Field Efficacy of an Attenuated Infectious Bronchitis Variant 2 Virus Vaccine in Commercial Broiler Chickens
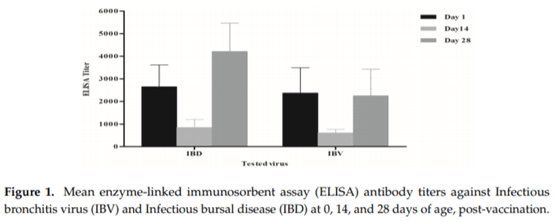
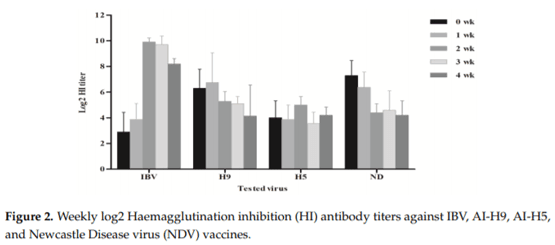
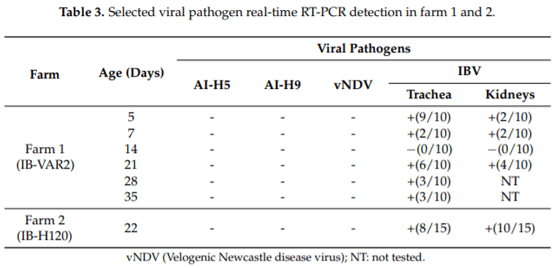
Egyptian poultry suffer from frequent respiratory disease outbreaks associated with Infectious Bronchitis Virus (IBV) variant 2 strains (Egy/VarII). Different vaccination programs using imported vaccines have failed to protect the flocks from field challenge. Recent studies confirmed a successful protection using homologous strains as live attenuated vaccines. In this study, a newly developed live attenuated IB-VAR2 vaccine representing the GI-23 Middle East IBV lineage was evaluated in day-old commercial broilers in an IBV-endemic area. A commercial broiler flock was vaccinated with the IB-VAR2 vaccine at day-old age followed by IB-H120 at day 16. The vaccinated flock was monitored on a weekly basis till the slaughter age. The health status and growth performance were monitored, and selected viral pathogen real-time RT-PCR (rRT-PCR) detection was conducted on a weekly basis. Finally, the flock was compared to a nearby farm with only the classical IB-H120 vaccination program. Results showed that the IB-VAR2 vaccine was tolerable in day-old broiler chicks. The IBV virus rRT-PCR detection was limited to the trachea as compared to its nephropathogenic parent virus. Respiratory disease problems and high mortalities were reported in the IB-H120-only vaccinated flock. An exposure to a wild-type Egy/VarII strain was confirmed in both flocks as indicated by partial IBV S1 gene sequence. Even though the IB-VAR2-vaccinated flock performance was better than the flock that received only IB-H120, the IBV ELISA (enzyme-linked immunosorbent assay) and log2 Haemagglutination inhibition (HI) antibody mean titers remained high (3128 ± 2713 and ≥9 log2, respectively) until the 28th day of age. The current study demonstrates the safety and effectiveness of IB-VAR2 as a live attenuated vaccine in day-old commercial broilers. Also, the combination of IB-VAR2 and classical IBV vaccines confers a broader protective immune response against IBV in endemic areas.
Keywords: IBV; attenuated vaccine; infectious bronchitis virus; Middle East; variant 2

- Cavanagh, D. Coronaviruses in poultry and other birds. Avian Pathol. 2005, 34, 439–448. [CrossRef] [PubMed]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [CrossRef] [PubMed]
- Abdel-Moneim, A.S.; El-Kady, M.F.; Ladman, B.S.; Gelb, J.J. S1 gene sequence analysis of a nephropathogenic strain of avian infectious bronchitis virus in Egypt. Virol. J. 2006, 3, 78. [CrossRef] [PubMed]
- Zanaty, A.; Naguib, M.M.; El-Husseiny, M.H.; Mady, W.; Hagag, N.; Arafa, A.S. The sequence of the full spike S1 glycoprotein of infectious bronchitis virus circulating in Egypt reveals evidence of intra-genotypic recombination. Arch. Virol. 2016, 161, 3583–3587. [CrossRef] [PubMed]
- Susan, S.; El-Hady, M.; Soliman, Y. Isolation and characterization of nephropathogenic strain of infectious bronchitis virus in egypt. J. Am. Sci. 2010, 6, 669–674.
- Zanaty, A.; Arafa, A.S.; Hagag, N.; El-Kady, M. Genotyping and pathotyping of diversified strains of infectious bronchitis viruses circulating in Egypt. World J. Virol. 2016, 5, 125–134. [CrossRef] [PubMed]
- Susan, S.; Ya, S.; Mm, E.-H. Preparation and Evaluation of Master Seed for infectious bronchitis vaccine from local variant isolate. Nat. Sci. 2011, 9, 145–150.
- Abdel-Moneim, A.S.; Afifi, M.A.; El-Kady, M.F. Emergence of a novel genotype of avian infectious bronchitis virus in Egypt. Arch. Virol. 2012, 157, 2453–2457. [CrossRef] [PubMed]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [CrossRef] [PubMed]
- Zhao, Y.; Cheng, J.L.; Liu, X.Y.; Zhao, J.; Hu, Y.X.; Zhang, G.Z. Safety and efficacy of an attenuated Chinese QX-like infectious bronchitis virus strain as a candidate vaccine. Vet. Microbiol. 2015, 180, 49–58. [CrossRef] [PubMed]
- Huo, Y.F.; Huang, Q.H.; Lu, M.; Wu, J.Q.; Lin, S.Q.; Zhu, F.; Zhang, X.M.; Huang, Y.Y.; Yang, S.H.; Xu, C.T. Attenuation mechanism of virulent infectious bronchitis virus strain with QX genotype by continuous passage in chicken embryos. Vaccine 2016, 34, 83–89. [CrossRef] [PubMed]
- Lee, H.J.; Youn, H.N.; Kwon, J.S.; Lee, Y.J.; Kim, J.H.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Characterization of a novel live attenuated infectious bronchitis virus vaccine candidate derived from a Korean nephropathogenic strain. Vaccine 2010, 28, 2887–2894. [CrossRef] [PubMed]
- Gelb, J., Jr.; Weisman, Y.; Ladman, B.S.; Meir, R. S1 gene characteristics and efficacy of vaccination against infectious bronchitis virus field isolates from the United States and Israel (1996 to 2000). Avian Pathol. 2005, 34, 194–203. [CrossRef] [PubMed]
- Awad, F.; Hutton, S.; Forrester, A.; Baylis, M.; Ganapathy, K. Heterologous live infectious bronchitis virus vaccination in day-old commercial broiler chicks: Clinical signs, ciliary health, immune responses and protection against variant infectious bronchitis viruses. Avian Pathol. 2016, 45, 169–177. [CrossRef] [PubMed]
- OIE. Avian Influenza. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2014.
- Slomka, M.J.; Pavlidis, T.; Banks, J.; Shell, W.; Mcnally, A.; Essen, S.; Brown, I.H. Validated H5 Eurasian real-time reverse transcriptase-polymerase chain reaction and its application in H5N1 outbreaks in 2005–2006. Avian Dis. 2007, 51, 373–377. [CrossRef] [PubMed]
- Ben Shabat, M.; Meir, R.; Haddas, R.; Lapin, E.; Shkoda, I.; Raibstein, I.; Perk, S.; Davidson, I. Development of a real-time TaqMan RT-PCR assay for the detection of H9N2 avian influenza viruses. J. Virol. Methods 2010, 168, 72–77. [CrossRef] [PubMed]
- Callison, S.A.; Hilt, D.A.; Boynton, T.O.; Sample, B.F.; Robison, R.; Swayne, D.E.; Jackwood, M.W. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J. Virol Methods 2006, 138, 60–65. [CrossRef] [PubMed]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [CrossRef] [PubMed]
- OIE. Avian Influenza. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2015.
- SAS Institute. SAS/GRAPH 9.1 Reference; SAS Institute: Cary, CA, USA, 2004.
- Hassan, K.E.; Shany, S.A.; Ali, A.; Dahshan, A.H.; El-Sawah, A.A.; El-Kady, M.F. Prevalence of avian respiratory viruses in broiler flocks in Egypt. Poult. Sci. 2016, 95, 1271–1280. [CrossRef] [PubMed]
- Haghighat-Jahromi, M.; Asasi, K.; Nili, H.; Dadras, H.; Shooshtari, A.H. Coinfection of avian influenza virus (H9N2 subtype) with infectious bronchitis live vaccine. Arch. Virol. 2008, 153, 651–655. [CrossRef] [PubMed]
- Tarnagda, Z.; Yougbare, I.; Kam, A.; Tahita, M.C.; Ouedraogo, J.B. Prevalence of infectious bronchitis and Newcastle disease virus among domestic and wild birds in H5N1 outbreaks areas. J. Infect. Dev. Ctries 2011, 5, 565–570. [CrossRef] [PubMed]
- Sultan, H.; Kilany, W.; Ali, A.; Elfeil, W.; Bazid, A.-H.; Sayed, M.E. Protective Efficacy of Different Live Attenuated Infectious Bronchitis Virus (IBV) Vaccination Regimes against Challenge with IBV Variant 2 Circulated in Middle East; Egyptian Veterinary Poultry Association (EVPA): Hurghda, Egypt, 2017; pp. 111–112.
- Bracewell, C.D.; Dawson, P.S.; Allan, W.H. Antibody responses to a live Newcastle disease vaccine when combined with a live infectious bronchitis vaccine. Vet. Rec. 1972, 90, 248–249. [CrossRef] [PubMed]
- Gelb, J., Jr.; Ladman, B.S.; Licata, M.J.; Shapiro, M.H.; Campion, L.R. Evaluating viral interference between infectious bronchitis virus and Newcastle disease virus vaccine strains using quantitative reverse transcription-polymerase chain reaction. Avian Dis. 2007, 51, 924–934. [CrossRef] [PubMed]
- Awad, F.; Forrester, A.; Baylis, M.; Lemiere, S.; Jones, R.; Ganapathy, K. Immune responses and interactions following simultaneous application of live Newcastle disease, infectious bronchitis and avian metapneumovirus vaccines in specific-pathogen-free chicks. Res. Vet. Sci. 2015, 98, 127–133. [CrossRef] [PubMed]
- De Wit, J.J.; Mekkes, D.R.; Kouwenhoven, B.; Verheijden, J.H. Sensitivity and specificity of serological tests for infectious bronchitis virus antibodies in broilers. Avian Pathol. 1997, 26, 105–118. [CrossRef] [PubMed]
- Terregino, C.; Toffan, A.; Beato, M.S.; De Nardi, R.; Vascellari, M.; Meini, A.; Ortali, G.; Mancin, M.; Capua, I. Pathogenicity of a QX strain of infectious bronchitis virus in specific pathogen free and commercial broiler chickens, and evaluation of protection induced by a vaccination programme based on the Ma5 and 4/91 serotypes. Avian Pathol. 2008, 37, 487–493. [CrossRef] [PubMed]
- Bru, T.; Vila, R.; Cabana, M.; Geerligs, H.J. Protection of chickens vaccinated with combinations of commercial live infectious bronchitis vaccines containing Massachusetts, Dutch and QX-like serotypes against challenge with virulent infectious bronchitis viruses 793B and IS/1494/06 Israel variant 2. Avian Pathol. 2017, 46, 52–58. [CrossRef] [PubMed]
- Okino, C.H.; Alessi, A.C.; Montassier Mde, F.; Rosa, A.J.; Wang, X.; Montassier, H.J. Humoral and cell-mediated immune responses to different doses of attenuated vaccine against avian infectious bronchitis virus. Viral Immunol. 2013, 26, 259–267. [CrossRef] [PubMed]
- Awad, F.; Chhabra, R.; Forrester, A.; Chantrey, J.; Baylis, M.; Lemiere, S.; Hussein, H.A.; Ganapathy, K. Experimental infection of IS/885/00-like infectious bronchitis virus in specific pathogen free and commercial broiler chicks. Res. Vet. Sci. 2016, 105, 15–22. [CrossRef] [PubMed]
- Awad, F.; Forrester, A.; Baylis, M.; Lemiere, S.; Ganapathy, K.; Hussien, H.A.; Capua, I. Protection conferred by live infectious bronchitis vaccine viruses against variant Middle East IS/885/00-like and IS/1494/06-like isolates in commercial broiler chicks. Vet. Rec. Open. 2015, 2, e000111. [CrossRef] [PubMed]







1. It is not possible To completely exclude the 793B group of vaccines from the priming of broilers for a day. You need to use for example (at 1day) Ibird793b+Mass. In the future (11-14 days) if you want to strengthen (increase strategy) 4/91+Mas(Ma 5). 2. H120-unfortunately loses its relevance. The reason is the deterioration of epizootics according to IBV





Well, but is it possible mutation in present situation though by applying vaccine starin H 120 strain its available in market but bacterial growth is controlled still mortality moves up waiting yr honourable opinion.

Thank you for this self-explanatory work. In our breeder farm, we observed that Infectious bronchitis is endemic. At some point, we stock commercial broiler in some of our pen houses and sprayed them with IB4/91 at first week. We started experiencing facial swelling, sinusitis with respiratory distress and of course mild mortalities for like 2 batches. We removed the IB4/91 from the vaccination program for the 3rd batch and there was no sign of IB diseases on the farm.
Does it mean that IB4/91 vaccination done for the commercial broiler stocked for the first 2 batches failed or the birds had already been exposed to the field strain before the vaccination?
A very sensitive and makes a diferent about how we should get our vaccines and tge tyoe according to climate and conditions over this ..plus had seen the boosting of tge vaccines but there are also some areas that drs are not taking on a conscience on this by doing as mentioned and have a detemined vaccines for the area.
Challenge are being seen and heads on to attain the resolution by taking time and make sure they come up with the correct vaccine. Thank so much had learnt and i suggest if all dr under vets should address this each and every region so that the selking of and ibv ot hypravia clone or NCD vaccines should be the right ones and proper administered to birds.
Regards,
Daniel.
.jpg&w=3840&q=75)






