Calcium Nutrition of Broilers: Current Perspectives and Challenges

1. Underwood, E.; Suttle, N. The Mineral Nutrition of Livestock, 3rd ed.; CABI Publishing: Wallingford, UK, 1999; p. 614.
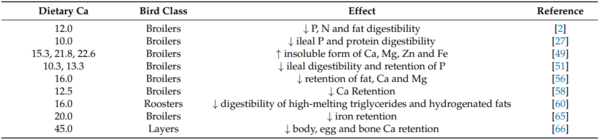
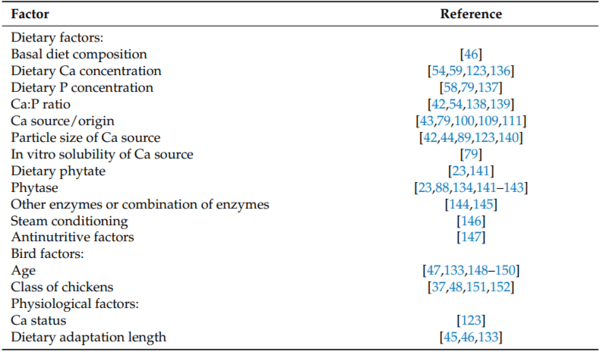
2. Mutucumarana, R.K.; Ravindran, V.; Ravindran, G.; Cowieson, A.J. Influence of Dietary Calcium Concentration on the Digestion of Nutrients along the Intestinal Tract of Broiler Chickens. J. Poult. Sci. 2014, 51, 392–401. [CrossRef]
3. Wilkinson, S.; Bradbury, E.; Thomson, P.; Bedford, M.; Cowieson, A. Nutritional geometry of calcium and phosphorus nutrition in broiler chicks. The effect of different dietary calcium and phosphorus concentrations and ratios on nutrient digestibility. Animal
2014, 8, 1080–1088. [CrossRef] [PubMed]
4. Mutucumarana, R.K. Measurement of True Ileal Phosphorous Digestibility in Feed Ingredients for Poultry. Ph.D. Thesis, Massey
University, Palmerston North, New Zealand, 2014.
5. Anwar, M.N. Measurement of True Ileal Calcium Digestibility of Feed Ingredients for Broiler Chickens. Ph.D. Thesis, Massey
University, Palmerston North, New Zealand, June 2017.
6. David, L.S. Studies on the Measurement of Calcium Digestibility in Raw Materials for Poultry and of Digestible Calcium
Requirement of Broiler Starters. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 30 September 2021.
7. van der Klis, J.D.; Verstegen, M.W. Phosphorous nutrition of poultry. In Recent Advances in Poultry Nutrition; Garnsworthy, P.C.,
Wiseman, J., Haresign, W., Eds.; Nottingham University Press: Nottingham, UK, 1996; pp. 71–83.
8. Veum, T.L. Phosphorus and calcium nutrition and metabolism. In Phosphorus and Calcium Utilization and Requirements in Farm
Animals; Vitti, D.M.S.S., Kebreab, E., Eds.; CABI Publishing: Wallingford, UK, 2010; pp. 94–111.
9. Adedokun, S.A.; Adeola, O. Calcium and phosphorus digestibility: Metabolic limits. J. Appl. Poult. Res. 2013, 22, 600–608.
[CrossRef]
10. Proszkowiec-Weglarz, M.; Angel, R. Calcium and phosphorous metabolism in broilers: Effect of homeostatic mechanism on calcium and phosphorous digestibility. J. Appl. Poult. Res. 2013, 22, 609–627. [CrossRef]
11. Etches, R.J. Calcium Logistics in the Laying Hen. J. Nutr. 1987, 117, 619–628. [CrossRef]
12. Whitehead, C.C. Overview of bone biology in the egg-laying hen. Poult. Sci. 2004, 83, 193–199. [CrossRef]
13. de Vries, S.; Kwakkel, R.P.; Dijkstra, J. Dynamics of calcium and phosphorus metabolism in laying hens. In Phosphorus and
Calcium Utilization and Requirements in Farm Animals; Vitti, D.M.S.S., Kebreab, E., Eds.; CABI Publishing: Wallingford, UK, 2010; pp. 133–150.
14. Nys, Y.; Guyot, N. Egg formation and chemistry. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., van Immerseel, F., Eds.; Woodhead Publishing, Elsevier Inc.: Duxford, UK, 2011; pp. 83–132.
15. Kim, W.; Bloomfield, S.; Sugiyama, T.; Ricke, S. Concepts and methods for understanding bone metabolism in laying hens. World’s
Poult. Sci. J. 2012, 68, 71–82. [CrossRef]
16. Korver, D.R. Calcium nutrition, bone metabolism, and eggshell quality in longer-persisting layer flocks. In Proceedings of the
31st Annual Australian Poultry Science Symposium, Sydney, Australia, 16–19 February 2020.
17. Billman, G.E. Homeostasis: The Underappreciated and Far Too Often Ignored Central Organizing Principle of Physiology. Front.
Physiol. 2020, 11, 200. [CrossRef]
18. Coon, C.; Leske, K.; Seo, S. The availability of calcium and phosphorus in feedstuffs. In Poultry Feedstuffs: Supply, Composition, and
Nutritive Value; McNab, J.M., Boorman, N., Eds.; CABI Publishing: New York, NY, USA, 2002; Volume 26, pp. 151–186.
19. Hurwitz, S.; Bar, A. The Sites of Calcium and Phosphate Absorption in the Chick. Poult. Sci. 1970, 49, 324–325. [CrossRef]
[PubMed]
20. Hurwitz, S.; Bar, A. Calcium and Phosphorus Interrelationships in the Intestine of the Fowl. J. Nutr. 1971, 101, 677–685. [CrossRef]
21. van der Klis, J.D.; Verstegen, M.; de Wit, W. Absorption of Minerals and Retention Time of Dry Matter in the Gastrointestinal
Tract of Broilers. Poult. Sci. 1990, 69, 2185–2194. [CrossRef]
22. Guinotte, F.; Gautron, J.; Nys, Y.; Soumarmon, A. Calcium solubilisation and retention in the gastrointestinal tract in chicks (Gallus domesticus) as a function of gastric acid secretion inhibition and of calcium carbonate particle size. Br. J. Nutr. 1995,
73, 125–139. [CrossRef] [PubMed]
23. Selle, P.H.; Cowieson, A.J.; Ravindran, V. Consequences of calcium interactions with phytate and phytase for poultry and pigs.
Livest. Sci. 2009, 124, 126–141. [CrossRef]
24. Dersjant-Li, Y.; Evans, C.; Kumar, A. Effect of phytase dose and reduction in dietary calcium on performance, nutrient digestibility, bone ash and mineralization in broilers fed corn-soybean meal-based diets with reduced nutrient density. Anim. Feed. Sci. Technol.
2018, 242, 95–110. [CrossRef]
25. Tancharoenrat, P.; Ravindran, V. Influence of tallow and calcium concentrations on the performance and energy and nutrient utilisation in broiler starters. Poult. Sci. 2014, 93, 1453–1462. [CrossRef]
26. Thomas, D.V.; Ravindran, V. Mineral Retention in Young Broiler Chicks Fed Diets Based on Wheat, Sorghum or Maize. AsianAustralas. J. Anim. Sci. 2009, 23, 68–73. [CrossRef]
27. Akter, M.M.; Graham, H.; Iji, P.A. Influence of different levels of calcium, non-phytate phosphorus and phytase on apparent metabolizable energy, nutrient utilization, plasma mineral concentration and digestive enzyme activities of broiler chickens. J.
Appl. Anim. Res. 2017, 46, 278–286. [CrossRef]
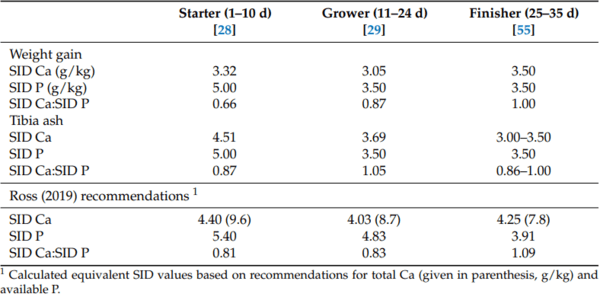
28. David, L.; Abdollahi, M.; Bedford, M.; Ravindran, V. Requirement of digestible calcium at different dietary concentrations of digestible phosphorus for broiler chickens. 1. Broiler starters (d 1 to 10 post-hatch). Poult. Sci. 2021, 100, 101439. [CrossRef]
[PubMed]
29. David, L.S.; Abdollahi, M.R.; Bedford, M.R.; Ravindran, V. Requirement of digestible calcium at different dietary concentrations of digestible phosphorus for broiler chickens. 2. Broiler growers (d 11 to 24 post-hatch). Poult. Sci. 2022, 101, 102135. [CrossRef]
30. Bronner, F. Calcium Absorption—A Paradigm for Mineral Absorption. J. Nutr. 1998, 128, 917–920. [CrossRef]
31. Vitti, D.M.; Silva Fiho, S.S.; Louvandini, J.C.D.; Dias, H.; Bueno, R.S.; Kebreab, E. Phosphorus and calcium utilization in ruminants using isotope dilution technique. In Phosphorus and Calcium Utilization and Requirements in Farm Animals; Vitti, D.M.S.S.,
Kebreab, E., Eds.; CABI Publishing: Wallingford, UK, 2010; pp. 45–67.
32. Rath, N.C.; Huff, G.R.; Huff, W.E.; Balog, J.M. Factors Regulating Bone Maturity and Strength in Poultry. Poult. Sci. 2000,
79, 1024–1032. [CrossRef]
33. Ravindran, V. Progress in ileal endogenous amino acid flow research in poultry. J. Anim. Sci. Biotechnol. 2021, 12, 1–11. [CrossRef]
34. Anwar, M.N.; Ravindran, V. Influence of methodology on the measurement of ileal endogenous calcium losses in broiler chickens.
J. Appl. Anim. Res. 2020, 48, 264–267. [CrossRef]
35. Tancharoenrat, P.; Ravindran, V.; Zaefarian, F.; Ravindran, G. Digestion of fat and fatty acids along the gastrointestinal tract of broiler chickens. Poult. Sci. 2014, 93, 371–379. [CrossRef] [PubMed]
36. Barua, M.; Abdollahi, M.; Zaefarian, F.; Wester, T.; Girish, C.; Chrystal, P.; Ravindran, V. Basal ileal endogenous amino acid flow in broiler chickens as influenced by age. Poult. Sci. 2021, 100, 101269. [CrossRef] [PubMed]
37. Mtei, A.; Abdollahi, M.; Schreurs, N.; Girish, C.; Ravindran, V. Dietary inclusion of fibrous ingredients and bird type influence apparent ileal digestibility of nutrients and energy utilization. Poult. Sci. 2019, 98, 6702–6712. [CrossRef]
38. Bronner, F.; Pansu, D. Nutritional Aspects of Calcium Absorption. J. Nutr. 1999, 129, 9–12. [CrossRef]
39. Jones, G.; Strugnell, S.A.; DeLUCA, H.F. Current Understanding of the Molecular Actions of Vitamin D. Physiol. Rev. 1998,
78, 1193–1231. [CrossRef]
40. Walk, C.L. The influence of calcium on phytase efficacy in non-ruminant animals. Anim. Prod. Sci. 2016, 56, 1345. [CrossRef]
41. Ravindran, V.; Abdollahi, M.R.; Bootwalla, S.M. Nutrient analysis, metabolizable energy, and digestible amino acids of soybean meals of different origins for broilers. Poult. Sci. 2014, 93, 2567–2577. [CrossRef] [PubMed]
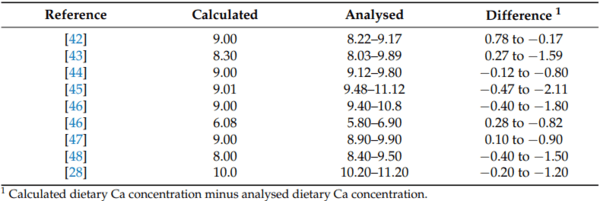
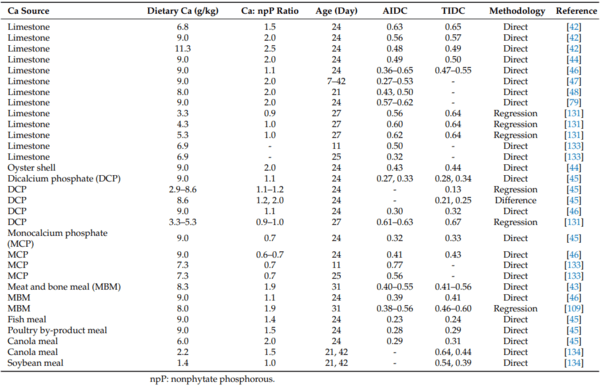
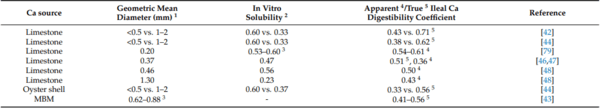
42. Anwar, M.N.; Ravindran, V.; Morel, P.C.H.; Ravindran, G.; Cowieson, A.J. Effect of limestone particle size and calcium to non-phytate phosphorus ratio on true ileal calcium digestibility of limestone for broiler chickens. Br. Poult. Sci. 2016, 57, 707–713.
[CrossRef]
43. Anwar, M.N.; Ravindran, V.; Morel, P.C.H.; Ravindran, G.; Cowieson, A.J. Measurement of true ileal calcium digestibility in meat and bone meal for broiler chickens using the direct method. Poult. Sci. 2016, 95, 70–76. [CrossRef]
44. Anwar, M.; Ravindran, V.; Morel, P.; Ravindran, G.; Cowieson, A. Effect of calcium source and particle size on the true ileal digestibility and total tract retention of calcium in broiler chickens. Anim. Feed. Sci. Technol. 2017, 224, 39–45. [CrossRef]
45. Anwar, M.; Ravindran, V.; Morel, P.; Cowieson, A. Measurement of the True Ileal Calcium Digestibility of Some Feed Ingredients for Broiler Chickens. Anim. Feed. Sci. Technol. 2018, 237, 118–128. [CrossRef]
46. David, L.S.; Abdollahi, M.R.; Ravindran, G.; Walk, C.L.; Ravindran, V. Studies on the measurement of ileal calcium digestibility of calcium sources in broiler chickens. Poult. Sci. 2019, 98, 5582–5589. [CrossRef] [PubMed]
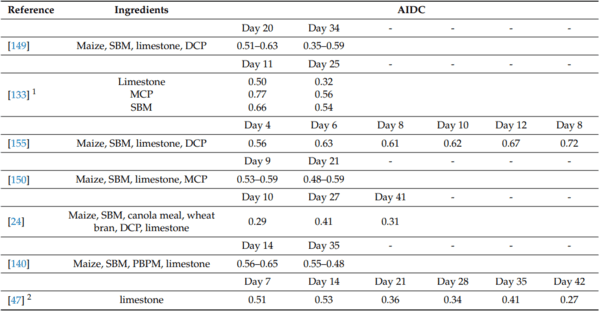
47. David, L.S.; Abdollahi, M.R.; Bedford, M.; Ravindran, V. Effect of age and dietary crude protein content on the apparent ileal calcium digestibility of limestone in broiler chickens. Anim. Feed. Sci. Technol. 2020, 263, 114468. [CrossRef]
48. David, L.S.; Abdollahi, M.R.; Bedford, M.R.; Ravindran, V. Comparison of the apparent ileal calcium digestibility of limestone in broilers and layers. Br. Poult. Sci. 2021, 62, 852–857. [CrossRef]
49. Shafey, T.M.; McDonald, M.W.; Dingle, J.G. Effects of dietary calcium and available phosphorus concentration on digesta pH and on the availability of calcium, iron, magnesium and zinc from the intestinal contents of meat chickens. Br. Poult. Sci. 1991,
32, 185–194. [CrossRef] [PubMed]
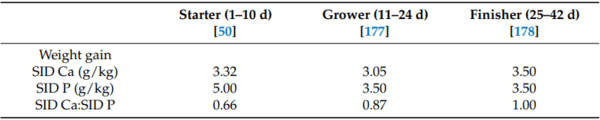
50. Walk, C.; Wang, Z.; Wang, S.; Wu, J.; Sorbara, J.; Zhang, J. Determination of the standardized ileal digestible calcium requirement of male Arbor Acres Plus broilers from hatch to day 10 post-hatch. Poult. Sci. 2021, 100, 101364. [CrossRef] [PubMed]
51. Abdollahi, M.; Duangnumsawang, Y.; Kwakkel, R.; Steenfeldt, S.; Bootwalla, S.; Ravindran, V. Investigation of the interaction between separate calcium feeding and phytase supplementation on growth performance, calcium intake, nutrient digestibility and energy utilisation in broiler starters. Anim. Feed. Sci. Technol. 2016, 219, 48–58. [CrossRef]
52. Tamim, N.M.; Angel, R. Phytate Phosphorus Hydrolysis As Influenced by Dietary Calcium and Micro-Mineral Source in Broiler
Diets. J. Agric. Food Chem. 2003, 51, 4687–4693. [CrossRef] [PubMed]
53. Tamim, N.; Angel, R.; Christman, M. Influence of dietary calcium and phytase on phytate phosphorus hydrolysis in broiler chickens. Poult. Sci. 2004, 83, 1358–1367. [CrossRef]
54. Plumstead, P.; Leytem, A.; Maguire, R.; Spears, J.; Kwanyuen, P.; Brake, J. Interaction of Calcium and Phytate in Broiler Diets. 1.
Effects on Apparent Prececal Digestibility and Retention of Phosphorus. Poult. Sci. 2008, 87, 449–458. [CrossRef] [PubMed]
55. David, L.; Abdollahi, M.; Bedford, M.; Ravindran, V. Requirement of digestible calcium at different dietary concentrations of digestible phosphorus for broiler chickens 3. Broiler finishers (d 25 to 35 post-hatch). Poult. Sci. 2023, 102, 102492. [CrossRef]
56. Atteh, J.O.; Leeson, S. Effects of Dietary Saturated or Unsaturated Fatty Acids and Calcium Levels on Performance and Mineral
Metabolism of Broiler Chicks. Poult. Sci. 1984, 63, 2252–2260. [CrossRef]
57. Atteh, J.O.; Leeson, S. Effects of Dietary Fat Level on Laying Hens Fed Various Concentrations of Calcium. Poult. Sci. 1985,
64, 2090–2097. [CrossRef] [PubMed]
58. Sebastian, S.; Touchburn, S.P.; Chavez, E.R.; Lague, P.C. Efficacy of supplemental microbial phytase at different dietary calcium levels on growth performance and mineral utilisation of broiler chickens. Poult. Sci. 1996, 75, 1516–1523. [CrossRef] [PubMed]
59. Gautier, A.E.; Walk, C.L.; Dilger, R.N. Influence of dietary calcium concentrations and the calcium-to-non-phytate phosphorus ratio on growth performance, bone characteristics, and digestibility in broilers. Poult. Sci. 2017, 96, 2795–2803. [CrossRef]
60. Edwards, H.M., Jr.; Dunahoo, W.S.; Carmon, J.L.; Fuller, H.L. Effect of protein, energy and fat content of the ration on calcium utilisation. Poult. Sci. 1960, 39, 1389–1394. [CrossRef]
61. Atteh, J.O.; Leeson, S. Effects of Dietary Fatty Acids and Calcium Levels on Performance and Mineral Metabolism of Broiler
Chickens. Poult. Sci. 1983, 62, 2412–2419. [CrossRef]
62. Davis, G.K. Effects of high calcium intakes on the absorption of other nutrients. Fed. Proc. 1959, 18, 1119–1124.
63. Shafey, T.M.; McDonald, M.W. The effects of dietary calcium, phosphorus, and protein on the performance and nutrient utilisation of broiler chickens. Poult. Sci. 1991, 70, 548–553. [CrossRef]
64. Ogura, Y. Leg abnormality associated with dietary excessive calcium carbonate in broiler chicks. Natl. Inst. Anim. Health Q. 1981,
21, 141–147.
65. Sell, J.L. Utilisation of iron by the chick as influenced by dietary calcium and phosphorus. Poult. Sci. 1965, 44, 550. [CrossRef]
66. Kebreab, E.; France, J.; Kwakkel, R.; Leeson, S.; Kuhi, H.D.; Dijkstra, J. Development and evaluation of a dynamic model of calcium and phosphorus flows in layers. Poult. Sci. 2009, 88, 680–689. [CrossRef] [PubMed]
67. Lawlor, P.G.; Lynch, P.B.; Caffrey, P.J.; O’Reilly, J.J.; O’Connell, M.K. Measurements of the acid-binding capacity of ingredients used in pig diets. Ir. Veter. J. 2005, 58, 447–452. [CrossRef] [PubMed]
68. Jasaitis, D.; Wohlt, J.; Evans, J. Influence of Feed Ion Content on Buffering Capacity of Ruminant Feedstuffs In Vitro. J. Dairy Sci.
1987, 70, 1391–1403. [CrossRef]
69. Wood-Gush, D.G.M.; Kare, M.R. The behaviour of calcium-deficient chickens. Br. Poult. Sci. 1966, 7, 285–290. [CrossRef]
70. Joshua, I.G.; Mueller, W.J. The development of a specific appetite for calcium in growing broiler chicks. Br. Poult. Sci. 1979,
20, 481–490. [CrossRef]
71. Wilkinson, S.J.; Bradbury, E.J.; Bedford, M.R.; Cowieson, A.J. Effect of dietary nonphytate phosphorus and calcium concentration on calcium appetite of broiler chicks. Poult. Sci. 2014, 93, 1695–1703. [CrossRef]
72. Abdollahi, M.; van Dalen, A.; Hendriks, W.; Ravindran, V. Influence of separate feeding of calcium on nutrient digestibility, energy utilisation and performance of young broilers fed pelleted wheat-based diets. Anim. Feed. Sci. Technol. 2015, 205, 122–130.
[CrossRef]
73. Lobaugh, B.; Joshua, I.G.; Mueller, W.J. Regulation of Calcium Appetite in Broiler Chickens. J. Nutr. 1981, 111, 298–306. [CrossRef]
[PubMed]
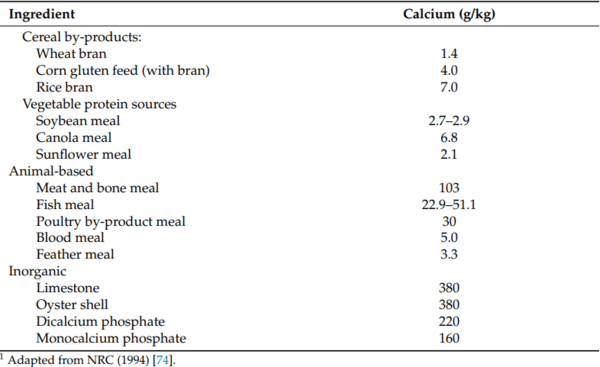
74. National Research Council. Nutrient Requirements of Poultry, 9th ed.; National Academic Press: Washington, DC, USA, 1994.
75. Sloss, L.L. Environments of Limestone Deposition. J. Sediment. Res. 1947, 17, 109–113. [CrossRef]
76. Reid, B.; Weber, C. Calcium Availability and Trace Mineral Composition of Feed Grade Calcium Supplements. Poult. Sci. 1976,
55, 600–605. [CrossRef]
77. Browning, L.; Cowieson, A. The concentration of strontium and other minerals in animal feed ingredients. J. Appl. Anim. Nutr.
2013, 2, e7. [CrossRef]
78. Plumstead, P.W.; Sinclair-Black, M.; Angel, C.R. The benefits of measuring calcium digestibility from raw materials in broilers, meat breeders, and layers. In Proceedings of the 31st Annual Australian Poultry Science Symposium, Sydney, Australia,
16–19 February 2020.
79. Anwar, M.N.; Ravindran, V.; Morel, P.C.H.; Ravindran, G.; Cowieson, A.J. Apparent ileal digestibility of calcium in lime-stone for broiler chickens. Anim. Feed Sci. Technol. 2016, 213, 142–147. [CrossRef]
80. Gilani, S.; Mereu, A.; Li, W.; Plumstead, P.; Angel, R.; Wilks, G.; Dersjant-Li, Y. Global survey of limestone used in poultry diets:
Calcium content, particle size and solubility. J. Appl. Anim. Nutr. 2022, 10, 19–30. [CrossRef]
81. Zhang, B.; Coon, C. The relationship of calcium intake, source, size, solubility in vitro and in vivo, and gizzard limestone retention in laying hens. Poult. Sci. 1997, 76, 1702–1706. [CrossRef]
82. Davin, R.; Kwakernaak, C.; Dersjant-Li, Y. Effect of two commercial limestone sources with different solubility on the efficacy of two phytases in 0–21 d old broilers. J. Appl. Anim. Nutr. 2020, 8, 61–73. [CrossRef]
83. Stillmak, S.J.; Sunde, M.L. The Use of High Magnesium Limestone in the Diet of the Laying Hen: 1. Egg Production. Poult. Sci.
1971, 50, 553–564. [CrossRef]
84. Cheng, T.K.; Coon, C.N. Effect of Calcium Source, Particle Size, Limestone Solubility In Vitro, and Calcium Intake Level on Layer
Bone Status and Performance. Poult. Sci. 1990, 69, 2214–2219. [CrossRef]
85. Rao, K.S.; Roland, D.A.; Adams, J.L.; Durboraw, W.M. Improved Limestone Retention in the Gizzard of Commercial Leghorn
Hens. J. Appl. Poult. Res. 1992, 1, 6–10. [CrossRef]
86. Kim, S.-W.; Li, W.; Angel, R.; Plumstead, P. Modification of a limestone solubility method and potential to correlate with in vivo limestone calcium digestibility. Poult. Sci. 2019, 98, 6837–6848. [CrossRef]
87. Manangi, M.K.; Coon, C.N. The effect of calcium carbonate particle size and solubility on the utilisation of phosphorus from phytase for broilers. Int. J. Poult. Sci. 2007, 6, 85–90.
88. Kim, S.-W.; Li, W.; Angel, R.; Proszkowiec-Weglarz, M. Effects of limestone particle size and dietary Ca concentration on apparent
P and Ca digestibility in the presence or absence of phytase. Poult. Sci. 2018, 97, 4306–4314. [CrossRef]
89. Li, W.; Angel, R.; Plumstead, P.; Enting, H. Effects of limestone particle size, phytate, calcium source, and phytase on standardized ileal calcium and phosphorus digestibility in broilers. Poult. Sci. 2021, 100, 900–909. [CrossRef] [PubMed]
90. Cheng, T.K.; Coon, C.N. Comparison of Various In Vitro Methods for the Determination of Limestone Solubility. Poult. Sci. 1990,
69, 2204–2208. [CrossRef]
91. de Witt, F.H.; van der Merwe, H.J.; Hayes, J.P.; Fair, M.D. Influence of particle size distribution on in vivo and in vitro limestone solubility. S. Afr. J. Anim. Sci. 2006, 36, 95–98.
92. Augspurger, N.R.; Baker, D.H. Phytase improves dietary calcium utilization in chicks, and oyster shell, carbonate, citrate, and citrate-malate forms of calcium are equally bioavailable. Nutr. Res. 2004, 24, 293–301. [CrossRef]
93. Scott, M.; Hull, S.; Mullenhoff, P. The Calcium Requirements of Laying Hens and Effects of Dietary Oyster Shell Upon Egg Shell
Quality. Poult. Sci. J. 1971, 50, 1055–1063. [CrossRef]
94. Roland, D.A., Sr. Eggshell quality IV: Oyster shell versus limestone and the importance of particle size or solubility of calcium source. World’s Poult. Sci. J. 1986, 42, 166–171. [CrossRef]
95. Viljoen, H. Utilisation of feed phosphates: Fact or confusion? AFMA Matrix 2001, 10, 24–27.
96. Sullivan, T.; Douglas, J.; Lapjatupon, W.; Struwe, F.; Gonzalez, N. Biological Value of Bone-Precipitated Dicalcium Phosphate in
Turkey Starter Diets. Poult. Sci. 1994, 73, 122–128. [CrossRef] [PubMed]
97. Carrodeguas, R.G.; De Aza, S. α-Tricalcium phosphate: Synthesis, properties and biomedical applications. Acta Biomater. 2011,
7, 3536–3546. [CrossRef] [PubMed]
98. Ravindran, V. Poultry Feed Availability and Nutrition in Developing Countries: Feed Supplements and Additives. Poultry
Development Review, Food and Agriculture Organisation of the United Nations, Rome. 2013. Available online: http://www.fao. org/ag/againfo/themes/en/poultry/AP_nutrition.html (accessed on 5 July 2018).
99. Scientific Steering Committee. Updated Opinion and Report on the Safety of Dicalcium Phosphate (DCP) and Tricalcium
Phosphate (TCP) from Bovine Bones, Used as an Animal Feed Additive or as Fertiliser. European Commission, Health and
Consumer Protection Directorate-General. 2003. Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/scicom_ssc_out322_en.pdf (accessed on 6 June 2021).
100. Hamdi, M.; Sola’-Oriol, D.; Davin, R.; Perez, J.F. Calcium sources and their interaction with the different levels of non-phytate phosphorous affect performance and bone mineralisation in broiler chickens. Poult. Sci. 2015, 94, 2136–2143. [CrossRef]
101. Rao, S.K.; Roland Sr, D.A.; Gordon, R.W. A method to determine and factors that influence in vivo solubilisation of phosphates in commercial leghorn hens. Poult. Sci. 1995, 74, 1644–1649. [CrossRef]
102. Kwon, W.B.; Kim, B.G. Standardised total tract digestibility of phosphorus in various inorganic phosphates fed to growing pigs.
Anim. Sci. J. 2017, 88, 918–924. [CrossRef]
103. Jacob, K.D.; Hill, W.L.; Marshall, H.L.; Reynolds, D.S. The Composition and Distribution of Phosphate Rock with Special Reference to the
United States; Technical Bulletin No. 364; United States Department of Agriculture: Washington, DC, USA, 1933.
104. Whitney, W.T.; Hollingsworth, C.A. Production of deflourinated phosphate rock. Calcining without fusion in rotary kilns. Ind.
Eng. Chem. 1949, 41, 1325–1327. [CrossRef]
105. Hendry, M.H.; Pesti, G.M. An investigation of calcium citrate-malate as a calcium source for young broiler chicks. Poult. Sci. 2002,
81, 1149–1155. [CrossRef]
106. Fernandes, I.J.; Lima, F.R.; Mendonça, C.X.; Mabe, I.; Albuquerque, R.; Leal, P.M. Relative bioavailability of phosphorus in feed and agricultural phosphates for poultry. Poult. Sci. 1999, 78, 1729–1736. [CrossRef] [PubMed]
107. Waldroup, P.W.; Adams, M.H. Evaluation of the Phosphorus Provided by Animal Proteins in the Diet of Broiler Chickens. J. Appl.
Poult. Res. 1994, 3, 209–218. [CrossRef]
108. Sulabo, R.C.; Stein, H.H. Digestibility of phosphorus and calcium in meat and bone meal fed to growing pigs. J. Anim. Sci. 2013,
91, 1285–1294. [CrossRef]
109. Anwar, M.; Ravindran, V.; Morel, P.; Ravindran, G.; Cowieson, A. Measurement of true ileal calcium digestibility in meat and bone meal for broiler chickens. Anim. Feed. Sci. Technol. 2015, 206, 100–107. [CrossRef]
110. National Research Council. Nutrient Requirements of Poultry, 8th ed.; National Academy Press: Washington, DC, USA, 1984.
111. Angel, R. Calcium to phosphorous ratios in broilers. In Proceedings of the 24th Australian Poultry Science Symposium, Sydney,
Australia, 17–20 February 2013.
112. Mutucumarana, R.K.; Ravindran, V.; Ravindran, G.; Cowieson, A.J. Measurement of true ileal digestibility and total tract retention of phosphorous in corn and canola meal for broiler chickens. Poult. Sci. 2014, 93, 412–419. [CrossRef] [PubMed]
113. Ravindran, V.; Bryden, W.L.; Kornegay, E.T. Phytates: Occurrence, bioavailability and implications in poultry nutrition. Poult.
Avian Biol. Rev. 1995, 6, 125–143.
114. Li, X.; Zhang, D.; Bryden, W.L. Calcium and phosphorus metabolism and nutrition of poultry: Are current diets formulated in excess? Anim. Prod. Sci. 2017, 57, 2304–2310. [CrossRef]
115. Applegate, T.; Angel, R.; Classen, H. Effect of dietary calcium, 25-hydroxycholecalciferol, or bird strain on small intestinal phytase activity in broiler chickens. Poult. Sci. 2003, 82, 1140–1148. [CrossRef]
116. Peeler, H.T. Biological Availability of Nutrients in Feeds: Availability of Major Mineral Ions. J. Anim. Sci. 1972, 35, 695–712.
[CrossRef]
117. Kiarie, E.; Nyachoti, C.M. Bioavailability of calcium and phosphorus in feedstuffs for farm animals. In Phosphorus and Calcium
Utilisation and Requirements in Farm Animals; Vitti, D.M.S.S., Kebreab, E., Eds.; CABI Publishing: Wallingford, UK, 2010; pp. 76–93.
118. Dilworth, B.C.; Day, E.J.; Hill, J.E. Availability of Calcium in Feed Grade Phosphates to the Chick. Poult. Sci. 1964, 43, 1132–1134.
[CrossRef]
119. Suttle, N.F. Mineral Nutrition of Livestock, 4th ed.; CABI Publishing: Wallingford, UK, 2010; p. 83.
120. Taylor, T.C. The availability of the calcium and phosphorous of plant materials for animals. Proc. Nutr. Soc. 1965, 24, 105–112.
[CrossRef]
121. Nelson, T.S.; McGillivray, J.J.; Shieh, T.R.; Wodzinski, R.J.; Ware, J.H. Effect of Phytate on the Calcium Requirement of Chicks.
Poult. Sci. 1968, 47, 1985–1989. [CrossRef]
122. Rodehutscord, M. Determination of phosphorus availability in poultry. Working group No. 2: Nutrition of the European
Federation of Branches of WPSA. World’s Poult. Sci. J. 2013, 69, 687–698.
123. Rao, K.S.; Roland Sr, D.A. In vivo limestone solubilisation in commercial leghorns: Role of dietary calcium level, limestone particle size, in vitro limestone solubility rate, and the calcium status of the hen. Poult. Sci. 1990, 69, 2170–2176. [CrossRef]
[PubMed]
124. Ravindran, V.; Hew, L.; Ravindran, G.; Bryden, W. A comparison of ileal digesta and excreta analysis for the determination of amino acid digestibility in food ingredients for poultry. Br. Poult. Sci. 1999, 40, 266–274. [CrossRef] [PubMed]
125. Ravindran, V. Additivity of apparent and true ileal amino acid digestibility coefficients in complete broiler diets. In Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007.
126. Angkanaporn, K.; Ravindran, V.; Bryden, W. Additivity of Apparent and True Ileal Amino Acid Digestibilities in Soybean Meal,
Sunflower Meal, and Meat and Bone Meal for Broilers. Poult. Sci. 1996, 75, 1098–1103. [CrossRef] [PubMed]
127. Cilliers, S.; Hayes, J.; Sales, J.; Chwalibog, A.; Du Preez, J. The additivity of TMEn values of various ingredients in a complete diet for ostriches and adult roosters. Anim. Feed. Sci. Technol. 1998, 71, 369–373. [CrossRef]
128. Hong, D.; Ragland, D.; Adeola, O. Additivity and associative effects of metabolisable energy and amino acid digestibility in barley and canola meal for White Pekin ducks. Poult. Sci. 2001, 80, 1600–1606. [CrossRef]
129. Hong, D.; Ragland, D.; Adeola, O. Additivity and associative effects of metabolisable energy and amino acid digestibility of corn, soybean meal, and wheat red dog for White Pekin ducks. J. Anim. Sci. 2002, 80, 3222–3229. [CrossRef]
130. Kong, C.; Adeola, O. Ileal endogenous amino acid flow response to nitrogen-free diets with differing ratios of corn starch to dextrose in broiler chickens. Poult. Sci. 2013, 92, 1276–1282. [CrossRef] [PubMed]
131. Zhang, F.; Adeola, O. True ileal digestibility of calcium in limestone and dicalcium phosphate are additive in diets of broiler chickens. Poult. Sci. 2018, 97, 4290–4296. [CrossRef] [PubMed]
132. Walk, C.L.; Romero, L.F.; Cowieson, A.J. Towards a digestible calcium system for broiler chicken nutrition: A review and recommendations for the future. Anim. Feed. Sci. Technol. 2021, 276, 114930. [CrossRef]
133. Angel, R.; Proszkowiec-Weglarza, M.; Jimenez-Moreno, E.; Kim, S.; Plumstead, P. Impact of the time and dietary calcium and phosphorous deficiencies on their digestibilities in single ingredients. In Proceedings of the European Symposium of Poultry
Nutrition, Potsdam, Germany, 26–29 August 2013.
134. David, L.S.; Abdollahi, M.R.; Bedford, M.R.; Ravindran, V. True ileal calcium digestibility in soybean meal and canola meal, and true ileal phosphorous digestibility in maize-soybean meal and maize-canola meal diets, without and with microbial phytase, for broiler growers and finishers. Br. Poult. Sci. 2020, 62, 293–303. [CrossRef]
135. Stein, H.H.; Sève, B.; Fuller, M.F.; Moughan, P.J.; De Lange, C.F.M. Invited review: Amino acid bioavailability and digestibility in pig feed ingredients: Terminology and application. J. Anim. Sci. 2007, 85, 172–180. [CrossRef] [PubMed]
136. Imari, Z.K.; Hassanabadi, A.; Moghaddam, H.N. Response of broiler chickens to calcium and phosphorus restriction: Effects on growth performance, carcase traits, tibia characteristics and total tract retention of nutrients. Ital. J. Anim. Sci. 2020, 19, 929–939.
[CrossRef]
137. Viveros, A.; Brenes, A.; Arija, I.; Centeno, C. Effects of microbial phytase supplementation on mineral utilization and serum enzyme activities in broiler chicks fed different levels of phosphorous. Poult. Sci. 2002, 81, 1172–1183. [CrossRef]
138. van Krimpen, M.M.; Van Diepen, J.T.M.; Van Wikselaar, P.G.; Bikker, P.; Jongbloed, A.W. Effects of Available Phosphorus (aP),
Calcium/aP Ratio, and Growth Rate on P Deposition, P Digestibility, Performance and Leg Quality in Broilers; Livestock Research Report
No. 670; Wageningen UR Livestock Research: Wageningen, The Netherlands, 2013.
139. Amerah, A.M.; Plumstead, P.W.; Barnard, L.P.; Kumar, A. Effect of calcium level and phytase addition on ileal phytate degradation and amino acid digestibility of broilers fed corn-based diets. Poult. Sci. 2014, 93, 906–915. [CrossRef]
140. Majeed, S.; Qudsieh, R.; Edens, F.; Brake, J. Limestone particle size, calcium and phosphorus levels, and phytase effects on live performance and nutrients digestibility of broilers. Poult. Sci. 2020, 99, 1502–1514. [CrossRef] [PubMed]
141. Ravindran, V.; Morel, P.C.; Partridge, G.G.; Hruby, M.; Sands, J.S. Influence of an Escherichia coli-derived phytase on nutrient utilisation in broiler starters fed diets containing varying concentrations of phytic acid. Poult. Sci. 2006, 85, 82–89. [CrossRef]
142. Walk, C.L.; Addo-Chidie, E.K.; Bedford, M.R.; Adeola, O. Evaluation of a highly soluble calcium source and phytase in the diets of broiler chickens. Poult. Sci. 2012, 91, 2255–2263. [CrossRef] [PubMed]
143. Bradbury, E.J.; Wilkinson, S.J.; Cronin, G.M.; Walk, C.L.; Cowieson, A.J. Effects of phytase, calcium source, calcium concentration and particle size on broiler performance, nutrient digestibility and skeletal integrity. Anim. Prod. Sci. 2018, 58, 271–283. [CrossRef]
144. Neto, R.M.; Cozannet, P.; Preynat, A. Phosphorus and calcium digestibility is improved with the addition of a carbohydrase complex in broiler chickens fed wheat-soybean-based diets. In Proceedings of the 20th European Symposium on Poultry Nutrition,
Prague, Czech Republic, 24–27 August 2015.
145. Moss, A.M.; Chrystal, P.V.; Dersjant-Li, Y.; Selle, P.H.; Liu, S.Y. Responses in digestibilities of macro-minerals, trace minerals and amino acids generated by exogenous phytase and xylanase in canola meal diets offered to broiler chickens. Anim. Feed Sci.
Technol. 2018, 240, 22–30. [CrossRef]
146. Attar, A.; Kermanshahi, H.; Golian, A. Effects of conditioning time and sodium bentonite on pellet quality, growth performance, intestinal morphology and nutrient retention in finisher broilers. Br. Poult. Sci. 2017, 59, 190–197. [CrossRef] [PubMed]
147. Mahmood, S.; Ali, H.; Ahmad, F.; Iqbal, Z. Estimation of tannins in different sorghum varieties and their effects on nutrient digestibility and absorption of some minerals in caged white leghorn layers. Int. J. Agric. Biol. 2014, 16, 217–221.
148. Fonolla, J.; Prieto, C.; Sanz, R. Influence of age on the nutrient utilization of diets for broilers. Anim. Feed. Sci. Technol. 1981,
6, 405–411. [CrossRef]
149. Shastak, Y.; Witzig, M.; Hartung, K.; Rodehutscord, M. Comparison of retention and prececal digestibility measurements in evaluating mineral phosphorus sources in broilers. Poult. Sci. 2012, 91, 2201–2209. [CrossRef]
150. Li, W.; Angel, R.; Kim, S.-W.; Jiménez-Moreno, E.; Proszkowiec-Weglarz, M.; Plumstead, P. Impacts of age and calcium on Phytase efficacy in broiler chickens. Anim. Feed. Sci. Technol. 2018, 238, 9–17. [CrossRef]
151. Mtei, A.W.; Abdollahi, M.R.; Schreurs, N.M.; Ravindran, V. Impact of corn particle size on nutrient digestibility varies depending on bird type. Poult. Sci. 2019, 98, 5504–5513. [CrossRef]
152. Ingelmann, C.-J.; Witzig, M.; Möhring, J.; Schollenberger, M.; Kühn, I.; Rodehutscord, M. Phytate degradation and phosphorus digestibility in broilers and turkeys fed different corn sources with or without added phytase. Poult. Sci. 2019, 98, 912–922.
[CrossRef]
153. Baker, S.; Herrman, T. Evaluating Particle Size, MF-2051. Feed Manufacturing; Department of Grain Science and Industry, Kansas
State University: Manhattan, KS, USA, 2002.
154. Zhang, B.; Coon, C.N. Improved In Vitro Methods for Determining Limestone and Oyster Shell Solubility. J. Appl. Poult. Res.
1997, 6, 94–99. [CrossRef]
155. Morgan, N.K.; Walk, C.L.; Bedford, M.R.; Burton, E.J. Contribution of intestinal- and cereal-derived phytase activity on phytate degradation in young broilers. Poult. Sci. 2015, 94, 1577–1583. [CrossRef] [PubMed]
156. Cosgrove, D.J.; Irving, G.C.J. Inositol Phosphates: Their Chemistry, Biochemistry and Physiology; Elsevier Scientific Publishing:
Amsterdam, The Netherlands, 1980.
157. Walters, H.; Coelho, M.; Coufal, C.; Lee, J. Effects of Increasing Phytase Inclusion Levels on Broiler Performance, Nutrient
Digestibility, and Bone Mineralization in Low-Phosphorus Diets. J. Appl. Poult. Res. 2019, 28, 1210–1225. [CrossRef]
158. Qian, H.; Kornegay, E.T.; Conner, D.E., Jr. Adverse effects of wide calcium:phosphorus ratios on supplemental phytase efficacy for weanling pigs fed two dietary phosphorus levels. J. Anim. Sci. 1996, 74, 1288–1297. [CrossRef] [PubMed]
159. Qian, H.; Kornegay, E.T.; Denbow, D.M. Phosphorus equivalence of microbial phytase in Turkey diets as influenced by calcium to phosphorous ratios and phosphorous levels. J. Anim. Sci. 1996, 75, 69–81.
160. Angel, R.; Tamim, N.M.; Applegate, T.J.; Dhandu, A.S.; Ellestad, L.E. Phytic Acid Chemistry: Influence on Phytin-Phosphorus
Availability and Phytase Efficacy. J. Appl. Poult. Res. 2002, 11, 471–480. [CrossRef]
161. Paiva, D.M.; Walk, C.L.; McElroy, A.P. Influence of dietary calcium level, calcium source, and phytase on bird performance and mineral digestibility during a natural necrotic enteritis episode. Poult. Sci. 2013, 92, 3125–3133. [CrossRef] [PubMed]
162. Akter, M.; Graham, H.; Iji, P.A. Response of broiler chickens to different levels of calcium, non-phytate phosphorus and phytase.
Br. Poult. Sci. 2016, 57, 799–809. [CrossRef]
163. Walk, C.L.; Bedford, M.R.; McElroy, A.P. Influence of limestone and phytase on broiler performance, gastrointestinal pH, and apparent ileal nutrient digestibility. Poult. Sci. 2012, 91, 1371–1378. [CrossRef]
164. Cowieson, A.J.; Wilcock, P.; Bedford, M.R. Super-dosing effects of phytase in poultry and other monogastrics. World’s Poult. Sci. J.
2011, 67, 225–236. [CrossRef]
165. Beeson, L.A.; Walk, C.L.; Bedford, M.R.; Olukosi, O.A. Hydrolysis of phytate to its lower esters can influence the growth performance and nutrient utilisation of broilers with regular or super doses of phytase. Poult. Sci. 2017, 96, 2243–2253. [CrossRef]
166. Bedford, M.; Rousseau, X. Recent findings regarding calcium and phytase in poultry nutrition. Anim. Prod. Sci. 2017,
57, 2311–2316. [CrossRef]
167. Kiarie, E.; Woyengo, T.; Nyachoti, C.M. Efficacy of New 6-Phytase from Buttiauxella spp. on Growth Performance and Nutrient
Retention in Broiler Chickens Fed Corn Soybean Meal-based Diets. Asian-Australas. J. Anim. Sci. 2015, 28, 1479–1487. [CrossRef]
168. Farhadi, D.; Karimi, A.; Sadeghi, G.; Rostamzadeh, J.; Bedford, M.R. Effects of a high dose of microbial phytase and myo-inositol supplementation on growth performance, tibia mineralization, nutrient digestibility, litter moisture content, and foot problems in broiler chickens fed phosphorus-deficient diets. Poult. Sci. 2017, 96, 3664–3675. [CrossRef]
169. Sommerfeld, V.; Künzel, S.; Schollenberger, M.; Kühn, I.; Rodehutscord, M. Influence of phytase or myo-inositol supplements on performance and phytate degradation products in the crop, ileum, and blood of broiler chickens. Poult. Sci. 2018, 97, 920–929.
[CrossRef]
170. Walk, C.L.; Olukosi, A.O. Influence of graded concentrations of phytase in high-phytate diets on growth performance, apparent ileal amino acid digestibility, and phytate concentration in broilers from hatch to 28 D post-hatch. Poult. Sci. 2019, 98, 3884–3893.
[CrossRef]
171. Ajuwon, K.M.; Sommerfeld, V.; Paul, V.; Däuber, M.; Schollenberger, M.; Kühn, I.; Adeola, O.; Rodehutscord, M. Phytase dosing affects phytate degradation and Muc2 transporter gene expression in broiler starters. Poult. Sci. 2019, 99, 981–991. [CrossRef]
[PubMed]
172. de Lange, L.; Kwakernaak, C. Ca-Release by Phytase in Broiler Diets; Schothorst Feed Research Report no. 1586; Schothorst:
Amersfoort, The Netherlands, 2017; 14p.
173. Ravindran, V.; Bryden, W.L. Amino acid availability in poultry—In vitro and in vivo measurements. Aust. J. Agric. Res. 1999,
50, 889–908. [CrossRef]
174. Lemme, A.; Ravindran, V.; Bryden, W. Ileal digestibility of amino acids in feed ingredients for broilers. World’s Poult. Sci. J. 2004,
60, 423–438. [CrossRef]
175. Ross. Ross Broiler: Nutrition Specifications; Aviagen: Huntsville, AL, USA, 2022; pp. 3–5.
176. Ross. Ross Broiler: Nutrition Specification; Aviagen: Huntsville, AL, USA, 2019; pp. 4–8.
177. Walk, C.L.; Wang, Z.; Wang, S.; Sorbara, J.O.B.; Zhang, J. Determination of the standardized ileal digestible calcium requirement of male Arbor Acres Plus broilers from day 11 to 24 post-hatch. Poult. Sci. 2022, 101, 101836. [CrossRef]
178. Walk, C.L.; Wang, Z.; Wang, S.; Sorbara, J.O.B.; Zhang, J. Determination of the standardized ileal digestible calcium requirement of male Arbor Acres Plus broilers from day 25 to 42 post-hatch. Poult. Sci. 2022, 101, 102146. [CrossRef]
179. Ravindran, V.; Adeola, O.; Rodehutscord, M.; Kluth, H.; van der Klis, J.; van Eerden, E.; Helmbrecht, A. Determination of ileal digestibility of amino acids in raw materials for broiler chickens—Results of collaborative studies and assay recommendations.
Anim. Feed. Sci. Technol. 2017, 225, 62–72. [CrossRef]
180. Létourneau-Montminy, M.P.; Narcy, A.; Dourmad, J.Y.; Pomar, C.; Crenshaw, T.D. Modeling the metabolic fate of dietary phosphorus and calcium and the dynamics of body ash content in growing pigs. J. Anim. Sci. 2015, 93, 1200. [CrossRef] [PubMed]
181. Lautrou, M.; Pomar, C.; Dourmad, J.Y.; Narcy, A.; Schmidely, P.; Letourneau-Montminy, M.P. Phosphorus and calcium requirements of bone mineralization of growing pigs predicted by mechanistic modelling. Animal 2020, 14, 313–322. [CrossRef]
[PubMed]
182. Misiura, M.M.; Filipe, J.A.N.; Walk, C.L.; Kyriazakis, I. Do nor neglect calcium: A systematic review and met-analysis (metaregression) of its digestibility and utilisation in growing and finishing pigs. Br. J. Nutr. 2018, 119, 1207–1219. [CrossRef]
183. Salisbury, F.; Cowieson, A.; Gous, R. Constraints on the modelling of calcium and phosphorus growth of broilers: A systematic review. World’s Poult. Sci. J. 2021, 77, 775–795. [CrossRef]






Gene Pesti, as evidenced by several research works (Zhand & Adeola, 2017; 2018; Fang et al., 2007 – poultry and pigs) and pointed by Aaron Cowieson, SID Ca/P is more additive in complex mixtures. Some nutrient recommendations have already started using SID P (NRC, 2012) in feed formulations as pointed by Joshua Jendza. Therefore, a concurrent use of SID Ca is necessary because of the interrelationship between Ca and P in the absorption and post-absorptive utilisation of these minerals. Excess P excretion and environmental pollution are reduced by the use of SID P which is a major benefit. In case of pigs, I like to point out that the Ca digestibility measured at both ileal- and total tract- level in pigs was similar as indicated by Zhang et al. (2016). Ca absorption is mostly completed in the jejunum irrespective of dietary Ca concentrations (Mutucumarana et al., 2014), but may be in the upper ileum sometimes (Hurwitz and Bar, 1970)? So, there is no possibility of mineral absorption in the second half of the ileum, I think. I don’t have direct practical experience with studies related to biological responses. But I do have a question regarding ‘highly available mineral source’. Because the availability of limestone was assumed to be 100% earlier. But this is not the case based on the recent digestibility studies.
Alvaro Dubois – Ca digestibility/excretion can be affected by several factors and therefore it is a complex subject, I think. As you said, dietary Ca concentration could be one of the factors. Dietary P, phytate, basal diet type, age & class of birds are some other factors which may influence Ca digestibility. I think, the digestibility/excretion trend of Ca cannot be compared directly with that of P.
.jpg&w=3840&q=75)










.jpg&w=3840&q=75)