Arginine impact in immune regulation and microbial homeostasis in poultry
Beyond protein synthesis: the emerging role of arginine in poultry nutrition and host-microbe interactions
Arginine is a functional amino acid essential for various physiological processes in poultry. The dietary essentiality of arginine in poultry stems from the absence of the enzyme carbamoyl phosphate synthase-I. The specific requirement for arginine in poultry varies based on several factors, such as age, dietary factors, and physiological status. Additionally, arginine absorption and utilization are also influenced by the presence of antagonists. However, dietary interventions can mitigate the effect of these factors affecting arginine utilization. In poultry, arginine is utilized by four enzymes, namely, inducible nitric oxide synthase arginase, arginine decarboxylase and arginine: glycine amidinotransferase (AGAT). The intermediates and products of arginine metabolism by these enzymes mediate the different physiological functions of arginine in poultry. The most studied function of arginine in humans, as well as poultry, is its role in immune response. Arginine exerts immunomodulatory functions primarily through the metabolites nitric oxide (NO), ornithine, citrulline, and polyamines, which take part in inflammation or the resolution of inflammation. These properties of arginine and arginine metabolites potentiate its use as a nutraceutical to prevent the incidence of enteric diseases in poultry. Furthermore, arginine is utilized by the poultry gut microbiota, the metabolites of which might have important implications for gut microbial composition, immune regulation, metabolism, and overall host health. This comprehensive review provides insights into the multifaceted roles of arginine and arginine metabolites in poultry nutrition and wellbeing, with particular emphasis on the potential of arginine in immune regulation and microbial homeostasis in poultry.
KEYWORDS arginine, poultry, gut health, nutraceuticals, immune response
1 Introduction
2 Arginine in poultry production
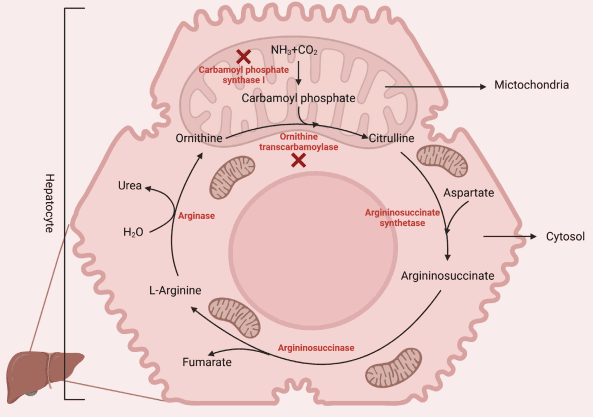
3 Arginine absorption and transport mechanisms
4 Nutritional antagonism: interaction of other amino acids with arginine
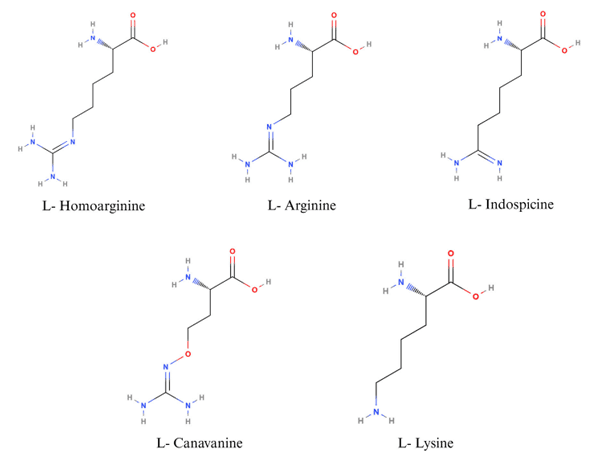
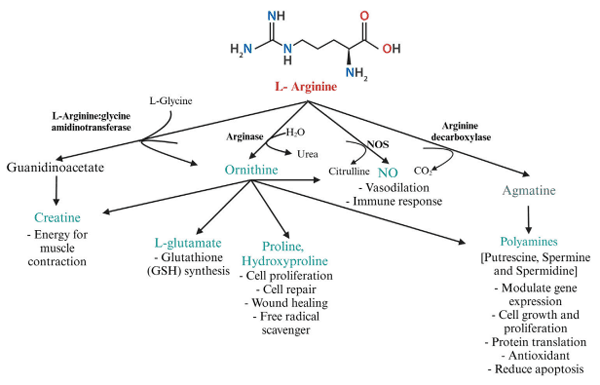
5 Arginine metabolism and physiological effects of metabolites in poultry
6 Arginine sparing effects of arginine metabolites
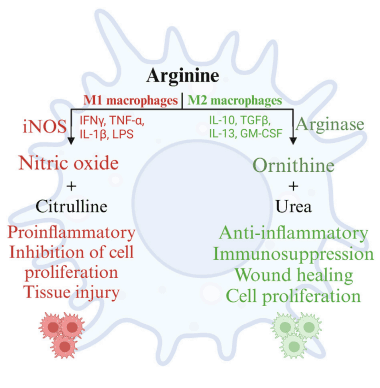
7 Arginine and the macrophage dichotomy
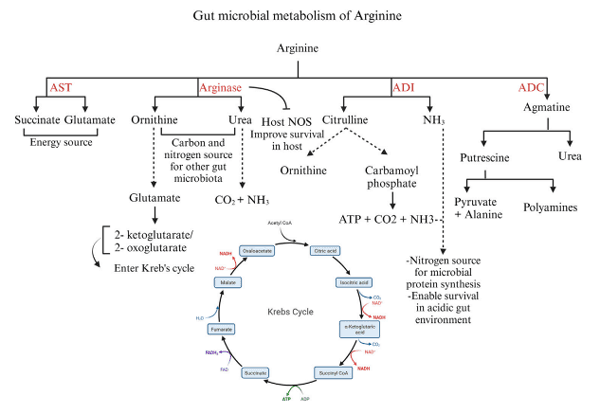
8 Arginine and the gut microbiota
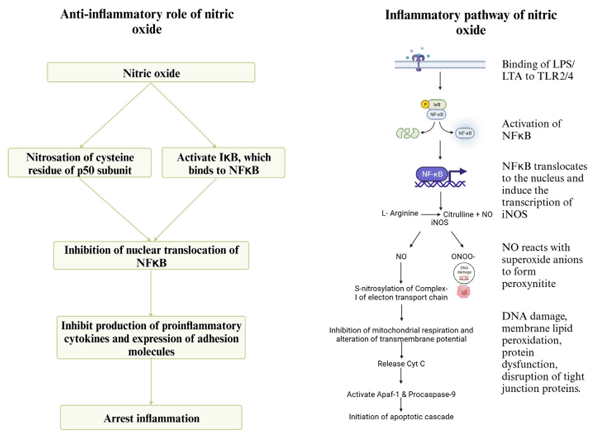
9 Arginine and necrotic enteritis
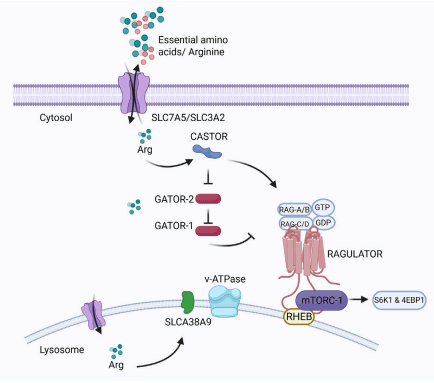
10 Emerging trends and future prospects of arginine in poultry production
Author contributions
Funding
Conflict of interest
Publisher’s note
2022-Cobb500-Broiler-Performance-Nutrition-Supplement, 2022 2022-Cobb500- Broiler-Performance-Nutrition-Supplement (2022) 2022-Cobb500-BroilerPerformance-Nutrition-Supplement.
Abramson, S. B., Amin, A. R., Clancy, R. M., and Attur, M. (2001). The role of nitric oxide in tissue destruction. Best Pract. Res. Clin. Rheumatology 15 (5), 831–845. doi:10. 1053/berh.2001.0196
Adams, S., Che, D., Qin, G., Farouk, M. H., Hailong, J., and Rui, H. (2019). Novel biosynthesis, metabolism and physiological functions of L-homoarginine. Curr. Protein Peptide Sci. 20 (2), 184–193. doi:10.2174/1389203719666181026170049
Ahmadipour, B., Khajali, F., and Sharifi, M. R. (2018). Effect of guanidinoacetic acid supplementation on growth performance and gut morpholog yin broiler chickens. Poult. Sci. J. 6 (1), 19–24.
Akasaka, N., and Fujiwara, S. (2020). The therapeutic and nutraceutical potential of agmatine, and its enhanced production using Aspergillus oryzae. Amino Acids 52 (2), 181–197. doi:10.1007/s00726-019-02720-7
Al-Daraji, H. J., and Salih, A. M. (2012). Effect of dietary L-arginine on productive performance of broiler chickens. Pak. J. Nutr. 11 (3), 252–257. doi:10.3923/pjn.2012.252.257
Allen, P. C., and Fetterer, R. H. (2000). Effect of Eimeria acervulina infections on plasma L-arginine. Poult. Sci. 79 (10), 1414–1417. doi:10.1093/ps/79.10.1414
Almquist, H. J., Mecchi, E., and Kratzer, F. H. (1941). Creatine formation in the chick. J. Biol. Chem. 141 (2), 365–373. doi:10.1016/s0021-9258(18)72782-8
Amiri, M., Ghasemi, H. A., Hajkhodadadi, I., and Farahani, A. H. K. (2019). Efficacy of guanidinoacetic acid at different dietary crude protein levels on growth performance, stress indicators, antioxidant status, and intestinal morphology in broiler chickens subjected to cyclic heat stressficacy of guanidinoacetic acid at different dietary crude protein levels on growth performance, stress indicators, antioxidant status, and intestinal morphology in broiler chickens subjected to cyclic heat stress. Anim. Feed Sci. Technol. 254, 114208. doi:10.1016/j.anifeedsci.2019.114208
Anderson, J. O., and Combs, G. F. (1952). Effect of single amino acid excesses on glucose metabolism and chick growth, as influenced by the dietary amino acid balance. J. Nutr. 46 (2), 161–170. doi:10.1093/jn/46.2.161
Applegate, T. J., and Angel, R. (2014). Nutrient requirements of poultry publication: history and need for an update. J. Appl. Poult. Res. 23 (3), 567–575. doi:10.3382/japr. 2014-00980
Application of nutritional immunology (2022). “Application of nutritional immunology in the mitigation of economic and production losses in the poultry industry associated with food-borne pathogens, coccidiosis, and necrotic enteritis,” in Proceedings of the Arkansas Nutrition Conference.
Arginine sparing potential of guanidinoacetic acid in broiler nutrition (2018). “Arginine sparing potential of guanidinoacetic acid in broiler nutrition,” in PSA Latin American Scientific Conference.
Austic, R. E., and Nesheim, M. C. (1970). Role of kidney arginase in variations of the arginine requirement of chicks. J. Nutr. 100 (7), 855–867. doi:10.1093/jn/100. 7.855
Ball, R. O., Urschel, K. L., and Pencharz, P. B. (2007). Nutritional consequences of interspecies differences in arginine and lysine metabolism. J. Nutr. 137 (6), 1626S–1641S. doi:10.1093/jn/137.6.1626S
Bell, E. A. (2003). Nonprotein amino acids of plants: significance in medicine, nutrition, and agriculture. J. Agric. Food Chem. 51 (10), 2854–2865. doi:10.1021/ jf020880w
Benoit, M., Desnues, B., and Mege, J. (2008). Macrophage polarization in bacterial infections. J. Immunol. 181 (6), 3733–3739. doi:10.4049/jimmunol.181.6.3733
Bowen, O. T., Erf, G. F., Chapman, M. E., and Wideman, R. F., Jr (2007). Plasma nitric oxide concentrations in broilers after intravenous injections of lipopolysaccharide or microparticles. Poult. Sci. 86 (12), 2550–2554. doi:10.3382/ps.2007-00288
Brake, J., Balnave, D., and Dibner, J. J. (1998). Optimum dietary arginine: lysine ratio for broiler chickens is altered during heat stress in association with changes in intestinal uptake and dietary sodium chloride. Br. Poult. Sci. 39 (5), 639–647. doi:10.1080/ 00071669888511
Bröer, S., and Fairweather, S. J. (2018). Amino acid transport across the mammalian intestine. Compr. Physiol. 9 (1), 343–373. doi:10.1002/cphy.c170041
Bron, P. A., Grangette, C., Mercenier, A., De Vos, W. M., and Kleerebezem, M. (2004). Identification of Lactobacillus plantarum genes that are induced in the gastrointestinal tract of mice. J. Bacteriol. 186 (17), 5721–5729. doi:10.1128/JB. 186.17.5721-5729.2004
Chamruspollert, M., Pesti, G. M., and Bakalli, R. I. (2002). Dietary interrelationships among arginine, methionine, and lysine in young broiler chicks. Br. J. Nutr. 88 (6), 655–660. doi:10.1079/BJN2002732
Choi, B. S., Martinez-Falero, I. C., Corset, C., Munder, M., Modolell, M., Müller, I., et al. (2009). Differential impact of L-arginine deprivation on the activation and effector functions of T cells and macrophages. J. Leucocyte Biol. 85 (2), 268–277. doi:10.1189/jlb. 0508310
Choi, Y., Choi, J., Groisman, E. A., Kang, D., Shin, D., and Ryu, S. (2012). Expression of STM4467-encoded arginine deiminase controlled by the STM4463 regulator contributes to Salmonella enterica serovar Typhimurium virulence. Infect. Immun. 80 (12), 4291–4297. doi:10.1128/IAI.00880-12
Chowdhury, V. S., Han, G., Bahry, M. A., Tran, P. V., Do, P. H., Yang, H., et al. (2017). L-Citrulline acts as potential hypothermic agent to afford thermotolerance in chicks. J. Therm. Biol. 69, 163–170. doi:10.1016/j.jtherbio.2017.07.007
Closs, E. I., and Mann, G. E. (2000). Membrane transport of L-arginine and cationic amino acid analogs. Nitric Oxide, 225–241. doi:10.1016/b978-012370420-7/50015-0
Closs, E. I., Simon, A., Vékony, N., and Rotmann, A. (2004). Plasma membrane transporters for arginine. J. Nutr. 134 (10), 2752S–2767S. doi:10.1093/jn/134.10.2752S
Corzo, A., Lee, J., Vargas, J. I., Silva, M., and Pacheco, W. J. (2021). Determination of the optimal digestible arginine to lysine ratio in Ross 708 male broilers. J. Appl. Poult. Res. 30 (1), 100136. doi:10.1016/j.japr.2020.100136
Cummings, N. E., and Lamming, D. W. (2017). Regulation of metabolic health and aging by nutrient-sensitive signaling pathways. Mol. Cell Endocrinol. 455, 13–22. doi:10. 1016/j.mce.2016.11.014
Dai, Z., Wu, G., and Zhu, W. (2011). Amino acid metabolism in intestinal bacteria: links between gut ecology and host health. Front. Bioscience-Landmark 16 (5), 1768–1786. doi:10.2741/3820
Dao, H. T., Clay, J. W., Sharma, N. K., Bradbury, E. J., and Swick, R. A. (2022b). Effects of l-arginine and l-citrulline supplementation in reduced protein diets on cecal fermentation metabolites of broilers under normal, cyclic warm temperature and necrotic enteritis challenge. Livest. Sci. 257, 104826. doi:10.1016/j.livsci.2022. 104826
Dao, H. T., Sharma, N. K., Barekatain, R., Kheravii, S. K., Bradbury, E. J., Wu, S., et al. (2022c). Supplementation of reduced protein diets with. Animal Prod. Sci. 62, 1250–1265. doi:10.1071/an21394
Dao, H. T., Sharma, N. K., Bradbury, E. J., and Swick, R. A. (2021a). Effects of L-arginine and L-citrulline supplementation in reduced protein diets for broilers under normal and cyclic warm temperature. Anim. Nutr. 7 (4), 927–938. doi:10.1016/j.aninu.2020.12.010
Dao, H. T., Sharma, N. K., Bradbury, E. J., and Swick, R. A. (2021b). Response of meat chickens to different sources of arginine in low-protein diets. J. Anim. Physiol. Anim. Nutr. 105 (4), 731–746. doi:10.1111/jpn.13486
Dao, H. T., Sharma, N. K., Daneshmand, A., Kumar, A., Bradbury, E. J., Wu, S., et al. (2022a). Supplementation of reduced protein diets with. Animal Prod. Sci. 62, 1236–1249. doi:10.1071/an21393
Dao, H. T., Sharma, N. K., Kheravii, S. K., Bradbury, E. J., Wu, S., and Swick, R. A. (2022d). Supplementation of reduced protein diets with. Animal Prod. Sci. 62 (13), 1266–1279. doi:10.1071/an21395
DeGroot, A. A., Braun, U., and Dilger, R. N. (2018). Efficacy of guanidinoacetic acid on growth and muscle energy metabolism in broiler chicks receiving arginine-deficient diets. Poult. Sci. 97 (3), 890–900. doi:10.3382/ps/pex378
D’mello, J. (2003a). Adverse effects of amino acids. Amino acids animal Nutr. 2, 125–142. doi:10.1079/9780851996547.0125 D’mello, J. (2003b). Amino acids as multifunctional molecules. Amino acids in animal nutrition. (Wallingford UK: Cabi Publishing). 1–14.
D’mello, J., and Lewis, D. (1970). Amino acid interactions in chick nutrition: I. The interrelationship between lysine and arginine. Br. Poult. Sci. 11 (3), 299–311. doi:10. 1080/00071667008415820
El-Hattab, A. W., Emrick, L. T., Craigen, W. J., and Scaglia, F. (2012). Citrulline and arginine utility in treating nitric oxide deficiency in mitochondrial disorders. Mol. Genet. Metab. 107 (3), 247–252. doi:10.1016/j.ymgme.2012.06.018
Esser, A., Gonçalves, D., Rorig, A., Cristo, A. B., Perini, R., and Fernandes, J. (2017). Effects of guanidionoacetic acid and arginine supplementation to vegetable diets fed to broiler chickens subjected to heat stress before slaughter. Braz. J. Poult. Sci. 19, 429–436. doi:10.1590/1806-9061-2016-0392
Fagundes, N. S., Milfort, M. C., Williams, S. M., Da Costa, M. J., Fuller, A. L., Menten, J. F., et al. (2020). Dietary methionine level alters growth, digestibility, and gene expression of amino acid transporters in meat-type chickens. Poult. Sci. 99 (1), 67–75. doi:10.3382/ps/pez588
Fathima, S., Hakeem, W. G. A., Shanmugasundaram, R., and Selvaraj, R. K. (2022). Necrotic enteritis in broiler chickens: a review on the pathogen, pathogenesis, and prevention. Microorganisms 10 (10), 1958. doi:10.3390/microorganisms10101958
Faure, M., Choné, F., Mettraux, C., Godin, J., Béchereau, F., Vuichoud, J., et al. (2007). Threonine utilization for synthesis of acute phase proteins, intestinal proteins, and mucins is increased during sepsis in rats. J. Nutr. 137 (7), 1802–1807. doi:10.1093/jn/137.7.1802
Fletcher, M. T., Al Jassim, R. A., and Cawdell-Smith, A. J. (2015). The occurrence and toxicity of indospicine to grazing animals. Agriculture 5 (3), 427–440. doi:10.3390/ agriculture5030427
Fosoul, SSAS, Azarfar, A., Gheisari, A., and Khosravinia, H. (2018). Energy utilisation of broiler chickens in response to guanidinoacetic acid supplementation in diets with various energy contents. Br. J. Nutr. 120 (2), 131–140. doi:10.1017/S0007114517003701
Geiger, R., Rieckmann, J. C., Wolf, T., Basso, C., Feng, Y., Fuhrer, T., et al. (2016). L-arginine modulates T cell metabolism and enhances survival and anti-tumor activity. Cell 167 (3), 829–842. doi:10.1016/j.cell.2016.09.031
Ghamari Monavvar, H., Moghaddam, G., and Ebrahimi, M. (2020). A review on the effect of arginine on growth performance, meat quality, intestine morphology, and immune system of broiler chickens. Iran. J. Appl. Animal Sci. 10 (4), 587–594.
Gharavi, A. T., Hanjani, N. A., Movahed, E., and Doroudian, M. (2022). The role of macrophage subtypes and exosomes in immunomodulation. Cell Mol. Biol. Lett. 27 (1), 83. doi:10.1186/s11658-022-00384-y
Gobert, A. P., McGee, D. J., Akhtar, M., Mendz, G. L., Newton, J. C., Cheng, Y., et al. (2001). Helicobacter pylori arginase inhibits nitric oxide production by eukaryotic cells: a strategy for bacterial survival. Proc. Natl. Acad. Sci. 98 (24), 13844–13849. doi:10.1073/ pnas.241443798
Grishin, D. V., Zhdanov, D. D., Pokrovskaya, M. V., and Sokolov, N. N. (2020). D-amino acids in nature, agriculture and biomedicine. All Life 13 (1), 11–22. doi:10. 1080/21553769.2019.1622596
Haghikia, A., Yanchev, G. R., Kayacelebi, A. A., Hanff, E., Bledau, N., Widera, C., et al. (2017). The role of L-arginine/L-homoarginine/nitric oxide pathway for aortic distensibility and intima-media thickness in stroke patients. Amino Acids 49 (6), 1111–1121. doi:10.1007/s00726-017-2409-2
Ham, D. J., Gleeson, B. G., Chee, A., Baum, D. M., Caldow, M. K., Lynch, G. S., et al. (2015). L-Citrulline protects skeletal muscle cells from cachectic stimuli through an iNOS-dependent mechanism. PLoS One 10 (10), e0141572. doi:10.1371/journal.pone. 0141572
Hegarty, M. P., and Pound, A. W. (1970). Indospicine, a hepatotoxic amino acid from Indigofera spicat A: isolation, structure, and biological studies. Aust. J. Biol. Sci. 23 (4), 831–842. doi:10.1071/bi9700831
Hernández, V. M., Arteaga, A., and Dunn, M. F. (2021). Diversity, properties and functions of bacterial arginases. FEMS Microbiol. Rev. 45 (6), fuab034. doi:10.1093/ femsre/fuab034
Ide, A. A., Hernández, V. M., Medina-Aparicio, L., Carcamo-Noriega, E., Girard, L., Hernández-Lucas, I., et al. (2020). Genetic regulation, biochemical properties and physiological importance of arginase from Sinorhizobium meliloti. Microbiology 166 (5), 484–497. doi:10.1099/mic.0.000909
Investigation of protein quality (1968). “Investigation of protein quality--ileal recovery of amino acids,” in Federation Proceedings.
Jahanian, R. (2009). Immunological responses as affected by dietary protein and arginine concentrations in starting broiler chicks. Poult. Sci. 88 (9), 1818–1824. doi:10. 3382/ps.2008-00386
Jimoh, O. A., Akinola, M. O., Oyeyemi, B. F., Oyeyemi, W. A., Ayodele, S. O., Omoniyi, I. S., et al. (2021). Potential of watermelon (Citrullus lanatus) to maintain oxidative stability of rooster semen for artificial insemination. J. Animal Sci. Technol. 63 (1), 46–57. doi:10.5187/jast.2021.e21
Johnson, S. L.-C. (2017). United States: US food and drug administration FDA. 2. Jones, J. D., Petersburg, S. J., and Burnett, P. C. (1967). The mechanism of the lysinearginine antagonism in the chick: effect of lysine on digestion, kidney arginase, and liver transamidinase. J. Nutr. 93 (1), 103–116. doi:10.1093/jn/93.1.103
Jung, J. W., Macalino, S. J. Y., Cui, M., Kim, J. E., Kim, H., Song, D., et al. (2019). Transmembrane 4 L six family member 5 senses arginine for mTORC1 signaling. Cell Metab. 29 (6), 1306–1319. doi:10.1016/j.cmet.2019.03.005
Kadirvel, R., and Kratzer, F. H. (1974). Uptake of L-arginine and L-lysine by the small intestine and its influence on arginine-lysine antagonism in chicks. J. Nutr. 104 (3), 339–343. doi:10.1093/jn/104.3.339
Kaul, S., Sharma, S. S., and Mehta, I. K. (2008). Free radical scavenging potential of L-proline: evidence from in vitro assays. Amino Acids 34 (2), 315–320. doi:10.1007/ s00726-006-0407-x
Khajali, F., Lemme, A., and Rademacher-Heilshorn, M. (2020). Guanidinoacetic acid as a feed supplement for poultry. Worlds Poult. Sci. J. 76 (2), 270–291. doi:10.1080/ 00439339.2020.1716651
Khajali, F., and Wideman, R. F. (2010). Dietary arginine: metabolic, environmental, immunological and physiological interrelationships. Worlds Poult. Sci. J. 66 (4), 751–766. doi:10.1017/s0043933910000711
Kidd, M. T., Kerr, B. J., and Anthony, N. B. (1997). Dietary interactions between lysine and threonine in broilers. Poult. Sci. 76 (4), 608–614. doi:10.1093/ps/76.4.608
Kim, Y. J., Lee, J., Lee, J. J., Jeon, S. M., Silwal, P., Kim, I. S., et al. (2022). Argininemediated gut microbiome remodeling promotes host pulmonary immune defense against nontuberculous mycobacterial infection. Gut microbes 14 (1), 2073132. doi:10.1080/19490976.2022.2073132
Kishton, R. J., Sukumar, M., and Restifo, N. P. (2016). Arginine arms T cells to thrive and survive. Cell metab. 24 (5), 647–648. doi:10.1016/j.cmet.2016.10.019
Kodambashi Emami, N., Golian, A., Rhoads, D. D., and Danesh Mesgaran, M. (2017). Interactive effects of temperature and dietary supplementation of arginine or guanidinoacetic acid on nutritional and physiological responses in male broiler chickens. Br. Poult. Sci. 58 (1), 87–94. doi:10.1080/00071668.2016.1257779
Korhonen, R., Korpela, R., and Moilanen, E. (2002). Signalling mechanisms involved in the induction of inducible nitric oxide synthase by Lactobacillus rhamnosus GG, endotoxin, and lipoteichoic acid. Inflammation 26 (5), 207–214. doi:10.1023/a: 1019720701230
Kusano, T., Berberich, T., Tateda, C., and Takahashi, Y. (2008). Polyamines: essential factors for growth and survival. Planta 228, 367–381. doi:10.1007/s00425-008-0772-7
Lehninger, A. L., Nelson, D. L., and Cox, M. M. (2005). Lehninger principles of biochemistry. Macmillan.
Lemme, A., Ringel, J., Sterk, A., and Young, J. F. (2007). “Supplemental guanidino acetic acid affects energy metabolism of broilers,” in Proceedings 16th European Symposium on Poultry Nutrition (Strasbourg France).
Leong, H. X., Simkevich, C., Lesieur-Brooks, A., Lau, B. W., Fugere, C., Sabo, E., et al. (2006). Short-term arginine deprivation results in large-scale modulation of hepatic gene expression in both normal and tumor cells: microarray bioinformatic analysis. Nutr. metabolism 3, 37–13. doi:10.1186/1743-7075-3-37
Le Plenier, S., Walrand, S., Cynober, L., and Moinard, C. (2011). OP049 direct action of citrulline on muscle protein synthesis: role of the mtorc1 pathway. Clin. Nutr. Suppl. 1 (6), 20. doi:10.1016/s1744-1161(11)70049-7
Li, C., and Lu, C. (2009). Arginine racemization by coupled catabolic and anabolic dehydrogenases. Proc. Natl. Acad. Sci. 106 (3), 906–911. doi:10.1073/pnas.0808269106
Li, Z., Wang, L., Ren, Y., Huang, Y., Liu, W., Lv, Z., et al. (2022). Arginase: shedding light on the mechanisms and opportunities in cardiovascular diseases. Cell Death Discov. 8 (1), 413. doi:10.1038/s41420-022-01200-4
Lima, M. B., Sakomura, N. K., Silva, E. P., Leme, B. B., Malheiros, E. B., Peruzzi, N. J., et al. (2020). Arginine requirements for maintenance and egg production for broiler breeder hens. Anim. Feed Sci. Technol. 264, 114466. doi:10.1016/j.anifeedsci.2020.114466
Linder, C. H. (2016). No title. Biochemical and functional properties of mammalian bone alkaline phosphatase isoforms during osteogenesis.
Liu, G., Ajao, A. M., Shanmugasundaram, R., Taylor, J., Ball, E., Applegate, T. J., et al. (2023). The effects of arginine and branched-chain amino acid supplementation to reduced-protein diet on intestinal health, cecal short-chain fatty acid profiles, and immune response in broiler chickens challenged with Eimeria spp. Poult. Sci. 102 (7), 102773. doi:10.1016/j.psj.2023.102773
Lu, C. (2006). Pathways and regulation of bacterial arginine metabolism and perspectives for obtaining arginine overproducing strains. Appl. Microbiol. Biotechnol. 70, 261–272. doi:10.1007/s00253-005-0308-z
Lumeng, C. N., Bodzin, J. L., and Saltiel, A. R. (2007). Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Invest 117 (1), 175–184. doi:10.1172/JCI29881
MacMicking, J., Xie, Q., and Nathan, C. (1997). Nitric oxide and macrophage function. Annu. Rev. Immunol. 15, 323–350. doi:10.1146/annurev.immunol.15.1.323
Majdeddin, M., Braun, U., Lemme, A., Golian, A., Kermanshahi, H., De Smet, S., et al. (2020). Guanidinoacetic acid supplementation improves feed conversion in broilers subjected to heat stress associated with muscle creatine loading and arginine sparing. Poult. Sci. 99 (9), 4442–4453. doi:10.1016/j.psj.2020.05.023
Marco, M. L., Bongers, R. S., de Vos, W. M., and Kleerebezem, M. (2007). Spatial and temporal expression of Lactobacillus plantarum genes in the gastrointestinal tracts of mice. Appl. Environ. Microbiol. 73 (1), 124–132. doi:10.1128/AEM.01475-06
Martí i Líndez, A., and Reith, W. (2021). Arginine-dependent immune responses. Cell. Mol. Life Sci. 78 (13), 5303–5324. doi:10.1007/s00018-021-03828-4
Maynard, C. W., and Kidd, M. T. (2022). Broiler amino acid research: then and now. Broiler Industry: IntechOpen.
McCafferty, D., Mudgett, J. S., Swain, M. G., and Kubes, P. (1997). Inducible nitric oxide synthase plays a critical role in resolving intestinal inflammation. Gastroenterology 112 (3), 1022–1027. doi:10.1053/gast.1997.v112.pm9041266
McNab, J. M. (1994). Amino acid digestibility and availability studies with poultry.
Meyer, L. E., Machado, L. B., Santiago, A. P. S., da-Silva, W. S., De Felice, F. G., Holub, O., et al. (2006). Mitochondrial creatine kinase activity prevents reactive oxygen species generation: antioxidant role of mitochondrial kinase-dependent ADP re-cycling activity. J. Biol. Chem. 281 (49), 37361–37371. doi:10.1074/jbc.M604123200
Miri, B., Ghasemi, H. A., Hajkhodadadi, I., and Farahani, A. H. K. (2022). Effects of low eggshell temperatures during incubation, in ovo feeding of L-arginine, and post-hatch dietary guanidinoacetic acid on hatching traits, performance, and physiological responses of broilers reared at low ambient temperature. Poult. Sci. 101 (1), 101548. doi:10.1016/j.psj.2021.101548
Miska, K. B., and Fetterer, R. H. (2017). The mRNA expression of amino acid and sugar transporters, aminopeptidase, as well as the di-and tri-peptide transporter PepT1 in the intestines of Eimeria infected broiler chickens. Poult. Sci. 96 (2), 465–473. doi:10.3382/ps/pew303
Miyashita, Y., Kuraji, R., Ito, H., and Numabe, Y. (2022). Wound healing in periodontal disease induces macrophage polarization characterized by different arginine-metabolizing enzymes. J. Periodont Res. 57 (2), 357–370. doi:10.1111/jre.12965
Molderings, G. J., and Haenisch, B. (2012). Agmatine (decarboxylated L-arginine): physiological role and therapeutic potential. Pharmacol. Ther. 133 (3), 351–365. doi:10. 1016/j.pharmthera.2011.12.005
Molecular and epigenetic basis of macrophage polarized activation. Seminars in immunology: Elsevier; 2015.
Montanez, R., Rodriguez-Caso, C., Sanchez-Jimenez, F., and Medina, M. A. (2008). In silico analysis of arginine catabolism as a source of nitric oxide or polyamines in endothelial cells. Amino Acids 34 (2), 223–229. doi:10.1007/s00726-007-0502-7
Muñoz-Fernández, M. A., and Fresno, M. (1998). The role of tumour necrosis factor, interleukin 6, interferon-γ and inducible nitric oxide synthase in the development and pathology of the nervous system. Prog. Neurobiol. 56 (3), 307–340. doi:10.1016/s0301- 0082(98)00045-8
Myers, G. S., Rasko, D. A., Cheung, J. K., Ravel, J., Seshadri, R., DeBoy, R. T., et al. (2006). Skewed genomic variability in strains of the toxigenic bacterial pathogen, Clostridium perfringens. Genome Res. 16 (8), 1031–1040. doi:10.1101/gr.5238106
Nabi, F., Arain, M. A., Bhutto, Z. A., Shah, Q. A., Bangulzai, N., Ujjan, N. A., et al. (2022). Effect of early feeding of L-arginine and L-threonine on hatchability and post-hatch performance of broiler chicken. Trop. Anim. Health Prod. 54 (6), 380. doi:10.1007/s11250-022-03378-2
Nathan, C., and Xie, Q. (1994). Nitric oxide synthases: roles, tolls, and controls. Cell 78 (6), 915–918. doi:10.1016/0092-8674(94)90266-6
National Research Council (1994a). Nutrient requirement of poultry. Washington, DC, USA: 9th revised editionNational Academic Press. National Research Council (1994b). Nutrient requirements of poultry. National Academies Press.
Nogueira, B. R. F., Sakomura, N. K., Reis, M. P., Leme, B. B., Létourneau-Montminy, M., and Viana, G. S. (2021). Modelling broiler requirements for lysine and arginine. Animals 11 (10), 2914. doi:10.3390/ani11102914
Osowska, S., Duchemann, T., Walrand, S., Paillard, A., Boirie, Y., Cynober, L., et al. (2006). Citrulline modulates muscle protein metabolism in old malnourished rats. Am. J. PhysiologyEndocrinology Metabolism 291 (3), E582–E586. doi:10.1152/ajpendo.00398.2005
Phang, J. M., Donald, S. P., Pandhare, J., and Liu, Y. (2008). The metabolism of proline, a stress substrate, modulates carcinogenic pathways. Amino Acids 35 (4), 681–690. doi:10.1007/s00726-008-0063-4
Portocarero, N., and Braun, U. (2021). The physiological role of guanidinoacetic acid and its relationship with arginine in broiler chickens. Poult. Sci. 100 (7), 101203. doi:10. 1016/j.psj.2021.101203
Potoka, D. A., Nadler, E. P., Upperman, J. S., and Ford, H. R. (2002). Role of nitric oxide and peroxynitrite in gut barrier failure. World J. Surg. 26 (7), 806–811. doi:10. 1007/s00268-002-4056-2
Qiu, Y., Yang, X., Wang, L., Gao, K., and Jiang, Z. (2019). L-arginine inhibited inflammatory response and oxidative stress induced by lipopolysaccharide via arginase1 signaling in IPEC-J2 cells. Int. J. Mol. Sci. 20 (7), 1800. doi:10.3390/ijms20071800
Qureshi, M. A. (2003). Avian macrophage and immune response: an overview. Poult. Sci. 82 (5), 691–698. doi:10.1093/ps/82.5.691
Ravindran, V., Hew, L. I., Ravindran, G., and Bryden, W. L. (1999). A comparison of ileal digesta and excreta analysis for the determination of amino acid digestibility in food ingredients for poultry. Br. Poult. Sci. 40 (2), 266–274. doi:10.1080/00071669987692
Ravindran, V., Hew, L. I., Ravindran, G., and Bryden, W. L. (2005). Apparent ileal digestibility of amino acids in dietary ingredients for broiler chickens. Animal Sci. 81 (1), 85–97. doi:10.1079/asc42240085
Rochell, S. J., Helmbrecht, A., Parsons, C. M., and Dilger, R. N. (2017). Interactive effects of dietary arginine and Eimeria acervulina infection on broiler growth performance and metabolism. Poult. Sci. 96 (3), 659–666. doi:10.3382/ps/pew295
Rodriguez, P. C., Quiceno, D. G., and Ochoa, A. C. (2007). L-arginine availability regulates T-lymphocyte cell-cycle progression. Blood 109 (4), 1568–1573. doi:10.1182/ blood-2006-06-031856
Rosenberg, M. M., and Zoebisch, O. C. (1952). A chick test for toxicity in forage legumes 1. Agron. J. 44 (6), 315–318. doi:10.2134/agronj1952.00021962004400060008x Ross-Broiler Nutrition Specifications 2022-EN (2022), Ross-Broiler Nutrition Specifications 2022-EN.
Ruan, D., Fouad, A. M., Fan, Q. L., Huo, X. H., Kuang, Z. X., Wang, H., et al. (2020). Dietary L-arginine supplementation enhances growth performance, intestinal antioxidative capacity, immunity and modulates gut microbiota in yellow-feathered chickens. Poult. Sci. 99 (12), 6935–6945. doi:10.1016/j.psj.2020.09.042
Rubin, L. L., Canal, C. W., Ribeiro, A., Kessler, A., Silva, I., Trevizan, L., et al. (2007). Effects of methionine and arginine dietary levels on the immunity of broiler chickens submitted to immunological stimuli. Braz. J. Poult. Sci. 9 (4), 241–247. doi:10.1590/ s1516-635x2007000400006
Sadeghi, G., Samie, A., Pourreza, J., and Rahmani, H. R. (2004). Canavanine content and toxicity of raw and treated bitter vetch (Vicia ervilia) seeds for broiler chicken. Int. J. Poult. Sci. 3 (8), 522–529. doi:10.3923/ijps.2004.522.529
Sadeghi, G. H., Mohammadi, L., Ibrahim, S. A., and Gruber, K. J. (2009a). Use of bitter vetch (Vicia ervilia) as a feed ingredient for poultry. Worlds Poult. Sci. J. 65 (1), 51–64. doi:10.1017/s004393390900004x
Sadeghi, G. H., Pourreza, J., Samei, A., and Rahmani, H. (2009b). Chemical composition and some anti-nutrient content of raw and processed bitter vetch (Vicia ervilia) seed for use as feeding stuff in poultry diet.Trop. Anim. Health Prod. 41, 85–93. doi:10.1007/s11250-008-9159-9
Sailer, M., Dahlhoff, C., Giesbertz, P., Eidens, M. K., de Wit, N., Rubio-Aliaga, I., et al. (2013). Increased plasma citrulline in mice marks diet-induced obesity and may predict the development of the metabolic syndrome. PloS one 8 (5), e63950. doi:10.1371/ journal.pone.0063950
San Martín, R., and Sobrevia, L. (2006). Gestational diabetes and the adenosine/ L-arginine/nitric oxide (ALANO) pathway in human umbilical vein endothelium. Placenta 27 (1), 1–10. doi:10.1016/j.placenta.2005.01.011
Satriano, J. (2004). Arginine pathways and the inflammatory response: interregulation of nitric oxide and polyamines: review article. Amino Acids 26 (4), 321–329. doi:10.1007/s00726-004-0078-4
Schriek, S. C. D., Staiger, E. K. P., and Michel, K. P. (2007). Bioinformatic evaluation of L-arginine catabolic pathways in 24 cyanobacteria and transcriptional analysis of genes encoding enzymes of L-arginine catabolism in the cyanobacterium Synechocystis sp. PCC 6803. BMC Genomics 8, 437–465. doi:10.1186/1471-2164-8-437
Seiler, N. (1987). Functions of polyamine acetylation. Can. J. Physiol. Pharmacol. 65 (10), 2024–2035. doi:10.1139/y87-317
Shapouri-Moghaddam, A., Mohammadian, S., Vazini, H., Taghadosi, M., Esmaeili, S., Mardani, F., et al. (2018). Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 233 (9), 6425–6440. doi:10.1002/jcp.26429
Sica, A., and Mantovani, A. (2012). Macrophage plasticity and polarization: in vivo veritas. J. Clin. Invest 122 (3), 787–795. doi:10.1172/JCI59643
Sirathonpong, O., Ruangpanit, Y., Songserm, O., Koo, E. J., and Attamangkune, S. (2019). Determination of the optimum arginine: lysine ratio in broiler diets. Animal Prod. Sci. 59 (9), 1705–1710. doi:10.1071/an18049
Srinongkote, S., Smriga, M., and Toride, Y. (2004). Diet supplied with L-lysine and Larginine during chronic stress of high stock density normalizes growth of broilers. Animal Sci. J. 75 (4), 339–343. doi:10.1111/j.1740-0929.2004.00195.x
Stuehr, D. J., Santolini, J., Wang, Z., Wei, C., and Adak, S. (2004). Update on mechanism and catalytic regulation in the NO synthases. J. Biol. Chem. 279 (35), 36167–36170. doi:10.1074/jbc.R400017200
Tamir, H., and Ratner, S. (1963a). Enzymes of arginine metabolism in chicks. Arch. Biochem. Biophys. 102 (2), 249–258. doi:10.1016/0003-9861(63)90178-4
Tamir, H., and Ratner, S. (1963b). A study of ornithine, citrulline and arginine synthesis in growing chicks. Arch. Biochem. Biophys. 102 (2), 259–269. doi:10.1016/ 0003-9861(63)90179-6
Tan, B., Li, X. G., Kong, X., Huang, R., Ruan, Z., Yao, K., et al. (2009). Dietary L-arginine supplementation enhances the immune status in early-weaned piglets. Amino Acids 37, 323–331. doi:10.1007/s00726-008-0155-1
Tan, B., Yin, Y., Kong, X., Li, P., Li, X., Gao, H., et al. (2010). L-Arginine stimulates proliferation and prevents endotoxin-induced death of intestinal cells. Amino Acids 38 (4), 1227–1235. doi:10.1007/s00726-009-0334-8
Tan, J., Applegate, T. J., Liu, S., Guo, Y., and Eicher, S. D. (2014). Supplemental dietary L-arginine attenuates intestinal mucosal disruption during a coccidial vaccine challenge in broiler chickens. Br. J. Nutr. 112 (7), 1098–1109. doi:10.1017/S0007114514001846
Tapeh, R. S., Zhandi, M., Zaghari, M., and Akhlaghi, A. (2017). Effects of guanidinoacetic acid diet supplementation on semen quality and fertility of broiler breeder roosters. Theriogenology 89, 178–182. doi:10.1016/j.theriogenology.2016.11.012
Torras-Llort, M., Torrents, D., Soriano-Garcia, J. F., Gelpi, J. L., Estévez, R., Ferrer, R., et al. (2001). Sequential amino acid exchange across b(0,+)-like system in chicken brush border jejunum. J. Membr. Biol. 180 (3), 213–220. doi:10.1007/s002320010072
Tossenberger, J., Rademacher, M., Németh, K., Halas, V., and Lemme, A. (2016). Digestibility and metabolism of dietary guanidino acetic acid fed to broilers. Poult. Sci. 95 (9), 2058–2067. doi:10.3382/ps/pew083
Uyanga, V. A., Jiao, H., Zhao, J., Wang, X., and Lin, H. (2020). Dietary L-citrulline supplementation modulates nitric oxide synthesis and anti-oxidant status of laying hens during summer season. J. Animal Sci. Biotechnol. 11, 103–116. doi:10.1186/s40104-020- 00507-5
Uyanga, V. A., Liu, L., Zhao, J., Wang, X., Jiao, H., and Lin, H. (2022). Central and peripheral effects of L-citrulline on thermal physiology and nitric oxide regeneration in broilers. Poult. Sci. 101 (3), 101669. doi:10.1016/j.psj.2021.101669
Uyanga, V. A., Sun, L., Liu, Y., Zhang, M., Zhao, J., Wang, X., et al. (2023). Effects of arginine replacement with L-citrulline on the arginine/nitric oxide metabolism in chickens: an animal model without urea cycle. J. Animal Sci. Biotechnol. 14 (1), 9–19. doi:10.1186/s40104-022-00817-w
van Meijl, L. E., Popeijus, H. E., and Mensink, R. P. (2010). Amino acids stimulate Akt phosphorylation, and reduce IL-8 production and NF-κB activity in HepG2 liver cells. Mol. Nutr. food Res. 54 (11), 1568–1573. doi:10.1002/mnfr.200900438
Vraneš, M., Ostojić, S., Tot, A., Papović, S., and Gadžurić, S. (2017). Experimental and computational study of guanidinoacetic acid self-aggregation in aqueous solution. Food Chem. 237, 53–57. doi:10.1016/j.foodchem.2017.05.088
Wang, J., Chen, L., Li, P., Li, X., Zhou, H., Wang, F., et al. (2008). Gene expression is altered in piglet small intestine by weaning and dietary glutamine supplementation. J. Nutr. 138 (6), 1025–1032. doi:10.1093/jn/138.6.1025
Watford, M. (2008). Glutamine metabolism and function in relation to proline synthesis and the safety of glutamine and proline supplementation. J. Nutr. 138 (10), 2003S–2007S. doi:10.1093/jn/138.10.2003S
Wentzel, A. S. No title. Polarized innate immunity: conservation of macrophage polarization in carp 2020.
Wolfson, R. L., Chantranupong, L., Saxton, R. A., Shen, K., Scaria, S. M., Cantor, J. R., et al. (2016). Sestrin2 is a leucine sensor for the mTORC1 pathway. Science 351 (6268), 43–48. doi:10.1126/science.aab2674
Wolfson, R. L., and Sabatini, D. M. (2017). The dawn of the age of amino acid sensors for the mTORC1 pathway. Cell metab. 26 (2), 301–309. doi:10.1016/j.cmet.2017.07.001
Wu, G. (2009). Amino acids: metabolism, functions, and nutrition. Amino Acids 37 (1), 1–17. doi:10.1007/s00726-009-0269-0
Wu, G. (2010). Functional amino acids in growth, reproduction, and health. Adv. Nutr. 1 (1), 31–37. doi:10.3945/an.110.1008
Wu, G., Bazer, F. W., Davis, T. A., Kim, S. W., Li, P., Marc Rhoads, J., et al. (2009). Arginine metabolism and nutrition in growth, health and disease. Amino Acids 37 (1), 153–168. doi:10.1007/s00726-008-0210-y
Wu, G., and Morris, S. M., Jr (1998). Arginine metabolism: nitric oxide and beyond. Biochem. J. 336 (1), 1–17. doi:10.1042/bj3360001
Zampiga, M., Laghi, L., Petracci, M., Zhu, C., Meluzzi, A., Dridi, S., et al. (2018). Effect of dietary arginine to lysine ratios on productive performance, meat quality, plasma and muscle metabolomics profile in fast-growing broiler chickens. J. animal Sci. Biotechnol. 9, 79–14. doi:10.1186/s40104-018-0294-5
Zhang, B., Gan, L., Shahid, M. S., Lv, Z., Fan, H., Liu, D., et al. (2019). In vivo and in vitro protective effect of arginine against intestinal inflammatory response induced by Clostridium perfringens in broiler chickens. J. animal Sci. Biotechnol. 10, 73–14. doi:10. 1186/s40104-019-0371-4
Zhang, B., Lv, Z., Li, H., Guo, S., Liu, D., and Guo, Y. (2017). Dietary l-arginine inhibits intestinal Clostridium perfringens colonisation and attenuates intestinal mucosal injury in broiler chickens. Br. J. Nutr. 118 (5), 321–332. doi:10.1017/S0007114517002094
Zhang, B., Lv, Z., Li, Z., Wang, W., Li, G., and Guo, Y. (2018). Dietary L-arginine supplementation alleviates the intestinal injury and modulates the gut microbiota in broiler chickens challenged by Clostridium perfringens. Front. Microbiol. 9, 1716. doi:10. 3389/fmicb.2018.01716
Zheng, P., Yu, B., He, J., Yu, J., Mao, X., Luo, Y., et al. (2017). Arginine metabolism and its protective effects on intestinal health and functions in weaned piglets under oxidative stress induced by diquat. Br. J. Nutr. 117 (11), 1495–1502. doi:10.1017/ S0007114517001519





Arginine is a powerful vasodilator which increases blood circulation and also helps in modulation of protective immune response. Under heat stress conditions arginine has the ability to alleviate heat induced stress & normalize the growth performance.








.jpg&w=3840&q=75)