
Content sponsored by:
Evonik Animal Nutrition
Functional amino acids to improve the immunity of pigs
Published: August 2, 2018
By: Dr. John K. Htoo, Evonik Nutrition & Care GmbH
Introduction
Despite increased bio-security measures, pigs are often exposed to chronic sub-clinical level of diseases and climatic (heat) stress in commercial farms, causing a lower feed intake and performance. Differences in health or immune status of the pigs is one of the reasons for the large variation in the performance of pigs between commercial farms (Pastorelli et al., 2012). The optimal levels of nutrients including amino acids (AA) must be supplied with the feeds to the animals if the growth, health, and productivity of the animal are to be maintained at optimal level.
The immune system is a defense system to protect the host from invading pathogens. The gastrointestinal tract (GIT; from the stomach to colon) is a key component of the body's systemic immune system. In addition to digestion, absorption of nutrients, the gut serves as an immunological barrier by secreting digestive enzymes and hormones in the enterocytes. The importance of a healthy immunity of animals on the efficiency of nutrient utilization and growth performance is well accepted. When speaking about gut immunity, the focus is mainly on weaned piglets because of their immature digestive and immune systems associated with incidence of gut disorders such as post-weaning diarrhea. Both systemic and gut immune challenges increased after the ban of antimicrobial growth promoters (AGPs) in animal feeds in the Europe Union (EU) in 2006.
There are many feed additives used to enhance the immune status of the animals including probiotics, prebiotics, organic acids, essential oils, phytogenic, enzymes, antibiotics (in some countries) and AA. Some functional AA are involved in immune system functioning (Li et al., 2007). During immune system stimulation (ISS), nutrients are redirected away from growth and towards tissues involved in immune response (Reeds and Jahoor, 2001). This implies that the requirement some key AA for the production of compounds that are involved in the immune response will be increased. The application of the “functional AAs” concept is one of the possible solutions to maintain gut health and promote growth because it can enhance immune status, gut integrity for weaned pigs especially when sanitary and climatic conditions are challenging and the AGPs are not or less used in the diets.
The intent of this review is to discuss the roles of functional AA in immunity of pigs and their applications to improve immunity by more closely meeting AA requirements during immune challenge conditions. For more in depth review of the effects of AA on immunity of animals see Li et. al. (2007) and Wu (2010).
Despite increased bio-security measures, pigs are often exposed to chronic sub-clinical level of diseases and climatic (heat) stress in commercial farms, causing a lower feed intake and performance. Differences in health or immune status of the pigs is one of the reasons for the large variation in the performance of pigs between commercial farms (Pastorelli et al., 2012). The optimal levels of nutrients including amino acids (AA) must be supplied with the feeds to the animals if the growth, health, and productivity of the animal are to be maintained at optimal level.
The immune system is a defense system to protect the host from invading pathogens. The gastrointestinal tract (GIT; from the stomach to colon) is a key component of the body's systemic immune system. In addition to digestion, absorption of nutrients, the gut serves as an immunological barrier by secreting digestive enzymes and hormones in the enterocytes. The importance of a healthy immunity of animals on the efficiency of nutrient utilization and growth performance is well accepted. When speaking about gut immunity, the focus is mainly on weaned piglets because of their immature digestive and immune systems associated with incidence of gut disorders such as post-weaning diarrhea. Both systemic and gut immune challenges increased after the ban of antimicrobial growth promoters (AGPs) in animal feeds in the Europe Union (EU) in 2006.
There are many feed additives used to enhance the immune status of the animals including probiotics, prebiotics, organic acids, essential oils, phytogenic, enzymes, antibiotics (in some countries) and AA. Some functional AA are involved in immune system functioning (Li et al., 2007). During immune system stimulation (ISS), nutrients are redirected away from growth and towards tissues involved in immune response (Reeds and Jahoor, 2001). This implies that the requirement some key AA for the production of compounds that are involved in the immune response will be increased. The application of the “functional AAs” concept is one of the possible solutions to maintain gut health and promote growth because it can enhance immune status, gut integrity for weaned pigs especially when sanitary and climatic conditions are challenging and the AGPs are not or less used in the diets.
The intent of this review is to discuss the roles of functional AA in immunity of pigs and their applications to improve immunity by more closely meeting AA requirements during immune challenge conditions. For more in depth review of the effects of AA on immunity of animals see Li et. al. (2007) and Wu (2010).
Immunity system in pigs
The immune system is an adaptive defense system to protect the host from invading pathogenic microorganisms, i.e. bacteria, viruses, fungi, and parasites (Kuby, 1994). There are two components of immune system: innate (nonspecific) and acquired (specific) immunity. The innate immune system consists of the cellular components, which include monocytes, macrophages, dendritic cells, neutrophils, natural killer cells and extracellular mediators such as cytokines, chemokines, acute phase proteins, complement system, epithelial barriers, and antimicrobial peptides (Parkin and Cohen, 2001; Tan et al., 2013).
Acquired or specific or immunity is induced by exposure to an antigen, naturally or via vaccination and developed more slowly (Kuby, 1994). Acquired immune system can be further divided into humoral and cell mediated immunity. During humoral immunity response, B lymphocytes in the blood secrete immunoglobulins (antibodies) to bind and eliminate the antigens (Parkin and Cohen, 2001). The cell-mediated immunity functions as the interaction of T cells (T lymphocytes) and their associated cytokines to eliminate intracellular antigens. Two major types of T cells are T helper (CD4+ Th) and T cytotoxic (CD8+). The CD4+ Th cells recognize foreign antigen, and assist other cells to eradicate the pathogen. The CD8+ cytotoxic cells are involved in antiviral and antitumour activity (Parkin and Cohen, 2001).
The immune system is an adaptive defense system to protect the host from invading pathogenic microorganisms, i.e. bacteria, viruses, fungi, and parasites (Kuby, 1994). There are two components of immune system: innate (nonspecific) and acquired (specific) immunity. The innate immune system consists of the cellular components, which include monocytes, macrophages, dendritic cells, neutrophils, natural killer cells and extracellular mediators such as cytokines, chemokines, acute phase proteins, complement system, epithelial barriers, and antimicrobial peptides (Parkin and Cohen, 2001; Tan et al., 2013).
Acquired or specific or immunity is induced by exposure to an antigen, naturally or via vaccination and developed more slowly (Kuby, 1994). Acquired immune system can be further divided into humoral and cell mediated immunity. During humoral immunity response, B lymphocytes in the blood secrete immunoglobulins (antibodies) to bind and eliminate the antigens (Parkin and Cohen, 2001). The cell-mediated immunity functions as the interaction of T cells (T lymphocytes) and their associated cytokines to eliminate intracellular antigens. Two major types of T cells are T helper (CD4+ Th) and T cytotoxic (CD8+). The CD4+ Th cells recognize foreign antigen, and assist other cells to eradicate the pathogen. The CD8+ cytotoxic cells are involved in antiviral and antitumour activity (Parkin and Cohen, 2001).
Gut immunity
The GIT is an important component of the body's immune system because it contains greater than 1012 lymphocytes and has a greater concentration of antibodies than any other site in the body (Mayer, 2000). In addition to digestion, absorption and metabolism of nutrients, the intestinal epithelial cells (i.e. enterocytes) secrete digestive enzymes and hormones, and form an immunological barrier between the external environment (e.g. nutrients, microbes and pathogens) and the animal (Stoll and Burrin, 2006). Besides, the GIT also serves as a home for various microbes, which synthesize and degrade nutrients. Thus, the GIT is one of the most metabolically active and complex tissues in the body.
The intestinal epithelial cells also participate in the innate immune system of the GIT by their ability to secrete mucus and antimicrobial peptides (Shao et al., 2001). The mucus layer is mainly made up of mucins which are secreted along the epithelium of the GIT to protect the gut wall from damage and maintain immune function (Li et al., 2007). Besides mucins, the intestinal epithelial cells also secrete antimicrobial peptides (defensins and cathelicidins), and immunoglobulins which all together restrict the interaction of potential pathogens with the gut mucosa (Oswald, 2006). Furthermore, the epithelial cells can produce cytokines including interleukin IL-1, IL-10, IL-15, and IL-18 and chemokines, crucial for the recruitment and activation of immune cells (Stadnyk A.W., 2002).
When speaking about gut immunity, the focus is mainly on weaned piglets because they are less mature and more susceptible to immune challenges compared to mature pigs. In newborn pigs, antibodies in sow’s milk (colostrum) provide the first source of immune protection. The neonatal pig is immunologically immature until about 4 week of age (Blecha, 2001). Currently, piglets are weaned at 3 to 4 weeks of age under commercial conditions. Thus, the first few days after weaning is a stressful time and is often associated with reduced feed intake and growth and increased incidence of diarrhea due to immature digestive and immune systems. Therefore, the development and maintenance of gut immunity is crucial not only for the innate immune defense of the piglets but also for development of the immune status and performance at the later stage of life.
The GIT is an important component of the body's immune system because it contains greater than 1012 lymphocytes and has a greater concentration of antibodies than any other site in the body (Mayer, 2000). In addition to digestion, absorption and metabolism of nutrients, the intestinal epithelial cells (i.e. enterocytes) secrete digestive enzymes and hormones, and form an immunological barrier between the external environment (e.g. nutrients, microbes and pathogens) and the animal (Stoll and Burrin, 2006). Besides, the GIT also serves as a home for various microbes, which synthesize and degrade nutrients. Thus, the GIT is one of the most metabolically active and complex tissues in the body.
The intestinal epithelial cells also participate in the innate immune system of the GIT by their ability to secrete mucus and antimicrobial peptides (Shao et al., 2001). The mucus layer is mainly made up of mucins which are secreted along the epithelium of the GIT to protect the gut wall from damage and maintain immune function (Li et al., 2007). Besides mucins, the intestinal epithelial cells also secrete antimicrobial peptides (defensins and cathelicidins), and immunoglobulins which all together restrict the interaction of potential pathogens with the gut mucosa (Oswald, 2006). Furthermore, the epithelial cells can produce cytokines including interleukin IL-1, IL-10, IL-15, and IL-18 and chemokines, crucial for the recruitment and activation of immune cells (Stadnyk A.W., 2002).
When speaking about gut immunity, the focus is mainly on weaned piglets because they are less mature and more susceptible to immune challenges compared to mature pigs. In newborn pigs, antibodies in sow’s milk (colostrum) provide the first source of immune protection. The neonatal pig is immunologically immature until about 4 week of age (Blecha, 2001). Currently, piglets are weaned at 3 to 4 weeks of age under commercial conditions. Thus, the first few days after weaning is a stressful time and is often associated with reduced feed intake and growth and increased incidence of diarrhea due to immature digestive and immune systems. Therefore, the development and maintenance of gut immunity is crucial not only for the innate immune defense of the piglets but also for development of the immune status and performance at the later stage of life.
Modulation of immune system stimulation on nutrient utilization
Pigs are often exposed to chronic sub-clinical level of disease in commercial farms, leading to a lower nutrient utilization and performance compared with what is potentially possible under good conditions, even if no clinical signs of disease are exhibited (Colditz, 2002; Pastorelli et al., 2012). A comprehensive review of Pastorelli et al. (2012) revealed that ISS especially bacterial infections of the GIT reduced body weight (BW) gain as much as 40% relative to unchallenged pigs, and the reduction in BW gain is partly due to the reduction in feed intake due to ISS. Systemic inflammation mediated by pro-inflammatory cytokines released by the innate (nonspecific) immune system is responsible for decreased feed consumption and body protein accretion (Li et al., 2007). Pro-inflammatory cytokines, particularly IL-1β, IL-6, and tumor necrosis factor (TNF)α, that are produced by stimulated macrophages, modify nutrient utilization during immune challenge, and have a direct effect on liver, brain, muscle and fat tissue (Colditz, 2002).
During ISS, nutrients are redirected away from growth and towards tissues involved in immune response (Reeds and Jahoor, 2001). Under such conditions, the liver becomes the major contributor to whole body protein synthesis, largely because of increased production of a wide range of acute phase proteins (APP), while synthesis and gain of muscle protein are reduced. Major APP in pigs include haptoglobin, fibrinogen, C-reactive protein, serum amyloid A, porcine major acute phase protein and albumin (Chen et al., 2003). These changes in rate and type of protein synthesis will have a direct impact on AA needs qualitatively and quantitatively at the tissue level (Reeds and Jahoor, 2001; Obled, 2002).
Immune cells utilizes AA, but more importantly the liver also need AA for gluconeogenesis and synthesis of APP and glutathione which are essential for immune function (Hunter and Grimble, 1994; Reeds and Jahoor, 2001). The immune status of animals greatly depends upon the availability of AA and other substrates for the synthesis of these proteins and peptides (Li et al., 2007). The AA profile required for immune functioning is different from that of AA required for muscle protein deposition (Reeds and Jahoor, 2001). As a result, AA requirements for immune functioning is not the same as for optimal growth performance. By more closely meeting AA requirements for maintaining a good immunity during ISS (e.g. sub-clinical level of disease), the negative impact of ISS on animal performance will be reduced, thereby also improving production efficiency.
Pigs are often exposed to chronic sub-clinical level of disease in commercial farms, leading to a lower nutrient utilization and performance compared with what is potentially possible under good conditions, even if no clinical signs of disease are exhibited (Colditz, 2002; Pastorelli et al., 2012). A comprehensive review of Pastorelli et al. (2012) revealed that ISS especially bacterial infections of the GIT reduced body weight (BW) gain as much as 40% relative to unchallenged pigs, and the reduction in BW gain is partly due to the reduction in feed intake due to ISS. Systemic inflammation mediated by pro-inflammatory cytokines released by the innate (nonspecific) immune system is responsible for decreased feed consumption and body protein accretion (Li et al., 2007). Pro-inflammatory cytokines, particularly IL-1β, IL-6, and tumor necrosis factor (TNF)α, that are produced by stimulated macrophages, modify nutrient utilization during immune challenge, and have a direct effect on liver, brain, muscle and fat tissue (Colditz, 2002).
During ISS, nutrients are redirected away from growth and towards tissues involved in immune response (Reeds and Jahoor, 2001). Under such conditions, the liver becomes the major contributor to whole body protein synthesis, largely because of increased production of a wide range of acute phase proteins (APP), while synthesis and gain of muscle protein are reduced. Major APP in pigs include haptoglobin, fibrinogen, C-reactive protein, serum amyloid A, porcine major acute phase protein and albumin (Chen et al., 2003). These changes in rate and type of protein synthesis will have a direct impact on AA needs qualitatively and quantitatively at the tissue level (Reeds and Jahoor, 2001; Obled, 2002).
Immune cells utilizes AA, but more importantly the liver also need AA for gluconeogenesis and synthesis of APP and glutathione which are essential for immune function (Hunter and Grimble, 1994; Reeds and Jahoor, 2001). The immune status of animals greatly depends upon the availability of AA and other substrates for the synthesis of these proteins and peptides (Li et al., 2007). The AA profile required for immune functioning is different from that of AA required for muscle protein deposition (Reeds and Jahoor, 2001). As a result, AA requirements for immune functioning is not the same as for optimal growth performance. By more closely meeting AA requirements for maintaining a good immunity during ISS (e.g. sub-clinical level of disease), the negative impact of ISS on animal performance will be reduced, thereby also improving production efficiency.
Amino acids with immune function
In addition to primary role of serving as a building block for protein synthesis, AA are also involved in various important metabolic pathways in the body. Amino acids that regulates key metabolic pathways to improve health, survival, growth, development, lactation, and reproduction of organisms are defined as functional AA (Wu, 2009). Indeed, some function AA modulate immune system functioning at various levels, i.e. in the GIT, thymus, spleen, lymph nodes and immune cells of the circulating blood (Cunningham-Rundles, 2002). In general, AA regulate the immune system by: 1) enhancing the immune status to prevent infections, and 2) reducing or eliminating established infections such as inflammation and autoimmunity (Yoneda et al., 2009). The most important functional AA include sulfur amino acids, i.e. methionine and cysteine (SAA; Met + Cys), tryptophan (Trp), threonine (Thr), glutamine (Gln), arginine (Arg) and glycine (Gly; Li et al., 2007). In the following, the most important functions beyond growth and beneficial effects of dietary supplementation with these functional AA mainly in pigs are summarized.
In addition to primary role of serving as a building block for protein synthesis, AA are also involved in various important metabolic pathways in the body. Amino acids that regulates key metabolic pathways to improve health, survival, growth, development, lactation, and reproduction of organisms are defined as functional AA (Wu, 2009). Indeed, some function AA modulate immune system functioning at various levels, i.e. in the GIT, thymus, spleen, lymph nodes and immune cells of the circulating blood (Cunningham-Rundles, 2002). In general, AA regulate the immune system by: 1) enhancing the immune status to prevent infections, and 2) reducing or eliminating established infections such as inflammation and autoimmunity (Yoneda et al., 2009). The most important functional AA include sulfur amino acids, i.e. methionine and cysteine (SAA; Met + Cys), tryptophan (Trp), threonine (Thr), glutamine (Gln), arginine (Arg) and glycine (Gly; Li et al., 2007). In the following, the most important functions beyond growth and beneficial effects of dietary supplementation with these functional AA mainly in pigs are summarized.
Sulfur-containing amino acids:
Methionine is a nutritionally essential AA while Cys is a nonessential AA because it can be synthesized from Met but cannot be transformed into Met. Methionine serves as a methyl donor for important processes such as DNA methylation and polyamine synthesis (Grimble, 2002), and these roles become increasingly important during immune challenge to enhance immune cells proliferation (Dwyer, 1979). Cysteine is the rate-limiting substrate for the synthesis of glutathione (GSH) which is the major intracellular antioxidant, consisting of a tripeptide of glutamate (Glu), Cys and Gly (Wu et al., 2004). The GSH is involved in immune function as it is needed for the activation of T-lymphocytes and leukocytes, and for the production of cytokines (Lu, 2009; Wu et al., 2004).
Cysteine is also needed to produce taurine which is important for immune functioning by acting as a cell membrane stabilizer and antioxidant (Grimble, 2002), and it is particularly abundant in leucocytes (Roth, 2007). For immune function, the utilization of Cys for the production of compounds that are involved in the immune response, such as GSH, taurine and APP is increased (Grimble, 2002). This implies that the need for Met to supply Cys also increases during immune challenge situations.
Immune system stimulation by lipopolysaccharides (LPS) injection per se does not affect the ileal digestibility of AA and energy but it reduced the ratio of whole-body nitrogen (N) and sulfur (S)balance indicating that SAA are preferentially preserved in favor of non-protein compounds such as glutathione to enhance immune status (Rakhshandeh et al., 2010). Rakhshandeh et al. (2014) found that ISS reduced protein deposition (Pd) and decreased daily SAA requirement (Fig. 1). However, ISS increased he maintenance SAA requirements, e.g. to achieve a constant PD of 50 g/d, unchallenged (ISS-) pigs need 1.63 g SID SAA intake while immune challenged (ISS+) pigs need 1.87 g SID SAA intake (i.e. 15% increase).
Cysteine is also needed to produce taurine which is important for immune functioning by acting as a cell membrane stabilizer and antioxidant (Grimble, 2002), and it is particularly abundant in leucocytes (Roth, 2007). For immune function, the utilization of Cys for the production of compounds that are involved in the immune response, such as GSH, taurine and APP is increased (Grimble, 2002). This implies that the need for Met to supply Cys also increases during immune challenge situations.
Immune system stimulation by lipopolysaccharides (LPS) injection per se does not affect the ileal digestibility of AA and energy but it reduced the ratio of whole-body nitrogen (N) and sulfur (S)balance indicating that SAA are preferentially preserved in favor of non-protein compounds such as glutathione to enhance immune status (Rakhshandeh et al., 2010). Rakhshandeh et al. (2014) found that ISS reduced protein deposition (Pd) and decreased daily SAA requirement (Fig. 1). However, ISS increased he maintenance SAA requirements, e.g. to achieve a constant PD of 50 g/d, unchallenged (ISS-) pigs need 1.63 g SID SAA intake while immune challenged (ISS+) pigs need 1.87 g SID SAA intake (i.e. 15% increase).
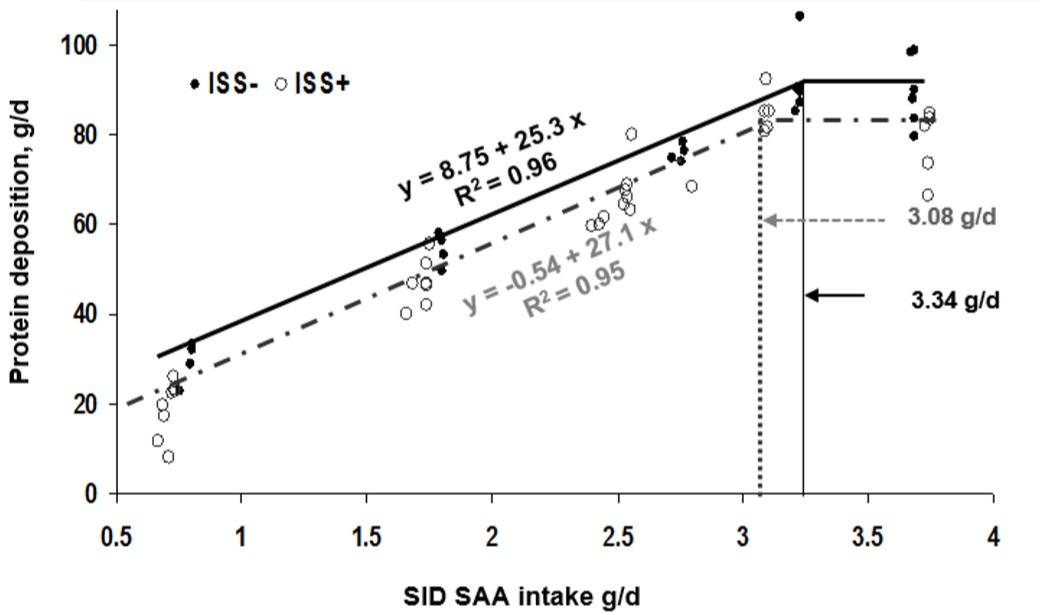
Figure 1: Impact of immune system stimulation (ISS) and standardized ileal digestible (SID) SAA intake on whole-body protein deposition in growing pigs (Rakhshandeh et al., 2014)
The concentration and fractional synthesis rate of plasma albumin decreased when LPS immune challenged pigs were fed a low Met+Cys diet compared with pigs fed a diet adequate in Met+Cys content (Litvak et al., 2013a). Ina N-balance study with growing pigs, Litvak et al. (2013b) showed that immune system stimulation (LPS injection) reduces protein deposition rate but the optimal dietary Met to Met+Cys ratio for maximum body protein deposition is increased from 57 to 59% (Fig. 2). This suggests that the increased need of dietary Met+Cys for production of GSH during sub-optimal immunity should better be provided via Met supply because cysteine (Cys) is extremely unstable and rapidly oxidizes tocystine (i.e. a dimerized form preserved in extracellular fluids), which releases free radicals resulting the toxic properties (Grimble, 2002).
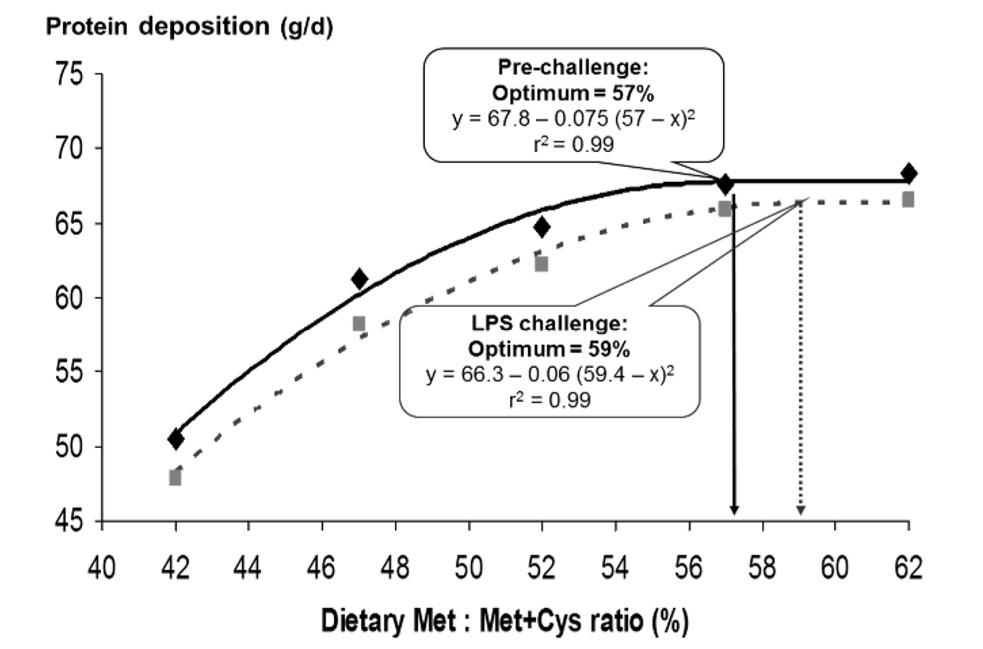
Figure 2: Protein Deposition (g/d) at Varying Levels of Met:Met+Cys without and with LPS challenge (Litvak et al., 2013b)
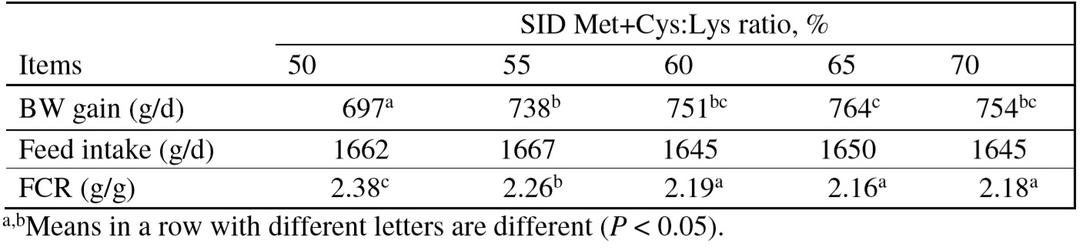
The dietary SID Met+Cys:Lys ratio to maximize body protein deposition increases from 55 to 75% when in growing pigs are immune challenged with LPS (Kim et al., 2012). Under commercial conditions, i.e. prone to pathogens and without in-feed antibiotics, the performance of 25-50 kg pigs were maximized at a dietary SID Met+Cys:Lys ratio of 65% (Zhang et al., 2015; Table 1). This agrees with Capozzalo et al. (2014) who reported that BW gain and feed conversion ratio (FCR) of 8-20 kg pigs infected with E. coli and fed antibiotic-free diets optimized at the SID Met+Cys:Lys ratio of 62.2%. These results indicate that the need of Met+Cys, including Met requirement for converting to Cys, is increased during immune challenge.
Table 1: Effect of dietary SID Met+Cys:Lys ratio on the performance of 25-50 kg growing pigs (Zhang et al., 2015)
Tryptophan:
In addition to being involved in protein synthesis and serotonin regulation, Trp is also is needed for modulation of the immune function through kynurenine pathway initiated by two enzymes. The enzyme tryptophan-2, 3-dioxygenase (TDO) in the liver regulates homeostatic plasma Trp concentration and another enzyme, indoleamine-2, 3-dioxygenase (IDO), which presents in various body tissues including intestine, stomach, lungs, brain and macrophages, is induced by inflammatory cytokine IFN-γ during immune system stimulation (Botting, 1995). More than 95% of dietary Trp, not utilized for protein synthesis, is metabolized through the kynurenine pathway forming various products including kynurenic acid and niacin.
Studies have shown that lung inflammation in pigs reduced plasma Trp levels (Melchior et al., 2004; Fig. 3) and increased IDO activity in lungs and associated lymph nodes (Le Floc’h et al., 2008) than pair-fed healthy piglets. Furthermore, they observed that piglets fed a low Trp diet had a higher plasma concentration of a major acute phase protein haptoglobin (which is relatively high in Trp content) compared with pigs fed a Trp-adequate diet. These results suggest that the use of Trp for the synthesis of kynurenine is increasedduringimmune challenge. Furthermore, Trp can be used to synthesize a neurohormone melatonin, which may act as free radical scavengers and have antioxidant properties (Le Floc'h and Seve, 2007).
In addition to being involved in protein synthesis and serotonin regulation, Trp is also is needed for modulation of the immune function through kynurenine pathway initiated by two enzymes. The enzyme tryptophan-2, 3-dioxygenase (TDO) in the liver regulates homeostatic plasma Trp concentration and another enzyme, indoleamine-2, 3-dioxygenase (IDO), which presents in various body tissues including intestine, stomach, lungs, brain and macrophages, is induced by inflammatory cytokine IFN-γ during immune system stimulation (Botting, 1995). More than 95% of dietary Trp, not utilized for protein synthesis, is metabolized through the kynurenine pathway forming various products including kynurenic acid and niacin.
Studies have shown that lung inflammation in pigs reduced plasma Trp levels (Melchior et al., 2004; Fig. 3) and increased IDO activity in lungs and associated lymph nodes (Le Floc’h et al., 2008) than pair-fed healthy piglets. Furthermore, they observed that piglets fed a low Trp diet had a higher plasma concentration of a major acute phase protein haptoglobin (which is relatively high in Trp content) compared with pigs fed a Trp-adequate diet. These results suggest that the use of Trp for the synthesis of kynurenine is increasedduringimmune challenge. Furthermore, Trp can be used to synthesize a neurohormone melatonin, which may act as free radical scavengers and have antioxidant properties (Le Floc'h and Seve, 2007).
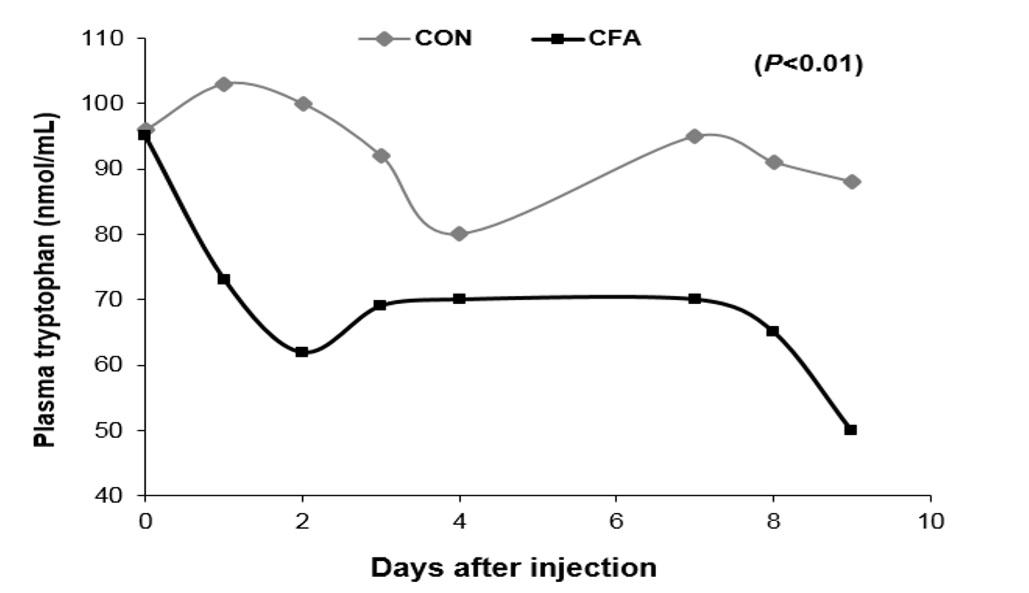
Figure 3: Plasma tryptophan concentration in pigs with lung inflammation (CFA, complete Freund’s adjuvant) and control (CON) healthy paired-fed weaned pigs (Melchior et al., 2004).
Pigs kept under poor sanitary conditions can induce a moderate inflammatory response in pigs. Le Floc’h et al. (2007) reported that the optimal feed intake and BW gain of weaned pigs kept under poor sanitary conditions were achieved at a higher SID Trp:Lysratio (21 vs. 18%) compared with those kept under good sanitary conditions (Fig. 4). The efficiency of Trp utilization for whole body protein deposition of growing pigs reduced during LPS challenge due to increased usage of Trp for immune functions, and a greater dietary Trp (7% increase) is needed to maintain body protein deposition levels similar to those of healthy pigs (Ridder et al., 2012).
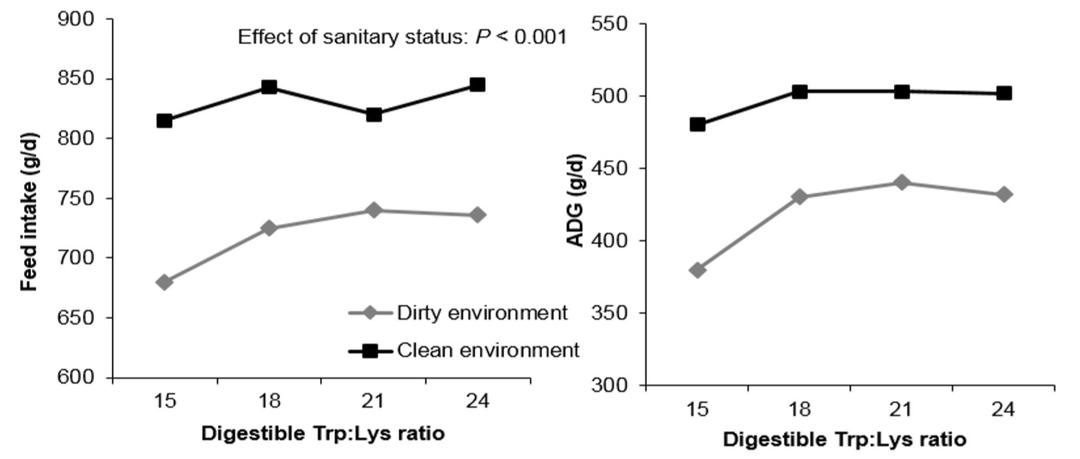
Figure 4: Effects of sanitary status and Trp:Lys ratio on feed intake and weight gain of growing pigs (Le Floc’h et al., 2007)
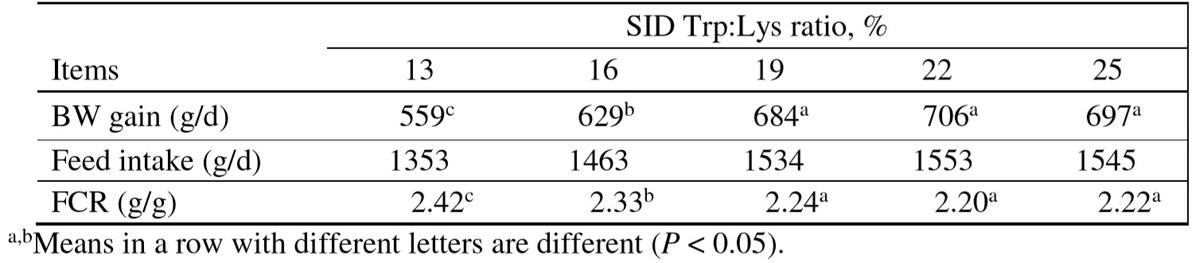
Dietary supplementations with L-Trp at a relatively high level (SID Trp:Lys ratio of 22%) to a AGP-free diet maximized the performance of 25-50 kg pigs raised under commercial conditions (Zhang et al., 2012; Table 2). More recently, Jayaraman et al. (2015) also reported that the mRNA expression of pro-inflammatory cytokine, TNF-α mRNA in ileal tissue of weaned pigs challenged with E. coli K88 linearly decreased with increasing SID Trp: Lys ratio, and an average optimal SID Trp: Lys of 22.6% optimized pig performance. This was in line with Capozzalo et al. (2015) who also found that increasing the dietary SID Trp: Lys ratio to 24% improved FCR and increased plasma levels of Trp and kynurenine of weaned pigs fed antibiotic-free diets and regardless of infection with E. coli. When weaned pigs were housed in a commercial farm with suboptimal immune challenge and fed antibiotic-free diet, suppling a higher dietary SID Trp: Lys ratio to 24% optimized BW gain and FCR (Capozzalo et al., 2013).
Table 2: Effect of dietary SID Trp:Lys ratio on the performance of growing pigs (Zhang et al., 2012)
Threonine:
Threonine plays a key role in immune function through its incorporation into immunoglobulins, which are antibodies produced by plasma cells in response to immune challenge. Threonine is the AA with the greatest amount in immunoglobulins, e.g. about 10.0% in milk immunoglobulins(Bowland, 1966). More than 60% of dietary Thr is used up in the first pass mechanism of the portal drained viscera (PDV; includes small and large intestine, stomach, pancreas, and spleen) in pigs (Stoll et al., 1998). Indeed, Thr requirement of parenterally fed piglets (i.e. bypassing the first pass by the PDV) was reduced by 55% compared with orally fed piglets (Bertolo et al., 1998). The synthesis of mucosal proteins in the GIT includes the secretion of proteins that are secreted into the lumen, including mucins which protect the gut from injury and pathogens. Mucins are particularly rich in Thr, which represents about 30% of the total AA of mucins and 11 % of the total endogenous protein in ileal digesta of pigs (Lien et al., 1997). Mucin proteins are continuously synthesized and resistant to digestion because they contain high density of O-linked oligosaccharides (Strous and Dekker, 1992). This mean that an increase in mucus secretion will directly increase endogenous losses of AA particularly Thr. Thus, Thr is a key AA for the integrity and immunity of the gastrointestinal tract.
Cuaron et al. (1984) showed that sows fed a diet adequate in Thr (sorghum and L-Thr based) had 20% more IgG in their plasma than sows fed the Thr-deficient diet at farrowing. Supplementing a low protein diet with 0.14% L-Thr during gestation increased milk IgG concentration at farrowing and during lactation (Hsu et al., 2001). Supplementation with L-Thr to contain a dietary Thr level higher than for optimal growth (i.e. Thr:Lys ratio of 99%) increased the production of serum antibody and IgG levels in 17-31 kg pigs challenged with bovine serum albumin (Li et al., 1999). Similarly, Wang et al. (2006) reported that the serum IgG level maximized at a higher dietary Thr level (SID Thr:Lys of 78%) while optimal growth performance was achieved at a SID Thr: Lys of 69% for 10-25 kg pigs challenged with ovalbumin injection (Fig. 5).
Cuaron et al. (1984) showed that sows fed a diet adequate in Thr (sorghum and L-Thr based) had 20% more IgG in their plasma than sows fed the Thr-deficient diet at farrowing. Supplementing a low protein diet with 0.14% L-Thr during gestation increased milk IgG concentration at farrowing and during lactation (Hsu et al., 2001). Supplementation with L-Thr to contain a dietary Thr level higher than for optimal growth (i.e. Thr:Lys ratio of 99%) increased the production of serum antibody and IgG levels in 17-31 kg pigs challenged with bovine serum albumin (Li et al., 1999). Similarly, Wang et al. (2006) reported that the serum IgG level maximized at a higher dietary Thr level (SID Thr:Lys of 78%) while optimal growth performance was achieved at a SID Thr: Lys of 69% for 10-25 kg pigs challenged with ovalbumin injection (Fig. 5).
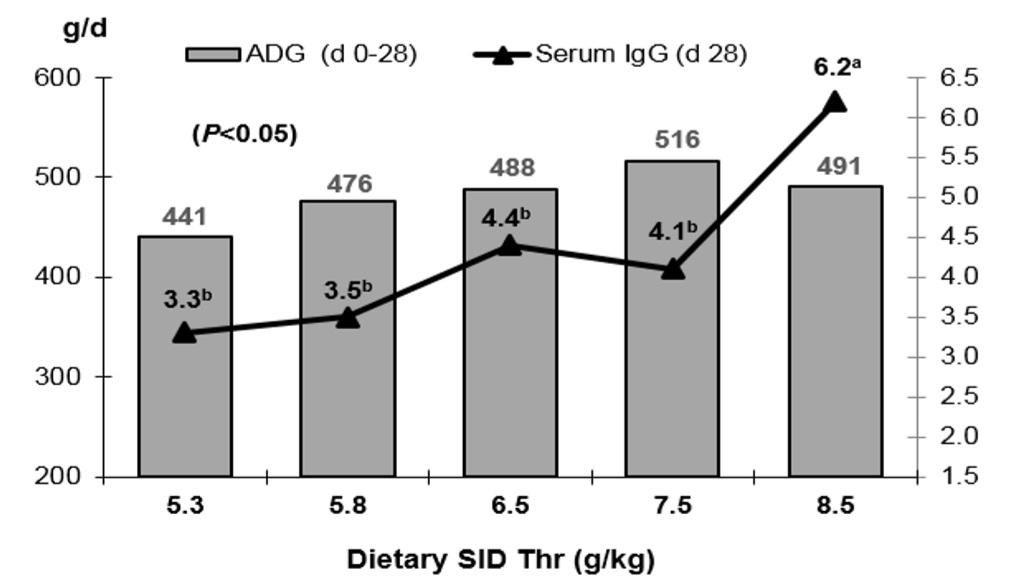
Figure 5: Effect of varying dietary Thr levels on growth and serum IgG concentration of piglets (Wang et al., 2006)
These results indicate the role of Thr in modulating animals’ immune function through its incorporation into immunoglobulin. Poor sanitary conditions of the farm can negatively affect the immune status of the animals. Indeed, Bikker et al., (2007) reported that the SID Thr: Lys ratio to optimize BW gain was higher at 71% for 25-110 kg pigs fed AGP-free diet compared with those fed AGP-added diet wherein BW gain was maximized at 65% SID Thr: Lys (Fig. 6). Jayaraman et al. (2014) also reported that poor sanitary conditions reduced growth rate and increasing SID Thr: Lys to 71% could improve gain: feed in piglets fed antibiotic-free diets.
Inadequate dietary Thr supply to piglets caused increased incidence of diarrhea, decreased mucosal weight and mucin secretion along the GIT (Law et al., 2007), and reduced villus height and villus height to crypt depth ratio in the ileum (Hamard et al., 2007). Wang et al. (2007) found that the fractional synthesis rate of jejunal mucosa and mucins were higher in weaned pigs fed the diet with the adequate Thr level (0.74 % SID Thr) compared with pigs which were pair-fed diets either an excess (1.11% SID Thr) or a deficiency of Thr (0.37% SID Thr).
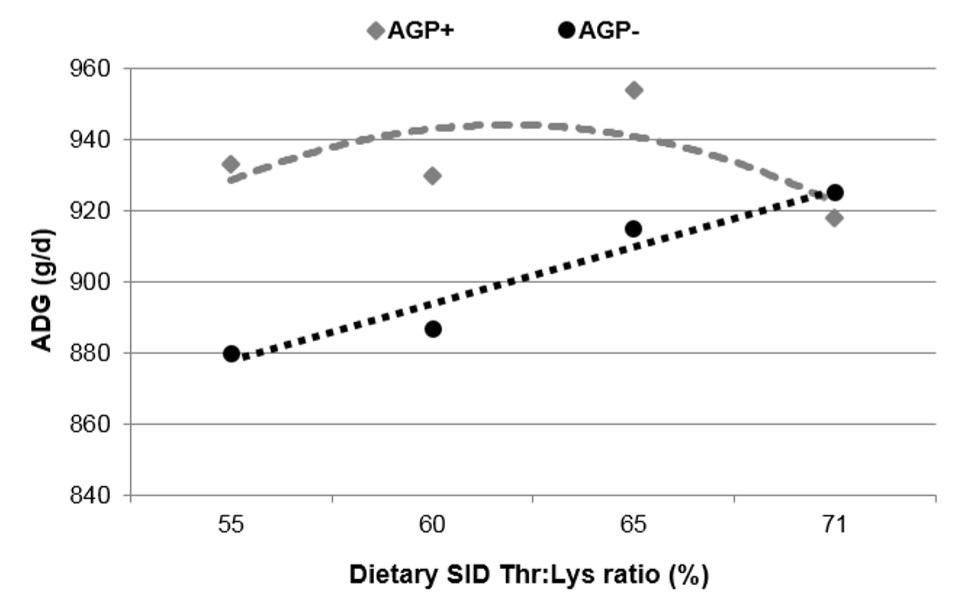 Figure 6: Effect of dietary SID Thr:Lys ratio on average daily gain (ADG) of 25-110 kg pigs fed diets with or without AGP (Bikker et al., 2006)
Figure 6: Effect of dietary SID Thr:Lys ratio on average daily gain (ADG) of 25-110 kg pigs fed diets with or without AGP (Bikker et al., 2006)Feeding wheat bran and barley based diets which are high in fiber (hemicellulose) increase endogenous losses of Thr in growing pigs compared with those fed a casein based diet (Myrie et al., 2008). With increasing dietary fiber (pectin), protein deposition was reduced to a larger extent when Thr limiting diets were fed compared with feeding a Lys-limiting diet mainly due to increased endogenous loss of Thr (Zhu and de Lange, 2001). Recently, Mathai et al. (2015) reported that the optimum SID Thr:Lys ratio for BW gain of 25 to 50 kg pigs fed high fiber diet (15% soy hulls) was higher (71%) compared with pigs fed a low fiber diet (66%), which indicates that feeding diets high in fiber increases the Thr demand for mucin production in pigs.
Glutamine:
Glutamine is the most abundant free AA in the body and milk of mammals (Wu et. al, 1996). Besides serving as a major fuel for rapidly dividing cells such as enterocytes and leukocytes of the small intestine, Gln involves in many metabolic processes including gluconeogenesis, immune response, and regulation of cellular redox state (Wu et al., 2007). There is extensively interconversion of Gln and Glu. In turn, Glu can partially substitute for Gln in several pathways, including ATP production and syntheses of arginine, ornithine, citrulline, alanine, proline, and aspartate (Reeds et al., 1997; Wu, 1998). As a precursor of Glu, Gln plays a role in the synthesis of glutathione (Reeds et al., 1997). Glutamine is also a precursor for the synthesis of nucleotides (purine and pyrimidine) that are essential for the proliferation of lymphocytes, and mucosal cells (Wu, 1998). Approximately 70% of ingested Gln is used up by the small intestine in the first pass and only 30% of Gln in the lumen enters the portal blood pool (Stoll and Burrin, 2006) highlighting the important role of Gln for maintaining intestinal barrier integrity and immune function.
Table 3: Effect of dietary glutamine on the performance of weaned pigs (Zou et al., 2006)
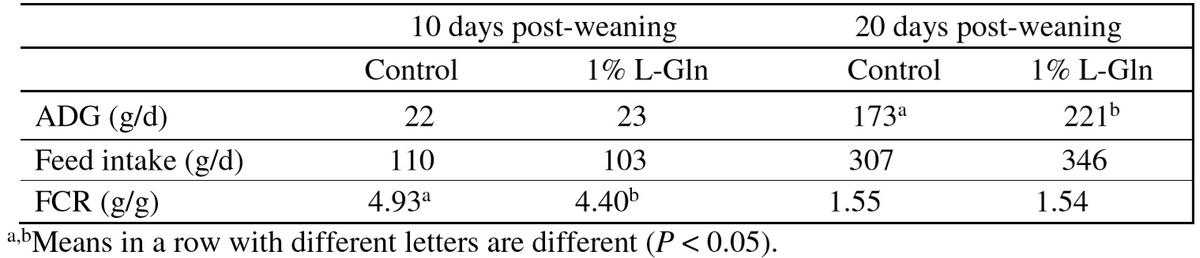
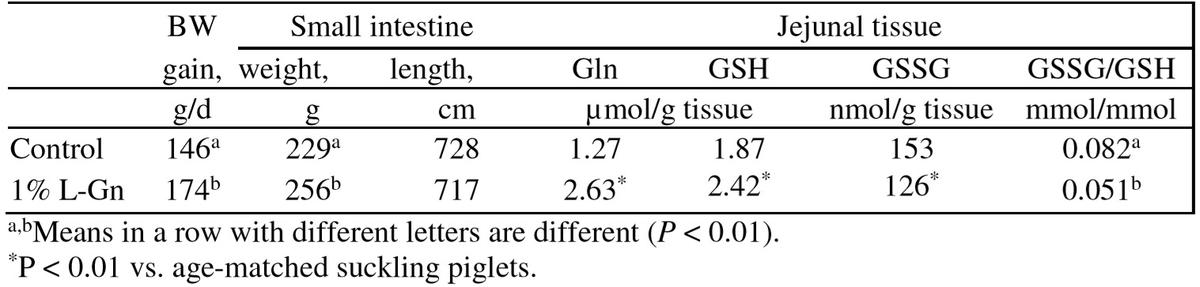
A relatively high supplementation with L-Gln at 4% increased white blood cells count and enhanced lymphocyte function in early-weaned pigs infected with E. coli (Yoo et al., 1997). Supplementation with 1% L-Gln to a diet adequate in all AA increased villus height in the jejunum, and increased gain: feed ratio in weaned piglets (Wu et al., 1996). Liu et al. (2002) also found increased jejunal villus height of weanling pigs by supplementation 1% L-Gln or L-Glu to a nutrient-adequate diet. Increased BW gain and improved feed: gain of weaned piglets was also observed by supplementation with 1% L-Gln to a diet adequate in all AA (de Abreu et al., 2010). Zou et al. (2006) also reported that supplementation with 1% L-Gln reduces diarrhea incidence and improved growth performance of weaned pigs (Table 3). Similarly, 1.0% L-Gln supplementation reduced intestinal expressions of genes that promote oxidative stress and increased intestinal glutathione concentration, small intestine growth and BW gain of weaned piglets (Wang et al., 2008; Table 4). Dietary addition with 0.8% 50:50 mix of Gln-Glu increased villus height of small intestine and BW gain of weaned pigs (Molino et al., 2012).
Table 4: Effect of dietary glutamine on performance and carcass of weaned pigs (d 21-28; Wang et al., 2008)
Arginine:
Arginine serves as a precursor for the synthesis of important molecules, including nitric oxide (NO), ornithine, citrulline, proline, glutamate, creatine and polyamines (Wu and Morris 1998). Arginine plays an important role in immunity by regulating NO synthesis by nitric-oxide synthase (NOS2) to produce antibodies by B-cells and T-cell receptor expression (De Jonge et al. 2002). Arg may function as antioxidant and ameliorate lipid peroxidation (Galli 2007). Moreover, Arg regulates the signaling via the mammalian target of rapamycin (mTOR) in the small intestine and skeletal muscle of piglets to initiate body protein synthesis (Yao et al. 2008).
Table 5: Effect of dietary Arg and Gln supply on performance of weaned pigs (Shan et al., 2012)
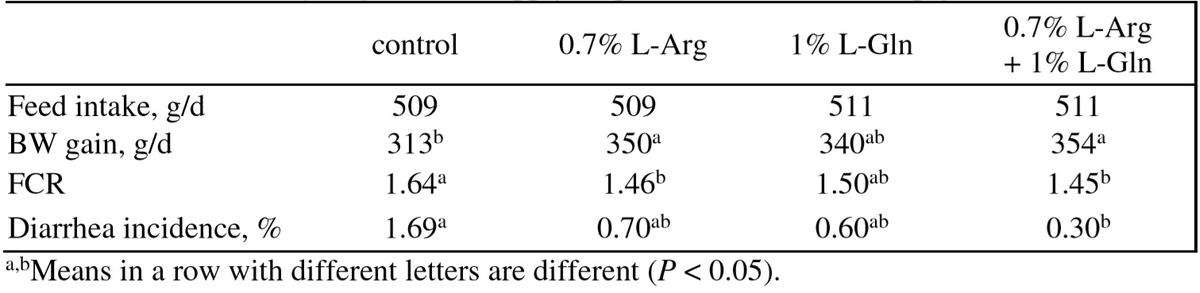
Dietary L-Arg addition enhanced immune status and gain: feed of piglets (Tan et al., 2009a) and enhanced immune status of pregnant sows (Kim et al., 2006). L-Arg addition (0.5%) to a diet adequate in all AA increased serum concentrations of albumin antibody IL-2 and IFN-γ and improved performance of immune challenged pigs(Han et al., 2009). Supplementation with 1% alleviated the impairment of gut function induced by LPS challenge in weaned pigs (Liu et al., 2009). L-Arg addition (0.6%) improved intestinal integrity and growth of weaned pigs (Wu et al., 2010). Improved gut health and BW gain of weaned pigs were obtained by supplementation of 0.7% L-Arg.HCl (0.7%) or in combination with 1% L-Gln to a diet adequate in AA (Shan et al., 2012; Table 5).
Table 6: Effect of dietary Arg supply on performance and carcass of 41-90 kg pigs (Tan et al., 2009b)
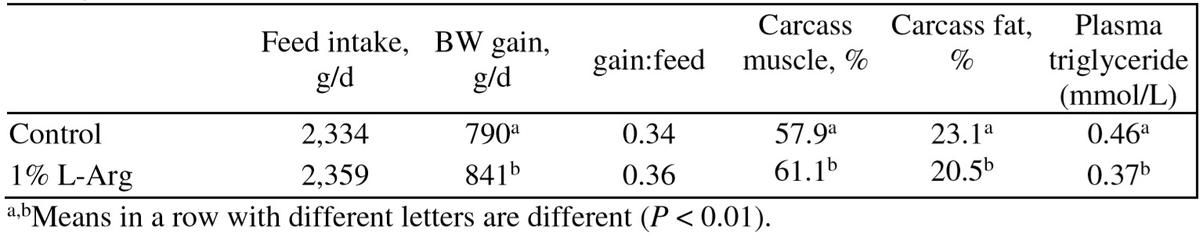
These results indicate that Arg supplementation can improve intestinal barrier function. Furthermore, dietary Arg supplementation (1.0%) increases muscle gain and reduces body fat and plasma triglyceride levels in 41-90 kg pigs (Tan et al., 2009b; Table 6). Ma et al. (2010) also reported that 1% L-Arg addition enhanced the anti-oxidative capacity and carcass quality of pigs.
Glycine:
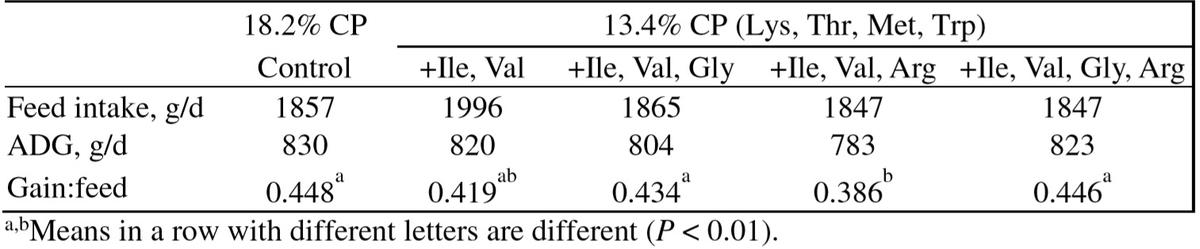
Glycine is involved in the synthesis of many important molecules, including the synthesis of GSH, serine, creatine, purine nucleotides and heme (Kim et al. 2007). Glycine plays a role in regulating the production of cytokines by leucocytes and immune function (Zhong et al. 2003). In addition, Gly itself is a potent antioxidant (Fang et al. 2002). In an in vitro study, adding Gly to Jejunal enterocytes of weaned pigs enhanced cell growth and protein synthesis and reduced attenuated apoptosis when exposed to an oxidative stress model induced by 4-hydroxynonenal(Wang et al., 2014). Glycine inhibits glutamine synthase, thereby making more Glu being available for the biosynthesis of NEAA in rats (Tate and Meister, 1971). Supplementing Gly or in combination of Gly and Arg (equal to a high protein diet) to a low-protein corn-soybean meal based diet with Lys, Met, Thr, Trp, Ile, and Val restores growth performance similar to that of 20-50 kg pigs fed a high protein control diet (Powell et al., 2011; Table 7) indicating that Gly is a potent AA for body protein synthesis.
Table 7: Effect of dietary supplementations with various amino acids on performance of 20-50 kg pigs (d 0-28; Powell et al., 2011)
Summary
This review paper summarizes the regulation of immune system including the gut immunity and their importance for achieving optimal performance and production efficiency. In addition to protein synthesis, AA are also involved in various important metabolic pathways including modulating immune system at various levels in the body. The roles of function AA for the immune functioning has received more attention during the last decade. During sub-clinical level of diseases, nutrients are prioritized to form tissues involved in immune response compromising growth, implying that the increased level of some key AA should be increased to maintain a good immunity and to reduce the negative impact on animal performance. The application of the “functional AA” concept is one of the possible solutions to maintain gut health and promote growth because it can enhance immune status, gut integrity for weaned pigs especially when sanitary and climatic conditions are challenging, and the AGPs are not or less used in the diets. The roles beyond growth and beneficial effects of dietary supplementation with the main functional AA, i.e. Met + Cys, Trp, Thr, Gln, Arg and Gly in pigs are described. Future research is warranted to quantitatively estimate the increased need of these functional AA to excerpt improvement in immunity, gut health and performance of pigs raised under sub-optimal conditions. Furthermore, there is a need to access the interaction or synergetic effect of various amino acids on animal’s immunity.
This review paper summarizes the regulation of immune system including the gut immunity and their importance for achieving optimal performance and production efficiency. In addition to protein synthesis, AA are also involved in various important metabolic pathways including modulating immune system at various levels in the body. The roles of function AA for the immune functioning has received more attention during the last decade. During sub-clinical level of diseases, nutrients are prioritized to form tissues involved in immune response compromising growth, implying that the increased level of some key AA should be increased to maintain a good immunity and to reduce the negative impact on animal performance. The application of the “functional AA” concept is one of the possible solutions to maintain gut health and promote growth because it can enhance immune status, gut integrity for weaned pigs especially when sanitary and climatic conditions are challenging, and the AGPs are not or less used in the diets. The roles beyond growth and beneficial effects of dietary supplementation with the main functional AA, i.e. Met + Cys, Trp, Thr, Gln, Arg and Gly in pigs are described. Future research is warranted to quantitatively estimate the increased need of these functional AA to excerpt improvement in immunity, gut health and performance of pigs raised under sub-optimal conditions. Furthermore, there is a need to access the interaction or synergetic effect of various amino acids on animal’s immunity.
References
Bertolo, R. F., C. Z. Chen, G. Law, P. B. Pencharz and R. O. Ball. 1998. Threonine requirement of neonatal piglets receiving total parenteral nutrition is considerably lower than that of piglets receiving an identical diet intragastrically. Journal of Nutrition 128: 1752-1759.
Bikker, P., J. Fledderus, L. Le Bellego and M. Rovers. 2007. Growth response of pigs to dietary threonine: lysine ratio is affected by the withdrawal of antimicrobial growth promoters. European Association for Animal Production 124: 557-560.
Blecha, F. 2001. Immunology. Pages 688-711 in Biology of the Domestic Pig. W. G. Pond and H. J. Mersmann, ed. Cornell Univ. Press, Ithaca, NY.
Botting, N. P. 1995. Chemistry and neurochemistry of the kynurenine pathway of tryptophan metabolism. Chem. Soc. Rev. 24: 401-412.
Bowland, J. P. 1966. in: L. K. Bustad, R. O. McClellan and M.P. Burns (Ed.): Swine in biomedical research, Frayn, USA.
Capozzalo, M. M., J. W. Resink, J. K. Htoo, J. C. Kim, C. F. M. de Lange, and C. F. Hansen. 2014.
Optimum sulfur amino acid to lysine ratio for weaner pigs infected with enterotoxigenic E. coli. J. Anim. Sci. 92 (Suppl. 2):40. (Abstr.).
Capozzalo, M. M., J.C. Kim, J.K. Htoo, C.F.M. de Lange, B.P. Mullan, J.W. Resink, C.F. Hansen, P.A. Stumbles, D.J. Hampson, N. Ferguson and J.R. Pluske. 2013. A tryptophan: lysine ratio of 0.24 gives optimum performance of weaner pigs under commercial conditions. Manipulating Pig Production XIV. Proceedings of the Fourteenth Biennial Conference of the Australasian Pig Science Association, November 24-27, 2013, Melbourne, Australia. p. 91.
Capozzalo, M. M., J. C. Kim, J. K. Htoo, C. F.M. de Lange, B. P. Mullan, C. F. Hansen, J-W. Resink, P. A. Stumbles, D. J. Hampson and J. R. Pluske. 2015. Effect of increasing the dietary tryptophan to lysine ratio on plasma levels of tryptophan, kynurenine and urea and on production traits in weaner pigs experimentally infected with an enterotoxigenic strain of Escherichia coli. Archives of Animal Nutrition 69(1):17-29.
Chen, H., J. Lin, H. Fung, L. Ho, P. Yang, W. Lee, Y. Lee and R. Chu. 2003. Serum acute phase proteins and swine health status. Can J Vet Res 67:283-290.
Cuaron, J. A., R. P. Chapple and R. A. Easter. 1984. Effect of lysine and threonine supplementation of sorghum gestation diets on nitrogen balance and plasma constituents in first-litter gilts. Journal of Animal Science 58: 631-637.
Cunningham-Rundles, S. (2002): Evaluation of the effects of nutrients on immune function. Pages 57-92 in: Calder P.C., Field C.J., Gill H. S. (eds) Nutrition and immune function. CABI Publishing. Wallingford, UK.
Colditz, I. G. 2002. Effects of the immune system on metabolism: implications for production and disease resistance in livestock. Livest Prod Sci 75: 257-268.
De Jonge, W. J., K. L. Kwikkers, A. A. Te Velde, S. J. H. van Deventer, M. A. Nolte, R. E. Mebius, J. M. Ruijter, M. C. Lamers and W. H. Lamers. 2002. Arginine deficiency affects early B cell maturation and lymphoid organ development in transgenic mice. J Clin Invest 110:1539-1548.
Dwyer, D. S. 1979. Regulation of the immune response by polyamines. Med Hypotheses 5: 11691181.
Fang, Y. Z., S. Yang and G. Wu. 2002. Free radicals, antioxidants, and nutrition. Nutrition 8, 872– 879.
Galli, F. 2007. Amino acid and protein modification by oxygen and nitrogen species. Amino Acids 32:497-499.
Grimble, R. F. 2002. Sulphur amino acids, glutathione and immune function. Pages 133-150 in: Nutrition and Immune Function; Calder PC, Field CJ, Gill HS. (Eds.); CABI Publishing: New York, NY, USA.
Hamard, A., B. Seve and N. Le Floch. 2007. Intestinal development and growth performance of early-weaned piglets fed a low-threonine diet. Animal 1: 1134-1142.
Han, J., Y. L. Liu, W. Fan, J. Chao, Y. Q. Hou, Y. L. Yin, H. L. Zhu, G. Q. Meng, and Z. Q. Che. 2009. Dietary L-arginine supplementation alleviates immunosuppression induced by cyclophosphamide in weaned pigs. Amino Acids 37:643-651.
Hsu, C. B., S. P. Cheng, J. C. Hsu and H. T. Yen. 2001. Effect of threonine addition to a low protein diet on IgG levels in body fluid of first litter sows and their piglets. Asian-Australian Journal of Animal Science 14: 1157-1163.
Hunter, E. A. and R. F. Grimble. 1994. Cysteine and methionine supplementation modulate the effect of tumour necrosis factor alpha on protein synthesis, glutathione and zinc concentration of liver and lung in rats fed a low protein diet. J Nutr 124(12): 2319-2328.
Jayaraman, B., J. K. Htoo and C.M. Nyachoti. 2014. Effects of dietary threonine:lysine ratio and sanitary conditions on performance and plasma urea nitrogen of weaned pigs fed antibiotic-free diets. J. Anim. Sci. 92 (E-Suppl. 1):218. (Abstr.).
Jayaraman, B., J. K. Htoo and C.M. Nyachoti. 2015. Effects of increasing standardized ileal digestible tryptophan:lysine ratio on performance and ileal expression of cytokine mRNA in weaned pigs challenged with Escherichia coli K88. J. Anim. Sci. 93 (Suppl. S3):201-202. (Abstr.).
Kim, S. W., R. D. Mateo, G. Wu, J. A. Carroll and I. Shinzato. 2006. Dietary L-arginine supplementation affects immune status of pregnant gilts. FASEB Journal. 20:A424. Kim, S. W., R. D. Mateo, Y. L. Yin and G. Wu. 2007. Functional amino acids and fatty acids for enhancing production performance of sows and piglets. Asian-Aust J Anim Sci 20, 295-306.
Kim, J. C., B. P. Mullan, B. Frey, H. G. Payne, and J. R. Pluske. 2012. Whole body protein deposition and plasma amino acid profiles in growing and/or finishing pigs fed increasing levels of sulfur amino acids with and without Escherichia coli lipopolysaccharide challenge. J. Anim. Sci. 90:362-365.
Kuby, J. 1994. Immunology. 2nd Ed. W. H. Freeman and Company. New York, New York.
Law, G. K., R. F. Bertolo, A. Adjiri-Awere, P. B. Pencharz and R. O. Ball. 2007. Adequate oral threonine is critical for mucin production and gut function in neonatal piglets. American Journal of Physiology-Gastrointestinal and Liver Physiology 292: G1293-G1301.
Le Floc’h, N., D. Melchior, L. Le Bellego, J. J. Matte, and B. Sève. 2007. Does sanitary status have an effect on tryptophan requirement for growth of postweaning piglets? (in French, with English abstract). J. Rech. Porcine France 39: 125-132.
Le Floc’h, N., D. Melchior, and B. Seve. 2008. Dietary tryptophan helps preserve tryptophan homeostasis in pigs suffering from lung inflammation. J. Anim. Sci. 86:3473-3479.
Le Floc’h, N., and B. Seve. 2007. Biological roles of tryptophan and its metabolism. Potential implications for pig feeding. Livest. Sci. 112:23-32.
Li, D. F., C. T. Xiao, S. Y. Qiao, J. H. Zhang, E. W. Johnson and P. A. Thacker. 1999. Effects of dietary threonine on performance, plasma parameters and immune function of growing pigs. Animal Feed Science and Technology 78: 179-188.
Li, P., Y-L. Yin, D. Li, S. W. Kim and G. Wu. 2007. Amino acids and immune function. Br J Nutr 98, 237-252.
Lien, K. A., W. C. Sauer and M. Fenton. 1997. Mucin output in ileal digesta of pigs fed a proteinfree diet. Zeitschrift für Ernährungswissenschaft 36: 182-190.
Litvak, N., J. K. Htoo, C. F. M. de Lange. 2013a. Restricting sulfur amino acid intake in growing pigs challenged with lipopolysaccharides decreases plasma protein and albumin synthesis. Can J Anim Sci 93, 505-515.
Litvak, N., A. Rakhshandeh, J. K. Htoo, C. F. M. de Lange. 2013b. Immune system stimulation increases the optimal dietary methionine to methionine plus cysteine ratio in growing pigs. J Anim Sci 91, 4188-4196.
Liu, Y., J. Han, J. Huang, X. Wang, F. Wang and J. Wang. 2009. Dietary L-arginine supplementation improves intestinal function in weaned pigs after an Escherichia coli lipopolysaccharide challenge. Asian-Aust. J. Anim. Sci.22 (12):1667-1675.
Liu, T., J. Peng, Y. Xiong, S. Zhou and X. Cheng. 2002. Effects of dietary glutamine and glutamate supplementation on small intestinal structure, active absorption and DNA, RNA concentrations in skeletal muscle tissue of weaned piglets during d 28 to 42 of age. Asian-Aust. J. Anim. Sci. 15 (2): 238-242.
Lu, S. C. 2009. Regulation of glutathione synthesis. Mol Aspects Med 30: 42-59.
Ma, X., Y. Lin, Z. Jiang, C. Zheng, G. Zhou, D. Yu, T. Cao, J. Wang, and F. Chen. 2010. Dietary arginine supplementation enhances antioxidative capacity and improves meat quality of finishing pigs. Amino Acids 38:95-102.
Molino, J. P., J. L. Donzele, R. F. M. de Oliveira, A. Saraiva, D. Haese, E. L. Fortes and M. F. de Sauza. 2012. L-glutamine and L-glutamate in diets with different lactose levels for piglets weaned at 21 days of age. R. Bras. Zootec. 41(1):98-105.
Myrie, S. B., R. F. Bertolo, W. C. Sauer and R. O. Ball. 2008. Effect of common antinutritive factors and fibrous feedstuffs in pig diets on amino acid digestibilities with special emphasis on threonine. Journal of Animal Science 86: 609-619.
Mathai, J. K., J. K. Htoo, J. Thomson, K. J. Touchette, and H. H. Stein (2015). Effects of dietary fiber on the optimum threonine:lysine ratio for 25- to 50-kg gilts. J. Anim. Sci. 93 (Suppl. s3):298. (Abstr.).
Mayer, L. 2000. Mucosal immunity and gastrointestinal antigen processing. J. Pediatr. Gastroenterol. Nutr. 30(Suppl.):S4-S12.
Melchior, D., B. Sève, and N. Le Floc’h. 2004. Chronic lung inflammation affects plasma amino acid concentrations in pigs. J. Anim. Sci. 82: 1091-1099.
Obled, C. 2003. Amino acid requirements in inflammatory states. Can J Anim Sci 83(3): 365-373.
Oswald, I. P. 2006. Role of intestinal epithelial cells in the innate immune defence of the pig intestine. Veterinary Research 37: 359-368.
Parkin, J. and B. Cohen. 2001. An overview of the immune system. Lancet 357, 1777-1789.
Pastorelli, H., J. van Milgen, P. Lovatto and L. Montagne. 2012. Meta-analysis of feed intake and growth responses of growing pigs after a sanitary challenge. Animal 6, 952-961. Powell, S., T. B. Bidner, R. L. Payne and L. L. Southern. 2011. Growth performance of 20- to 50kilogram pigs fed low-crude-protein diets supplemented with histidine, cystine, glycine, glutamic acid, or arginine. J Anim Sci 89:3643-3650.
Rakhshandeh, A., J. K. Htoo and C. F. M. de Lange. 2010. Immune system stimulation of growing pigs does not alter apparent ileal amino acid digestibility but reduces the ratio between whole body nitrogen and sulfur retention. Livest Sci 134, 21-23.
Rakhshandeh, A., J. K. Htoo, N. Karrow, S. P. Miller and C. F. M. de Lange. 2014. Impact of immune system stimulation on ileal nutrient digestibility and utilization of methionine plus cysteine intake for whole body protein deposition in growing pigs. Br J Nutr 111, 101-110.
Reeds, P. J., D. G. Burrin, B. Stoll, F. Jahoor, L. Wykes, J. Henry and M. E. Frazer. 1997. Enteral glutamate is the preferential source for mucosal glutathione synthesis in fed piglets. American Journal of Physiology 273: E408-E415.
Reeds, P. J. and F. Jahoor. 2001. The amino acid requirements of disease. Clin Nutr 20 (S1): 1522.
Ridder, K. de, C. L. Levesque, J. K. Htoo, and C. F. M. de Lange. 2012. Immune system stimulation reduces the efficiency of tryptophan utilization for body protein deposition in growing pigs. J. Anim. Sci. 90:3485-3491.
Roth, E. 2007. Immune and cell modulation by amino acids. Clin Nutr 26: 535-544.
Shao L., Serrano D., Mayer, L. 2001: The role of epithelial cells in immune regulation in the gut, Semin. Immunol. 13:163-176.
Shan, Y., A. Shan, J. Li, and C. Zhou. 2012. Dietary supplementation of arginine and glutamine enhances the growth and intestinal mucosa development of weaned piglets. Livestock Science 150:369–373.
Stadnyk A.W. 2002. Intestinal epithelial cells as a source of inflammatory cytokines and chemokines, Can. J. Gastroenterol. 16:241-246.
Stoll, B. and D. G. Burri. (2006. Measuring splanchnic amino acid metabolism in vivo using stable isotopic tracers. Journal of Animal Science 84 (Suppl. 13): E60-72.
Strous, G. J. and J. Dekker. 1992. Mucin-type glycoproteins. Critical review in Biochemistry and Biology 27: 57-92.
Tan B., X.G. Li, X. Kong, R. Huang, Z. Ruan, K. Yao, Z. Deng, M. Xie, I. Shinzato, Y. Yin and G. Wu. 2009a. Dietary L-arginine supplementation enhances the immune status in early-weaned piglets. Amino Acids 37:323-331.
Tan, B., Yin Y., Z. Liu, X. Li., H. Xu., X. Kong, R. Huang, W. Tang, I. Shinzato, S. B. Smith and G. Wu. 2009b. Dietary L-arginine supplementation increases muscle gain and reduces body fat mass in growing-finishing pigs. Amino Acids. 37:169-175.
Tan, B., M. Xie and Y. Yin. 2013. Amino acids and immune functions. Pages 175-185 in: Nutritional and Physiological Functions of Amino Acids in Pigs. Eds. F. Blachier, G. Wu and Y. Yin. Heidelberg, Germany: Springer. Tate, S. S. and A. Meister. 1971. Regulation of rat liver glutamine synthase: Activation by αketoglutarate and inhibition by glycine, alanine, and carbamyl phosphate. Proc. Natl. Acad. Sci. USA 68:781-785.
Stoll, B. 2006. Intestinal uptake and metabolism of threonine: Nutritional Impact. Advances in Pork Production 17: 257-263.
Wang, X., S. Y. Qiao, M. Liu and Y. X. Ma. 2006. Effects of graded levels of true ileal digestible threonine on performance, serum parameters and immune function of 10-25 kg pigs. Animal Feed Science and Technology 129: 264-278.
Wang, X., S. Y. Qiao, Y. L. Yin, L. Yue, Z. Wang and G. Wu. 2007. A deficiency or excess of dietary threonine reduces protein synthesis in jejunum and skeletal muscle of young pigs. Journal of Nutrition 137: 1442-1446.
Wang, J. J., L. X. Chen, P. Li, X. L. Li, H. J. Zhou, F. L. Wang, D. F. Li , Y. L. Yin and G. Wu. 2008. Gene expression is altered in piglet small intestine by weaning and dietary glutamine supplementation. J Nutr 138:1025-1032.
Wang, W., Z. Wu., G. Lin., S. Hu, B. Wang, Z. Dai and G. Wu. 2014. Glycine stimulates protein synthesis and inhibits oxidative stress in pig small intestinal epithelial cells. J. Nutr. 144(10):15401548.
Wu, G., S. A. Meier and D. A. Knabe. 1996. Dietary glutamine supplementation prevents jejunal atrophy in weaned pigs. J Nutr 126, 2578-2584.
Wu, G. 1998. Intestinal mucosal amino acid catabolism. J. Nutr. 128:1249-1252.
Wu, G. Y. and S. M. Jr. Morris. 1998. Arginine metabolism: nitric oxide and beyond. Biochem J 336:1-17.
Wu, G., Y. Fang, S. Yang, J. R. Lupton and N. D. Turner. 2004. Glutathione metabolism and its implications for health. J Nutr 134: 489-492.
Wu, G., W. B. Fuller, T. A. Davis, L. A. Jaeger,G. A. Johnson, S. W. Kim, D. A. Knabe, C. J. Meininger, T. E. Spencer and Y-L Yin. 2007. Important roles for the arginine family of amino acids in swine nutrition and production. Livestock Sci. 112:8-22.
Wu, G. 2009. Amino acids: metabolism, functions, and nutrition. Amino Acids. 37:1-17.
Wu, G. 2010. Functional amino acids in growth, reproduction, and health. Adv. Nutr. 1: 31-37.
Yao K, Y. L. Yin, W. Y. Chu, Z. Liu, D. Deng, T. Li, R. Huang, J. Zhang, B. Tan, W. Wang and G. Wu. 2008. Dietary arginine supplementation increases mTOR signaling activity in skeletal muscle of neonatal pigs. J Nutr 138:867-872.
Yoneda, J., A. Andou and K. Takehana. 2009. Regulatory roles of amino acids in immune response. Curr Rheumatol Rev 5:252-258.
Yoo, S. S., C. J. Field and M. I. McBurney. 1997. Glutamine supplementation maintains intramuscular glutamine concentrations and normalizes lymphocyte function in infected early weaned pigs. J Nutr 127, 2253–2259.
Zhang, G. J., Q. L. Song, C. Y. Xie, L. C. Chu, P. A. Thacker, J. K. Htoo, and S. Y. Qiao. 2012. Estimation of the ideal ratio of standardized ileal digestible tryptophan to lysine for growing pigs fed low crude protein diets supplemented with crystalline amino acids. Livestock Sci. 149:260266.
Zhang, G. J., P.A. Thacker, J.K. Htoo, S.Y. Qiao (2015): Optimum proportion of standardized ileal digestible sulfur amino acid to lysine to maximize the performance of 25–50 kg growing pigs fed reduced crude protein diets fortified with amino acids. Czech J. Anim. Sci., 60(7): 302–310. Zhong, Z., M. D. Wheeler, X. L. Li, M. Froh, P. Schemmer, M. Yin, H. Bunzendaul, B. Bradford and J. J. Lemasters. 2003. Glycine: a novel anti-inflammatory, immunomodulatory, and cytoprotective agent. Curr Opin Clin Nutr Metab Care 6, 229-240.
Zhu, C. L., M. Rademacher and C. F. M. de Lange. 2005. Increasing dietary pectin level reduces utilization of digestible threonine intake, but not lysine intake, for body protein deposition in growing pigs. Journal of Animal Science 83: 1044-1053.
Zou, X. T., G. H. Zheng, X. K. J. Fang and J. F. Fang. 2006. Effects of glutamine on growth performance of weanling piglets. Czech J. Anim. Sci. 51 (10):444-448.
Related topics:
Authors:


Influencers who recommended :
Dr. Andreas Lemme, Caroline Gonzalez-Vega and 1 moreRecommend
Comment
Share

Would you like to discuss another topic? Create a new post to engage with experts in the community.




