Egg production and quality responses to increasing isoleucine supplementation in Shaver white hens fed a low crude protein corn-soybean meal diet fortified with synthetic amino acids between 20 and 46 weeks of age
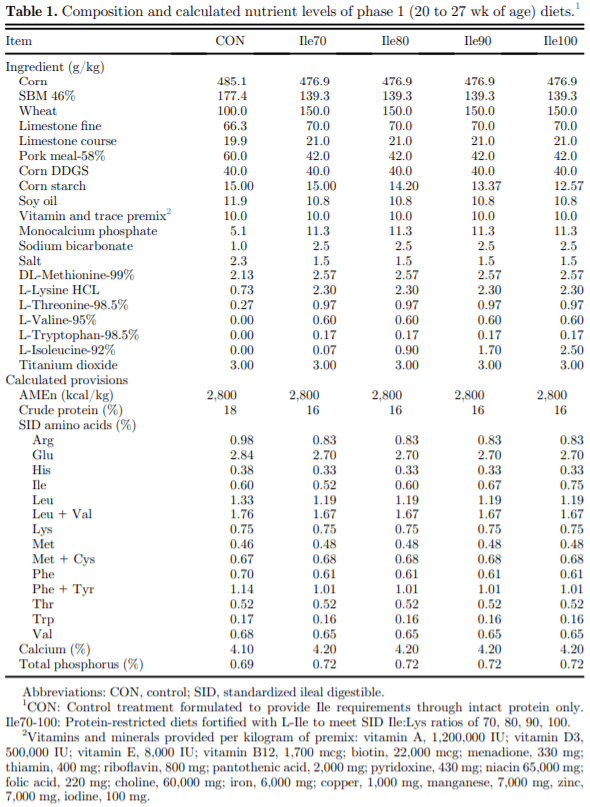
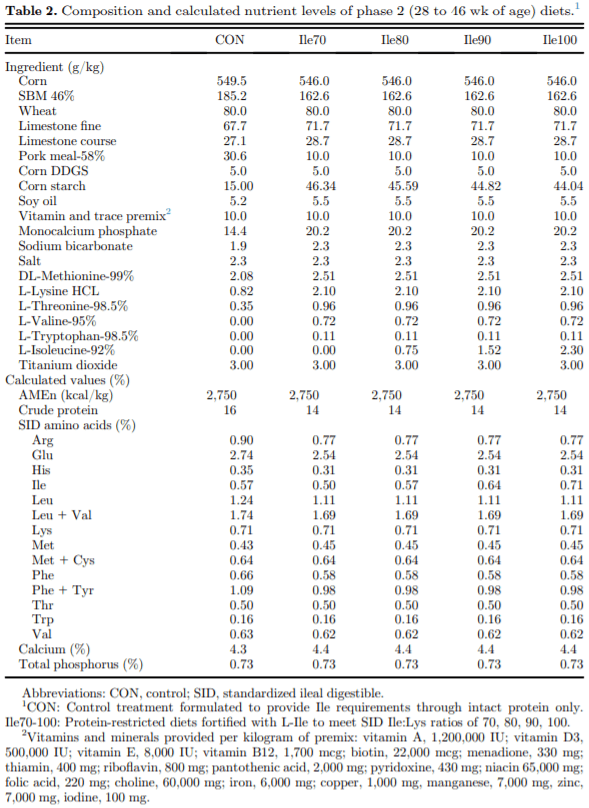
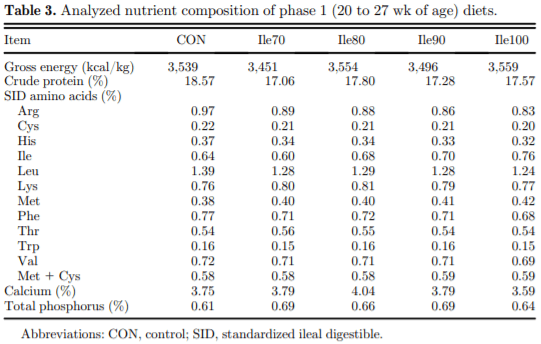
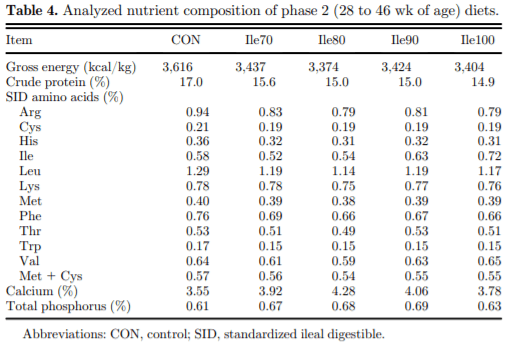
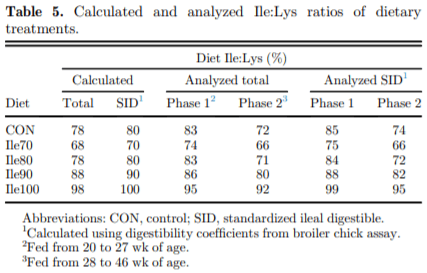
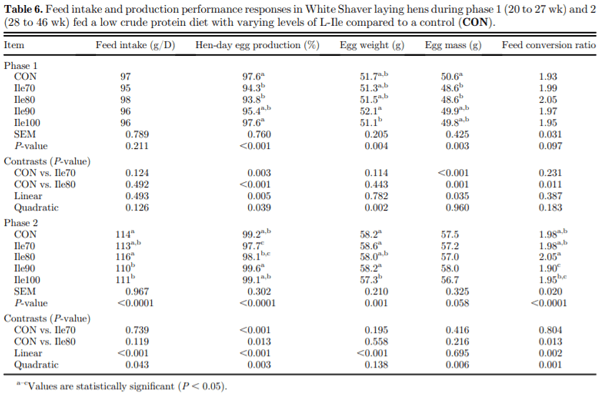
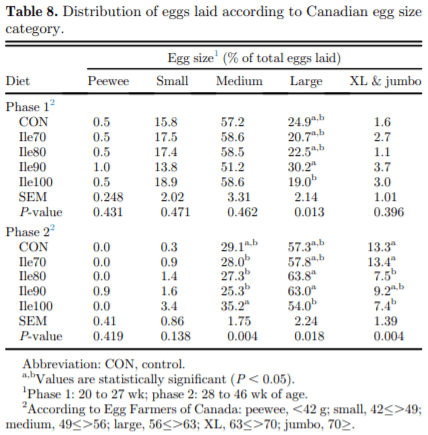
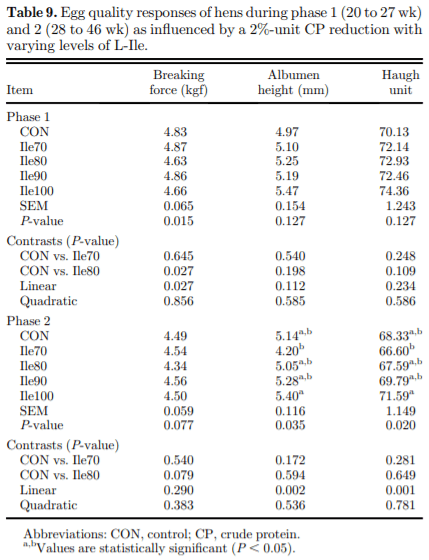
The present study evaluated production performance responses to Ile supplementation in laying hens fed low crude protein (LCP), amino acid (AA) balanced diets. A total of 179 Shaver white pullets were distributed into 30 battery cages (6 birds/cage, n 5 6) and observed over the course of 27 wk in a 2-phase (20 to 27 and 28 to 46 wk of age) feeding program. Five isocaloric diets were formulated for standardized ileal digestible (SID) Lys intake of 750 and 710 mg/D in phase 1 and 2, respectively, and included a positive control with standard levels of crude protein (CP) (CON; 18 and 16% CP for phases 1 and 2), and 4 LCP diets (16 and 14% CP for phase 1 and 2, respectively) with graded levels of Ile to satisfy SID Ile:Lys ratios of 70 (Ile70), 80 (Ile80), 90 (Ile90), and 100% (Ile100). Based on analyzed dietary AA, the calculated SID Ile:Lys of LCP diets were 75, 84, 88, 99% and 66, 72, 82, 95% for phase 1 and 2, respectively. Dietary treatments significantly (P, 0.05) affected feed intake, hen-day egg production (HDEP), egg weight (EW), feed conversion ratio, and egg quality (Haugh unit) and composition (yolk to albumen). Lowering dietary CP negatively affected HDEP with a 3.3 and 1.5% reduction in phase 1 and 2, respectively, and this was restored with the addition of Ile (P, 0.001) suggesting that Ile was limiting in the LCP basal diet. Average EW was reduced in Ile100 only; however, the Ile:Lys appeared to influence egg size uniformity, with Ile90 producing a greater proportion of large (56 g EW . 63 g) eggs, suggesting that Ile may be used to manipulate EW at the expense of HDEP. Overall, the results indicated that CP in laying hen diets can be reduced by 2% units if fortified with synthetic AA (Met, Lys, Thr, Trp) 1 Ile, with optimal responses observed between 82 and 88% SID Ile:Lys.
Key words: laying hen, isoleucine, low crude protein, egg production and quality.
Adedokun, S., P. Utterback, C. Parsons, O. Adeola, M. Lilburn, and T. Applegate. 2009. Comparison of amino acid digestibility of feed ingredients in broilers, laying hens and caecectomised roosters. Br. Poult. Sci. 50:350–358.
Adedokun, S., P. Jaynes, M. Adb El-Hack, R. Payne, and T. Applegate. 2014. Standardized ileal amino acid digestibility of meat and bone meal and soybean meal in laying hens and broilers. Poult. Sci. 93:420–428.
Adedokun, S., P. Jaynes, R. Payne, and T. Applegate. 2015. Stan-dardized ileal amino acid digestibility of corn, corn distillers’ dried grains with solubles, wheat middlings, and bakery by-products in broilers and laying hens. Poult. Sci. 94:2480–2487.
Bain, M. M., Y. Nys, and I. C. Dunn. 2016. Increasing persistency in lay and stabilising egg quality in longer laying cycles. What are the challenges? Br. Poult. Sci. 57:330–338.
Baker, D., A. Batal, T. Parr, N. Augspurger, and C. Parsons. 2002. Ideal ratio (relative to lysine) of tryptophan, threonine, isoleucine, and valine for chicks during the second and third weeks posthatch. Poult. Sci. 81:185–494.
Berres, J., S. L. Vieira, M. T. Kidd, D. Taschetto, D. M. Freitas, R. Barros, and E. T. Nogueira. 2010. Supplementing l-valine and l-isoleucine in low-protein corn and soybean meal all-vegetable diets for broilers. J. Appl. Poult. Res. 19:373–379.
Blair, R., J. Jacob, S. Ibrahim, and P. Wang. 1999. A quantitative assessment of reduced protein diets and supplements to improve nitrogen utilization. J. Appl. Poult. Res. 8:25–47.
Bray, D. J. 1964. Studies with corn-soya laying diets: 7. Limiting amino acids in a 60:40 blend of corn and soybean protein. Poult. Sci. 43:396–401.
Bray, D. J. 1969. Studies with corn-soya laying diets 8. Requirements for limiting amino acids—the basal diet and the requirements for isoleucine, lysine and tryptophan. Poult. Sci. 48:674–684.
Bregendahl, K., S. Roberts, B. Kerr, and D. Hoehler. 2008. Ideal ratios of isoleucine, methionine, methionine plus cystine, threonine, tryptophan, and valine relative to lysine for white leghorn-type laying hens of twenty-eight to thirty-four weeks of age. Poult. Sci. 87:744–758.
Burley, H., P. Patterson, and M. Elliot. 2013. Effect of a reduced crude protein, amino acid-balanced diet on hen performance, production costs, and ammonia emissions in a commercial laying hen flock. J. Appl. Poult. Res. 22:217–228.
Canadian Council on Animal Care (CCAC). 2009. CCAC guidelines on: the care and use of farm animals in research, teaching and testing. Accessed Mar. 2018. Ottawa ON, Canada. https://www. ccac.ca/Documents/Standards/Guidelines/Farm_Animals.pdf.
Casartelli, E., R. Filardi, O. Junqueira, A. Laurentiz, V. Assuena, and K. Duarte. 2005. Commercial laying hen diets formulated accord-ing to different recommendations of total and digestible amino acids. Braz. J. Poult. Sci. 7:177–180.
Centraal Veevoederbureau (CVB). 1996. Aminozurenbohoefte van leghennen en vleeskuikens [Amino Acid Requirements for Laying Hens and Broiler Chickens]. Centraal Veevoederbureau, Lelystad, the Netherlands.
Coon, C., and B. Zhang. 1999. Ideal amino acid profile for layers examined. Feedstuffs 71:13–15.
Corzo, A., R.E. Loar, II, and M. T. Kidd. 2009. Limitations of dietary isoleucine and valine in broiler chick diets1. Poult. Sci. 88:1934–1938.
Dong, X. Y., M. M. M. Azzam, and X. T. Zou. 2016. Effects of dietary L-isoleucine on laying performance and immunomodulation of laying hens. Poult. Sci. 95:2297–2305.
Egg Farmers of Canada. 2017. Egg Farmers of Canada annual report. Accessed Jan. 2019. https://www.eggfarmers.ca/wp-content/ uploads/2018/03/2017_Egg-Farmers-of-Canada-Annual-Report. pdf.
Egg Farmers of Canada. 2018. Egg Farmers of Canada annual report. Accessed June 2019. https://www.eggfarmers.ca/wp-content/ uploads/2019/03/2018_Egg-Farmers-of-Canada-Annual-Report. pdf.
Elliot, M. 2008. Amino acid nutrition of commercial pullets and layers. Pages 193–221 in Proceedings of Intermountain Nutrition Con-ference. Utah Agricultural Experiment Station, Salt Lake City, UT, pp. 193–221.
Harms, R., and F. Ivey. 1993. Performance of commercial laying hens fed various supplemental amino acids in a corn-soybean meal diet. J. Appl. Poult. Res. 2:273–282.
Harms, R., and G. Russel. 1993. Optimizing egg mass with amino acid supplementation of a low-protein diet. Poult. Sci. 72:1892–1896.
Harms, R. H., and G. B. Russell. 2000. Evaluation of the isoleucine requirement of the commercial layer in a corn-soybean meal diet. Poult. Sci. 79:1154–1157.
Hendrix ISA. 2011. Nutrition Management Guide. 2011 ed. ISA Hendrix Genetics, Saint Brieuc, France.
Huang, K., X. Li, V. Ravindran, and W. Bryden. 2006. Comparison of apparent ileal amino acid digestibility of feed ingredients measured with broilers, layers and roosters. Poult. Sci. 85:625–634.
Huyghebaert, G., G. De Groote, E. A. Butler, and T. R. Morris. 1991. Optimum isoleucine requirement of laying hens and the effect of age. Br. Poult. Sci. 32:471–481.
Jensen, L. S. 1991. Protein and amino acid nutrition of laying hens and broilers. Pages 159–170 in Proceedings of the 26th Annual Northwest Animal Nutrition Conference. Henschel, K.K., ed. Beaverton, OR.
Ji, F., S. Fu, B. Ren, S. Wu, H. Zhang, H. Yue, J. Gao, A. Helmbrecht, and G. Qi. 2014. Evaluation of amino-acid supplemented diets varying in protein levels for laying hens. J. Appl. Poult. Res. 23:384–392.
Keshavarz, K., and M. Jackson. 1992. Performance of growing pullets and laying hens fed low-protein, amino acid-supplemented diets. Poult. Sci. 71:905–918.
Kim, E. J., P. L. Utterback, T. J. Applegate, and C. M. Parsons. 2011. Comparison of amino acid digestibility of feedstuffs determined with the precision-fed cecectomized rooster assay and the stan-dardized ileal amino acid digestibility assay. Poult. Sci. 90:2511– 2519.
Kim, E. J., P. L. Utterback, and C. M. Parsons. 2012a. Comparison of amino acid digestibility coefficients for soybean meal, canola meal, fish meal, and meat and bone meal among 3 different bioassays. Poult. Sci. 91:1350–1355.
Kim, E. J., P. L. Utterback, and C. M. Parsons. 2012b. Comparison of amino acid digestibility coefficients for corn, corn gluten meal, and corn distillers dried grains with solubles among 3 different bio-assays. Poult. Sci. 91:3141–3147.
Kristensen, H., and C. Wathes. 2000. Ammonia and poultry welfare: a review. Worlds Poult. Sci. 56:235–245.
Leeson, S., and J. D. Summers. 2005. Commercial Poultry Production, 3rd ed. University Books, Guelph, ON.
de Lima, M., N. Sakomura, J. Dorigam, E. da Silva, N. Ferreira, and J. Fernandes. 2016. Maintenance valine, isoleucine and tryptophan requirements for poultry. Poult. Sci. 95:842–850.
de Lima, M., N. K. Sakomura, E. Silva, J. C. P. Dorigam, N. T. Ferreira, E. Malheiros, and J. Fernandes. 2018. The optimal digestible valine, isoleucine and tryptophan intakes of broiler breeder hens for rate of lay. Anim. Feed Sci. Technol. 238:29–38.
Mello, H. H. de C., P. C. Gomes, T. C. da Rocha, J. L. Donzele, R. L. de Almeida, A. R. Troni, B. R. de Carvalho, and G. da S. Viana. 2012. Determination of digestible isoleucine: lysine ratio in diets for laying hens aged 42-58 weeks. Rev. Bras. Zootec. 41:1313–1317.
Meluzzi, A., F. Sirri, N. Tallarico, and A. Franchini. 2001. Nitrogen retention and performance of brown laying hens on diets with different protein content and constant concentration of amino acids and energy. Br. Poult. Sci. 42:213–217.
Miller, E., M. Sunde, H. Bird, and C. Elvehjem. 1954. The isoleucine requirements of the laying hen. Poult. Sci. 33:1201–1209.
Morris, T., and R. Gous. 1988. Partitioning of the response to protein between egg number and egg weight. Br. Poult. Sci. 29:93–99.
Mutucumarana, R., V. Ravindran, G. Ravindran, and A. J. Cowieson. 2014. Influence of dietary calcium concentration on the digestion of nutrients along the intestinal tract of broiler chickens. J. Poult. Sci. 51:392–401.
Mwaniki, Z., M. Neijat, and E. Kiarie. 2018. Egg production and quality responses of adding up to 7.5% defatted black soldier fly larvae meal in a corn–soybean meal diet fed to Shaver White Leghorns from wk 19 to 27 of age. Poult. Sci. 97:2829–2835.
National Research Council. 1994. Nutrient Requirements of Poultry, 9th rev. ed. National Academy Press, Washington, DC.
Ospina Rojas, I., A. Murakami, J. Fanhani, K. Picoli, and M. Baptista Barbosa. 2015. Tryptophan, threonine and isoleucine supplemen-tation in low-protein diets for commercial laying hens. Semin. Ci^enc. Agrar. 36:1735–1744.
Peganova, S., F. Hirche, and K. Eder. 2003. Requirement of trypto-phan in relation to the supply of large neutral amino acids in laying hens. Poult. Sci. 82:815–822.
Roberts, S., H. Xin, B. Kerr, J. Russell, and K. Bregendahl. 2007. Effects of dietary fiber and reduced crude protein on nitrogen balance and egg production in laying hens. Poult. Sci. 86:1716–1725.
Rocha, T. C. da, J. L. Donzele, P. C. Gomes, H. H. de C. Mello, A. R. Troni, and G. da S. Viana. 2013. Ideal digestible isoleucine: digestible lysine ratio in diets for laying hens aged 24-40 weeks. Rev. Bras. Zootec. 42:780–784.
Shivazad, M., R. Harms, G. Russell, D. Faria, and R. Antar. 2002. Re-evaluation of the isoleucine requirement of the commercial layer. Poult. Sci. 81:1869–1872.
Sohail, S., M. Bryant, and D. Roland. 2002. Influence of supple-mental lysine, isoleucine, threonine, tryptophan and total sulfur amino acids on egg weight of Hyline W-36 hens. Poult. Sci. 81:1038–1044.
Summers, J. D. 1993. Reducing nitrogen excretion of the laying hen by feeding lower crude protein diets. Poult. Sci. 72:1473–1478.
Torki, M., M. Ahmad, H. A. Ghasemi, and A. Zardast. 2015. Response of laying hens to feeding low-protein amino acid-supplemented diets under high ambient temperature: performance, egg quality, leukocyte profile, blood lipids and excreta pH. Int. J. Biometeorol. 59:575–584.




Ilona Parenteau, As in the market we have synthetic lysine to valine, the study with isoleucine to define a better relationship with lysine is of fundamental importance, since this amino acid is the possible limiting factor after valine, so it is the isoleucine x lysine relationship that defines maximum level of protein reduction that can be used since there is no commercially available isoleucine. And of course, to use valine, this information is a reference.
As this amino acid is the limiting factor after valine. As a suggestion, I would like to comment on the results we obtained in studies conducted with laying hens, the ideal ratio between sulfur amino acids (MET + CIST) with lysine is 100: 100. Finally, I congratulate the authors for using a sub-optimal level of lysine, which is the most correct practice of determining the amino acid relationship between any amino acid and lysine.








.jpg&w=3840&q=75)