FB1.
6. Discussion
In recent years, research investigating the effects of Fusarium mycotoxins on the intestinal and immune functions has made substantial progress. However, only limited information is available on the interaction between mycotoxins and infectious diseases. The aforementioned literature data indicates that Fusarium mycotoxins may influence the animal and human host susceptibility to enteric, systemic and respiratory infectious diseases. Depending on host, pathogen and mycotoxin characteristics, exposure to Fusarium mycotoxins can generally exacerbate infectious diseases. On the other hand, T-2 has been shown to decrease the colonization capacity of Salmonella in the pig intestine. Fusarium mycotoxins may influence the host–pathogen interaction by negatively affecting the intestinal barrier function and the innate and adaptive immune response [9,23,26]. Fusarium mycotoxins affect the morphology and the barrier function of the intestinal layer [9], leading to increased translocation of different bacterial species including Salmonella enterica and E. coli, to the systemic compartment. The negative influence of these mycotoxins on the function of macrophages results in impaired phagocytosis of bacterial and fungal pathogens. However, also the adaptive immune response is targeted, demonstrated by the effect on gene expression of several cytokines, leading to an altered Th1 and Th2 response.
The economic impact of mycotoxins on animal production is generally considered to be mainly due to losses related to direct effects on animal health and trade losses related to grain rejection [91]. It is clear, however, that the indirect influence of myocotoxins on animal health, by enhancing infectious diseases, should also be taken into account. These effects, as reviewed here, occur even at low to moderate mycotoxin contamination levels of feed [8]. Some publications showed that these effects can even occur at contamination levels below the European guidance levels, suggesting that the legislation may not cover all deleterious health effects of mycotoxins.
Fusarium mycotoxins have various acute and chronic effects on humans [92]. DON could play a role in diseases such as inflammatory bowel disease (IBD) [20,93]. Taken into account conditions such as environmental, socio-economic and food production, it seems plausible that the risk for food-associated mycotoxin exposure is even higher in developing countries [94]. Besides the risk for acute mycotoxicosis in developing countries [95], results obtained in animals suggest that low to moderate concentrations of these mycotoxins could also influence human susceptibility to infectious diseases.
The effect of multi-mycotoxin contamination and of less well-known or emerging mycotoxins on the human or animal susceptibility to infectious diseases is rather unknown. Multi-mycotoxin contamination of feed is frequently occurring, raising the question on the impact on animal toxicity of this phenomenon [3]. Several in vitro and in vivo studies demonstrated an enhanced toxicity and more severe immune suppression compared to single mycotoxin contamination [96–98]. In addition, plant metabolites of mycotoxins may also be present in feed and are known as masked mycotoxins [99]. Fusarium fungi and infected plants may produce conjugated forms of, for instance, DON, such as 3-AcDON (3-acetylDON), 15-AcDON and DON-3G (DON-3-glucoside). Furthermore, mycotoxins can also be conjugated by certain food-processing techniques. These conjugated forms could have a direct toxic effect, or may be hydrolyzed to their precursor mycotoxin in the digestive tract of animals, resulting in higher exposure levels [100–102]. The influence of mycotoxin co-occurrence and masked mycotoxins on human and animal susceptibility to infectious diseases will be an important research question in the future.
Global warming and increasing world population of humans are further important issues. Climate changes may affect the global distribution of mycotoxigenic fungi and their mycotoxins [103,104], but also the distribution of infectious diseases [105]. Livestock farming will remain an important component of the global food supply in the future. Animal health, including the impact of mycotoxins and susceptibility to infectious diseases, will be important future topics to produce enough safe food for the entire human population.
In conclusion, Fusarium mycotoxins may alter the human and animal susceptibility to infectious diseases by affecting the intestinal health and the innate and adaptive immune system. Further research will be necessary to investigate the impact of mycotoxins on infectious diseases and to develop practical, economically justified, solutions to counteract mycotoxin contamination of feed and food, and its effects on human and animal health.
Acknowledgments
G. Antonissen was supported by a PhD fellowship from Biomin GmbH, Herzogenburg, Austria.
Conflicts of Interest
The authors declare no conflict of interest.
This article was originally published in Toxins 2014, 6, 430-452; doi:10.3390/toxins6020430. This is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
 Click here to enlarge the image
Click here to enlarge the image
References
1. Binder, E.M. Managing the risk of mycotoxins in modern feed production. Anim. Feed Sci. Tech. 2007, 133, 149–166.
2. Filtenborg, O.; Frisvad, J.C.; Thrane, U. Moulds in food spoilage. Int. J. Food Microbiol. 1996, 33, 85–102.
3. Placinta, C.; D’mello, J.; Macdonald, A. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Tech. 1999, 78, 21–37.
4. D’mello, J.; Placinta, C.; Macdonald, A. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Tech. 1999, 80, 183–205.
5. Smith, T.K.; Diaz, G.; Swamy, H. Current Concepts in Mycotoxicoses in Swine. In The Mycotoxin Blue Book; Diaz, D.E. Ed.; Nottingham University Press: Nottingham, UK, 2005; pp. 235–248.
6. Devegowda, G.; Murthy, T. Mycotoxins: Their Effects in Poultry and Some Practical Solutions. In The Mycotoxin Blue Book; Diaz, D.E. Ed.; Nottingham University Press: Nottingham, UK, 2005; pp. 25–56.
7. Devreese, M.; de Backer, P.; Croubels, S. Overview of the most important mycotoxins for the pig and poultry husbandry. Vlaams Diergeneeskundig Tijdschrift 2013, 82, 171–180.
8. Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide-long term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899.
9. Bouhet, S.; Oswald, I.P. The effects of mycotoxins, fungal food contaminants, on the intestinal epithelial cell-derived innate immune response. Vet. Immunol. Immun. 2005, 108, 199–209.
10. Oswald, I.P. Role of intestinal epithelial cells in the innate immune defence of the pig intestine. Vet. Res. 2006, 37, 359–368.
11. Schenk, M.; Mueller, C. The mucosal immune system at the gastrointestinal barrier. Best Pract. Res. CL GA 2008, 22, 391–409.
12. Maresca, M.; Mahfoud, R.; Garmy, N.; Fantini, J. The mycotoxin deoxynivalenol affects nutrient absorption in human intestinal epithelial cells. J. Nutr. 2002, 132, 2723–2731.
13. Sergent, T.; Parys, M.; Garsou, S.; Pussemier, L.; Schneider, Y.-J.; Larondelle, Y. Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular metabolism at realistic intestinal concentrations. Toxicol. Lett. 2006, 164, 167–176.
14. Pinton, P.; Nougayrède, J.-P.; Del Rio, J.-C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.-P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharm. 2009, 237, 41–48.
15. Bouhet, S.; Hourcade, E.; Loiseau, N.; Fikry, A.; Martinez, S.; Roselli, M.; Galtier, P.; Mengheri, E.; Oswald, I.P. The mycotoxin fumonisin B1 alters the proliferation and the barrier function of porcine intestinal epithelial cells. Toxicol. Sci. 2004, 77, 165–171.
16. Awad, W.A.; Bohm, J.; Razzazi-Fazeli, E.; Zentek, J. Effects of feeding deoxynivalenol contaminated wheat on growth performance, organ weights and histological parameters of the intestine of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2006, 90, 32–37.
17. Yunus, A.W.; Blajet-Kosicka, A.; Kosicki, R.; Khan, M.Z.; Rehman, H.; Bohm, J. Deoxynivalenol as a contaminant of broiler feed: Intestinal development, absorptive functionality and metabolism of the mycotoxin. Poult. Sci. 2012, 91, 852–861.
18. Hoerr, F.; Carlton, W.; Yagen, B. Mycotoxicosis caused by a single dose of T-2 toxin or diacetoxyscirpenol in broiler chickens. Vet. Pathol. 1981, 18, 652–664.
19. Awad, W.A.; Hess, M.; Twaruzek, M.; Grajewski, J.; Kosicki, R.; Böhm, J.; Zentek, J. The impact of the fusarium mycotoxin deoxynivalenol on the health and performance of broiler chickens. Int. J. Mol. Sci. 2011, 12, 7996–8012.
20. Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820.
21. Obremski, K.; Zielonka, L.; Gajecka, M.; Jakimiuk, E.; Bakula, T.; Baranowski, M.; Gajecki, M. Histological estimation of the small intestine wall after administration of feed containing deoxynivalenol, T-2 toxin and zearalenone in the pig. Pol. J. Vet. Sci. 2008, 11, 339–345.
22. Obremski, K.; Gajecka, M.; Zielonka, L.; Jakimiuk, E.; Gajecki, M. Morphology and ultrastructure of small intestine mucosa in gilts with zearalenone mycotoxicosis. Pol. J. Vet. Sci. 2004, 8, 301–307.
23. Bondy, G.S.; Pestka, J.J. Immunomodulation by fungal toxins. J. Toxicol. Env. Heal. B 2000, 3, 109–143.
24. Osselaere, A.; Devreese, M.; Goossens, J.; Vandenbroucke, V.; de Baere, S.; de Backer, P.; Croubels, S. Toxicokinetic study and absolute oral bioavailability of deoxynivalenol, T-2 toxin and zearalenone in broiler chickens. Food Chem. Toxicol. 2012, 51, 350–355.
25. Corrier, D. Mycotoxicosis: Mechanisms of immunosuppression. Vet. Immunol. Immun. 1991, 30, 73–87.
26. Oswald, I.; Marin, D.; Bouhet, S.; Pinton, P.; Taranu, I.; Accensi, F. Immunotoxicological risk of mycotoxins for domestic animals. Food Addit. Contam. 2005, 22, 354–360.
27. Vandenbroucke, V.; Croubels, S.; Martel, A.; Verbrugghe, E.; Goossens, J.; van Deun, K.; Boyen, F.; Thompson, A.; Shearer, N.; de Backer, P. The mycotoxin deoxynivalenol potentiates intestinal inflammation by Salmonella Typhimurium in porcine ileal loops. PLoS One 2011, 6, doi: 10.1371/journal.pone.0023871.
28. Verbrugghe, E.; Vandenbroucke, V.; Dhaenens, M.; Shearer, N.; Goossens, J.; de Saeger, S.; Eeckhout, M.; D'herde, K.; Thompson, A.; Deforce, D. T-2 toxin induced Salmonella Typhimurium intoxication results in decreased Salmonella numbers in the cecum contents of pigs, despite marked effects on Salmonella-host cell interactions. Vet. Res. 2012, 43, 1–18.
29. Lillehoj, H.S.; Lillehoj, E.P. Avian coccidiosis. A review of acquired intestinal immunity and vaccination strategies. Avian Dis. 2000, 44, 408–425.
30. Chapman, H.D.; Barta, J.R.; Blake, D.; Gruber, A.; Jenkins, M.; Smith, N.C.; Suo, X.; Tomley, F.M. A selective review of advances in coccidiosis research. Adv. Parasitol. 2013, 83, 93–171.
31. Lillehoj, H. Role of T lymphocytes and cytokines in coccidiosis. Int. J. Parasitol. 1998, 28, 1071–1081.
32. Lillehoj, H.; Min, W.; Dalloul, R. Recent progress on the cytokine regulation of intestinal immune responses to Eimeria. Poult. Sci. 2004, 83, 611–623.
33. Lillehoj, H.; Kim, C.; Keeler, C.; Zhang, S. Immunogenomic approaches to study host immunity to enteric pathogens. Poult. Sci. 2007, 86, 1491–1500.
34. Girgis, G.N.; Sharif, S.; Barta, J.R.; Boermans, H.J.; Smith, T.K. Immunomodulatory effects of feed-borne fusarium mycotoxins in chickens infected with coccidia. Exp. Biol. Med. 2008, 233, 1411–1420.
35. Girgis, G.N.; Barta, J.R.; Girish, C.K.; Karrow, N.A.; Boermans, H.J.; Smith, T.K. Effects of feed-borne fusarium mycotoxins and an organic mycotoxin adsorbent on immune cell dynamics in the jejunum of chickens infected with Eimeria maxima. Vet. Immunol. Immun. 2010, 138, 218–223.
36. Girgis, G.; Barta, J.; Brash, M.; Smith, T. Morphologic changes in the intestine of broiler breeder pullets fed diets naturally contaminated with fusarium mycotoxins with or without coccidial challenge. Avian Dis. 2010, 54, 67–73.
37. Békési, L.; Hornok, S.; Szigeti, G.; Dobos-Kovács, M.; Széll, Z.; Varga, I. Effect of F-2 and T-2 fusariotoxins on experimental Cryptosporidium baileyi infection in chickens. Int. J. Parasitol. 1997, 27, 1531–1536.
38. Varga, I.; Ványi, A. Interaction of T-2 fusariotoxin with anticoccidial efficacy of lasalocid in chickens. Int. J. Parasitol. 1992, 22, 523–525.
39. Andrews-Polymenis, H.L.; Bäumler, A.J.; McCormick, B.A.; Fang, F.C. Taming the elephant: Salmonella biology, pathogenesis, and prevention. Infect. Immun. 2010, 78, 2356–2369.
40. Ohl, M.E.; Miller, S.I. Salmonella: A model for bacterial pathogenesis. Annu. Rev. Med. 2001, 52, 259–274.
41. Vandenbroucke, V.; Croubels, S.; Verbrugghe, E.; Boyen, F.; De Backer, P.; Ducatelle, R.; Rychlik, I.; Haesebrouck, F.; Pasmans, F. The mycotoxin deoxynivalenol promotes uptake of Salmonella Typhimurium in porcine macrophages, associated with ERK1/2 induced cytoskeleton reorganization. Vet. Res. 2009, 40, doi:10.1051/vetres/2009045.
42. Burel, C.; Tanguy, M.; Guerre, P.; Boilletot, E.; Cariolet, R.; Queguiner, M.; Postollec, G.; Pinton, P.; Salvat, G.; Oswald, I.P. Effect of low dose of fumonisins on pig health: Immune status, intestinal microbiota and sensitivity to Salmonella. Toxins 2013, 5, 841–864.
43. Skjolaas, K.; Burkey, T.; Dritz, S.; Minton, J. Effects of Salmonella enterica serovars Typhimurium (st) and Choleraesuis (sc) on chemokine and cytokine expression in swine ileum and jejunal epithelial cells. Vet. Immunol. Immun. 2006, 111, 199–209.
44. Boyen, F.; Haesebrouck, F.; Maes, D.; van Immerseel, F.; Ducatelle, R.; Pasmans, F. Non-typhoidal Salmonella infections in pigs: A closer look at epidemiology, pathogenesis and control. Vet. Microbiol. 2008, 130, 1–19.
45. Ziprin, R.; Elissalde, M. Effect of T-2 toxin on resistance to systemic Salmonella Typhimurium infection of newly hatched chickens. Am. J. Vet. Res. 1990, 51, 1869–1872.
46. Tai, J.; Pestka, J. Impaired murine resistance to Salmonella Typhimurium following oral exposure to the trichothecene t-2 toxin. Food Cem. Toxicol. 1988, 26, 691–698.
47. Tai, J.-H.; Pestka, J. T-2 toxin impairment of murine response to Salmonella Typhimurium: A histopathologic assessment. Mycopathologia 1990, 109, 149–155.
48. Vidal, D.; Mavet, S. In vitro and in vivo toxicity of t-2 toxin, a fusarium mycotoxin, to mouse peritoneal macrophages. Infect. Immun. 1989, 57, 2260–2264.
49. Mittrücker, H.-W.; Kaufmann, S. Immune response to infection with Salmonella Typhimurium in mice. J. Leukocyte Biol. 2000, 67, 457–463.
50. Tai, J.; Pestka, J. Synergistic interaction between the trichothecene T-2 toxin and Salmonella Typhimurium lipopolysaccharide in C3H/HeN and C3H/HeJ mice. Toxicol. Lett. 1988, 44, 191–200.
51. Hara-Kudo, Y. Effects of deoxynivalenol on Salmonella Enteritidis infection. Mycotoxins 1996, 1996,42, 51–56.
52. Rothkötter, H.; Sowa, E.; Pabst, R. The pig as a model of developmental immunology. Hum. Exp. Toxicol. 2002, 21, 533–536.
53. Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57.
54. Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140.
55. Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201.
56. Oswald, I.P.; Desautels, C.; Laffitte, J.; Fournout, S.; Peres, S.Y.; Odin, M.; le Bars, P.; le Bars, J.; Fairbrother, J.M. Mycotoxin fumonisin B1 increases intestinal colonization by pathogenic escherichia coli in pigs. Appl. Environ. Microb. 2003, 69, 5870–5874.
57. Baines, D.; Sumarah, M.; Kuldau, G.; Juba, J.; Mazza, A.; Masson, L. Aflatoxin, fumonisin and shiga toxin-producing Escherichia coli infections in calves and the effectiveness of celmanax/dairyman’s choice applications to eliminate morbidity and mortality losses. Toxins 2013, 5, 1872–1895.
58. Devriendt, B.; Verdonck, F.; Wache, Y.; Bimczok, D.; Oswald, I.P.; Goddeeris, B.M.; Cox, E. The food contaminant fumonisin B1 reduces the maturation of porcine CD11r1+ intestinal antigen presenting cells and antigen-specific immune responses, leading to a prolonged intestinal etec infection. Vet. Res. 2009, 40, 1–14.
59. Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430.
60. Li, Y.; Ledoux, D.; Bermudez, A.; Fritsche, K.; Rottinghaust, G. Effects of moniliformin on performance and immune function of broiler chicks. Poult. Sci. 2000, 79, 26–32.
61. Li, Y.; Ledoux, D.; Bermudez, A.; Fritsche, K.; Rottinghaus, G. The individual and combined effects of fumonisin B1 and moniliformin on performance and selected immune parameters in turkey poults. Poult. Sci. 2000, 79, 871–878.
62. Barbara, A.J.; Trinh, H.T.; Glock, R.D.; Glenn Songer, J. Necrotic enteritis-producing strains of Clostridium perfringens displace non-necrotic enteritis strains from the gut of chicks. Vet. Microbiol. 2008, 126, 377–382.
63. Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; van Immerseel, F. Necrotic enteritis in broilers: An updated review on the pathogenesis. Avian Pathol. 2011, 40, 341–347.
64. Williams, R. Intercurrent coccidiosis and necrotic enteritis of chickens: Rational, integrated disease management by maintenance of gut integrity. Avian Pathol. 2005, 34, 159–180.
65. Keyburn, A.L.; Boyce, J.D.; Vaz, P.; Bannam, T.L.; Ford, M.E.; Parker, D.; di Rubbo, A.; Rood, J.I.; Moore, R.J. NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens. PLoS Pathog. 2008, 4, doi:10.1371/journal.ppat.0040026.
66. Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Haesebrouck, F.; Timbermont, L.; Verlinden, M.; Janssens, G.P.J.; Eeckhout, M.; de Saeger, S.; et al. Deoxynivalenol predisposes for necrotic enteritis by affecting the intestinal barrier in broilers. In Proceedings of the International Poultry Scientific Forum, Atlanta, Georgia, USA, 28–29 January 2013; pp. 9–10.
67. Crumlish, M.; Dung, T.; Turnbull, J.; Ngoc, N.; Ferguson, H. Identification of Edwardsiella ictaluri from diseased freshwater catfish, Pangasius hypophthalmus (sauvage), cultured in the mekong delta, Vietnam. J. Fish Dis. 2002, 25, 733–736.
68. Ferguson, H.; Turnbull, J.; Shinn, A.; Thompson, K.; Dung, T.T.; Crumlish, M. Bacillary necrosis in farmed Pangasius hypophthalmus (sauvage) from the Mekong delta, Vietnam. J. Fish Dis. 2001, 24, 509–513.
69. Newton, J.; Wolfe, L.; Grizzle, J.; Plumb, J. Pathology of experimental enteric septicaemia in channel catfish, Ictalurus punctatus (rafinesque), following immersion-exposure to Edwardsiella ictaluri. J. Fish Dis. 1989, 12, 335–347.
70. Manning, B.B.; Terhune, J.S.; Li, M.H.; Robinson, E.H.; Wise, D.J.; Rottinghaus, G.E. Exposure to feedborne mycotoxins T-2 toxin or ochratoxin a causes increased mortality of channel catfish challenged with Edwardsiella ictaluri. J. Aquat. Anim. Health 2005, 17, 147–152.
71. Manning, B.B.; Abbas, H.K.; Wise, D.J.; Greenway, T. The effect of feeding diets containing deoxynivalenol contaminated corn on channel catfish (Ictalurus punctatus) challenged with edwardsiella ictaluri. Aquac. Res. 2013, doi:10.1111/are.12123.
72. Hooft, J.M.; Elmor, A.E.H.I.; Encarnação, P.; Bureau, D.P. Rainbow trout (Oncorhynchus mykiss) is extremely sensitive to the feed-borne Fusarium mycotoxin deoxynivalenol. Aquaculture 2011, 311, 224–232.
73. Pósa, R.; Donkó, T.; Bogner, P.; Kovács, M.; Repa, I.; Magyar, T. Interaction of Bordetella bronchiseptica, Pasteurella multocida, and fumonisin B1 in the porcine respiratory tract as studied by computed tomography. Can. J. Vet. Res. 2011, 75, 176–182.
74. Halloy, D.J.; Gustin, P.G.; Bouhet, S.; Oswald, I.P. Oral exposure to culture material extract containing fumonisins predisposes swine to the development of pneumonitis caused by Pasteurella multocida. Toxicology 2005, 213, 34–44.
75. Maes, D.; Segales, J.; Meyns, T.; Sibila, M.; Pieters, M.; Haesebrouck, F. Control of Mycoplasma hyopneumoniae infections in pigs. Vet. Microbiol. 2008, 126, 297–309.
76. Pósa, R.; Magyar, T.; Stoev, S.; Glávits, R.; Donkó, T.; Repa, I.; Kovács, M. Use of computed tomography and histopathologic review for lung lesions produced by the interaction between Mycoplasma hyopneumoniae and fumonisin mycotoxins in pigs. Vet. Pathol. 2013, 50, doi:10.1177/0300985813480510.
77. Chanter, N.; Magyar, T.; Rutter, J.M. Interactions between Bordetella bronchiseptica and toxigenic Pasteurella multocida in atrophic rhinitis of pigs. Res. Vet. Sci. 1989, 47, 48–53.
78. Davies, R.L.; MacCorquodale, R.; Baillie, S.; Caffrey, B. Characterization and comparison of Pasteurella multocida strains associated with porcine pneumonia and atrophic rhinitis. J. Med. Microbiol. 2003, 52, 59–67.
79. Niyo, K.; Richard, J.; Niyo, Y.; Tiffany, L. Effects of T-2 mycotoxin ingestion on phagocytosis of Aspergillus fumigatus conidia by rabbit alveolar macrophages and on hematologic, serum biochemical, and pathologic changes in rabbits. Am. J. Vet. Res. 1988, 49, 1766–1773.
80. Li, S.-J.; Pasmans, F.; Croubels, S.; Verbrugghe, E.; Van Waeyenberghe, L.; Yang, Z.; Haesebrouck, F.; Martel, A. T-2 toxin impairs antifungal activities of chicken macrophages against Aspergillus fumigatus conidia but promotes the pro-inflammatory responses. Avian Pathol. 2013, 42, 457–463.
81. Nibert, M.; Furlong, D.; Fields, B. Mechanisms of viral pathogenesis. Distinct forms of reoviruses and their roles during replication in cells and host. J. Clin. Invest. 1991, 88, 727–934.
82. Li, M.; Cuff, C.F.; Pestka, J. Modulation of murine host response to enteric reovirus infection by the trichothecene deoxynivalenol. Toxicol. Sci. 2005, 87, 134–145.
83. Jones, R. Avian reovirus infections. Rev. Sci. Tech. OIE 2000, 19, 614–625.
84. Jones, R.; Kibenge, F. Reovirus-induced tenosynovitis in chickens: The effect of breed. Avian Pathol. 1984, 13, 511–528.
85. Benavente, J.; Martínez-Costas, J. Avian reovirus: Structure and biology. Virus Res. 2007, 123, 105–119.
86. Li, M.; Cuff, C.F.; Pestka, J.J. T-2 toxin impairment of enteric reovirus clearance in the mouse associated with suppressed immunoglobulin and IFN-γ responses. Toxicol. Appl. Pharm. 2006, 214, 318–325.
87. Chand, R.J.; Trible, B.R.; Rowland, R.R. Pathogenesis of porcine reproductive and respiratory syndrome virus. Curr. Opin. Virol. 2012, 2, 256–263.
88. Rowland, R.; Morrison, R. Challenges and opportunities for the control and elimination of porcine reproductive and respiratory syndrome virus. Transbound. Emerg. Dis. 2012, 59, 55–59.
89. Ramos, C.M.; Martinez, E.M.; Carrasco, A.C.; Puente, J.H.L.; Quezada, F.; Perez, J.T.; Oswald, I.P.; Elvira, S.M. Experimental trial of the effect of fumonisin B and the PRRS virus in swine. J. Anim. Vet. Adv. 2010, 9, 1301–1310.
90. Bane, D.P.; Neumann, E.J.; Hall, W.F.; Harlin, K.S.; Slife, R.L.N. Relationship between fumonisin contamination of feed and mystery swine disease-a case-control study. Mycopathologia 1992, 117, 121–124.
91. Wu, F. Measuring the economic impacts of Fusarium toxins in animal feeds. Anim. Feed Sci. Tech. 2007, 137, 363–374.
92. Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144.
93. Maresca, M.; Fantini, J. Some food-associated mycotoxins as potential risk factors in humans predisposed to chronic intestinal inflammatory diseases. Toxicon 2010, 56, 282–294.
94. Wagacha, J.; Muthomi, J. Mycotoxin problem in africa: Current status, implications to food safety and health and possible management strategies. Int. J. Food Microbiol. 2008, 124, 1–12.
95. Wild, C.P.; Gong, Y.Y. Mycotoxins and human disease: A largely ignored global health issue. Carcinogenesis 2010, 31, 71–82.
96. Wan, L.Y.M.; Turner, P.C.; El-Nezami, H. Individual and combined cytotoxic effects of Fusarium toxins (deoxynivalenol, nivalenol, zearalenone and fumonisins B1) on swine jejunal epithelial cells. Food Chem. Toxicol. 2013, 57, 276–283.
97. Grenier, B.; Oswald, I. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313.
98. Grenier, B.; Loureiro-Bracarense, A.P.; Lucioli, J.; Pacheco, G.D.; Cossalter, A.M.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Individual and combined effects of subclinical doses of deoxynivalenol and fumonisins in piglets. Mol. Nutr. Food. Res. 2011, 55, 761–771.
99. De Boevre, M.; di Mavungu, J.D.; Landschoot, S.; Audenaert, K.; Eeckhout, M.; Maene, P.; Haesaert, G.; De Saeger, S. Natural occurrence of mycotoxins and their masked forms in food and feed products. World Mycotoxin J. 2012, 5, 207–219.
100. Nagl, V.; Schwartz, H.; Krska, R.; Moll, W.-D.; Knasmüller, S.; Ritzmann, M.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in rats. Toxicol. Lett. 2012, 213, 367–373.
101. Dall’Erta, A.; Cirlini, M.; Dall’Asta, M.; del Rio, D.; Galaverna, G.; Dall’Asta, C. Masked mycotoxins are efficiently hydrolysed by the human colonic microbiota, releasing their toxic aglycones. Chem. Res. Toxicol. 2013, 26, 305–312.
102. Broekaert, N.; Devreese, M.; de Mil, T.; Fraeyman, S.; de Baere, S.; de Saeger, S.; de Backer, P.; Croubels, S. Development and validation of an LC-MS/MS method for the toxicokinetic study of deoxynivalenol and its acetylated derivatives in animal plasma. Anal. Bioanal. Chem. Submitted.
103. Paterson, R.R.M.; Lima, N. How will climate change affect mycotoxins in food? Food Res. Int. 2010, 43, 1902–1914.
104. Magan, N.; Medina, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre-and postharvest. Plant Pathol. 2011, 60, 150–163.
105. Shuman, E.K. Global climate change and infectious diseases. N. Engl. J. Med. 2010, 362, 1061–1063.
106. Gerberick, G.F.; Sorenson, W.; Lewis, D. The effects of T-2 toxin on alveolar macrophage function in vitro. Environ. Res. 1984, 33, 246–260.
107. Deshmukh, S.; Asrani, R.; Jindal, N.; Ledoux, D.; Rottinghaus, G.; Sharma, M.; Singh, S. Effects of fusarium moniliforme culture material containing known levels of fumonisin B1 on progress of Salmonella Gallinarum infection in Japanese quail: Clinical signs and hematologic studies. Avian Dis. 2005, 49, 274–280.
108. Kanai, K.; Kondo, E. Decreased resistance to mycobacterial infection in mice fed a trichothecene compound (T-2 toxin). Jpn. J. Med. Sci. Biol. 1984, 37, 97.
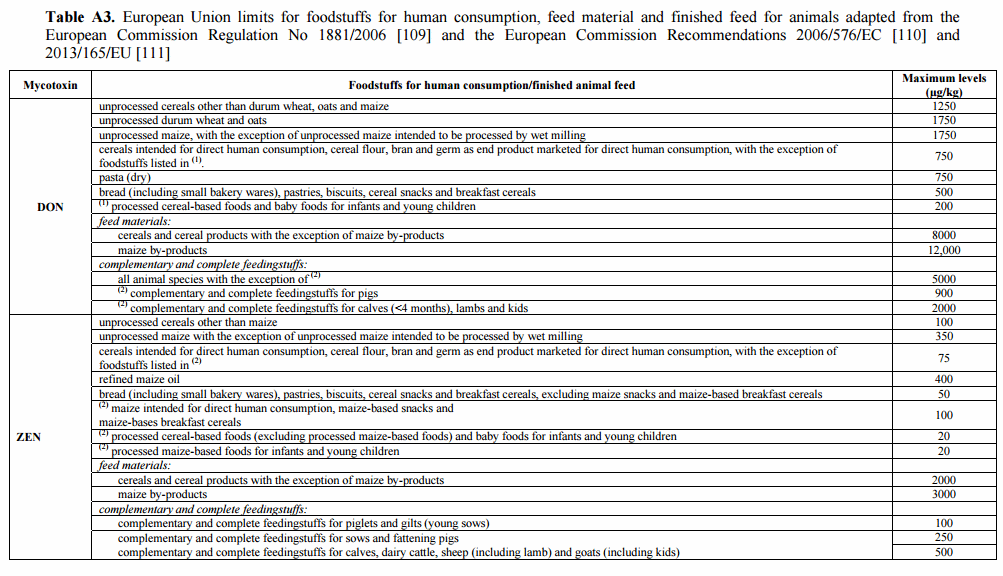
109. European Commission. Commission Regulation (EC) of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs no. 1881/2006. Off. J. Eur. Union 2006, L364, 5–24.
110. European Commission. Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding (2006/576/EC). Off. J. Eur. Union 2006, L229, 7–9.
111. European Commission. Commission Recommendation of 27 March 2013 on the presence of T-2 and HT-2 toxin in cereals and cereal products (2013/165/EU). Off. J. Eur. Union 2013, L91, 12–15.
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).