Effects of feeding high-protein corn distillers dried grains and a mycotoxin mitigation additive on growth performance, carcass characteristics, and pork fat quality of growing–finishing pigs
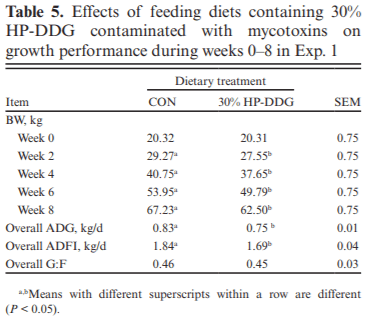
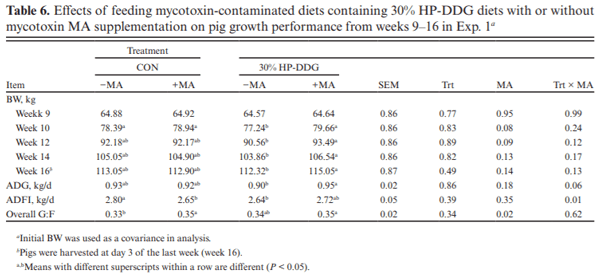
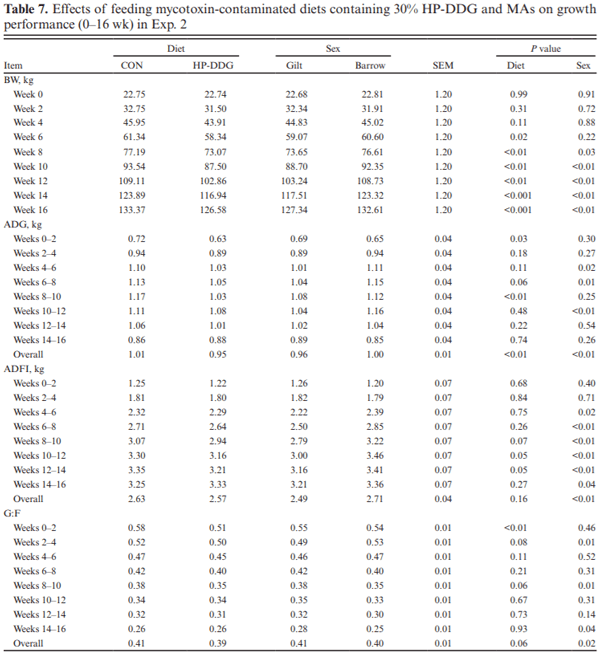
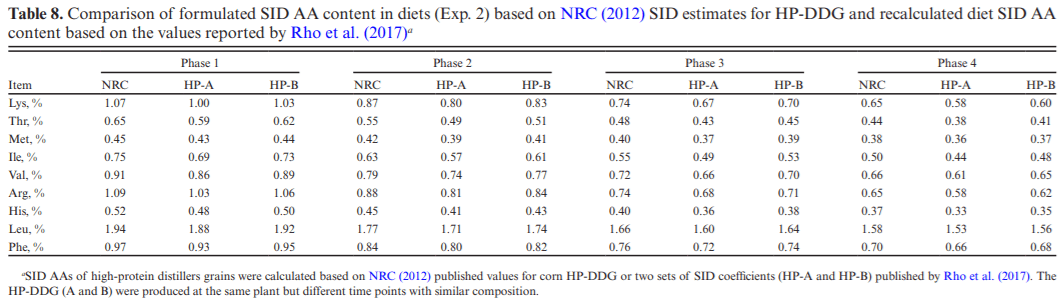
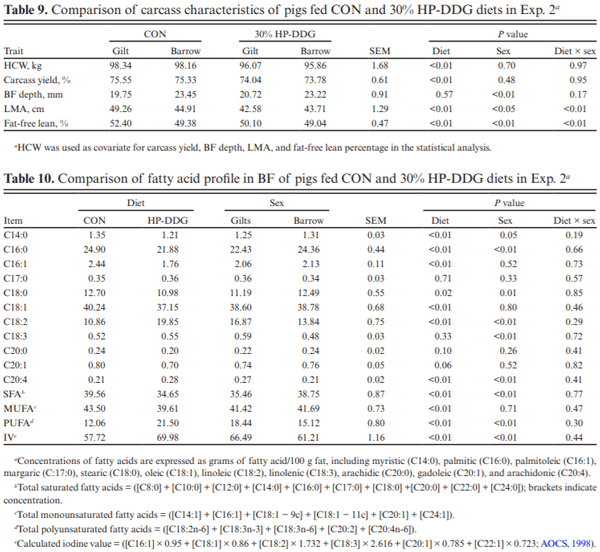
Two experiments investigated the effects of feeding diets containing 30% of novel high-protein distillers dried grains (HP-DDG) sources to growing–finishing pigs on growth performance, carcass characteristics, and pork fat quality. A four-phase feeding program was used in both experiments, and diets within phases were formulated based on National Research Council (NRC; 2012) recommendations for metabolizable energy and standardized ileal digestible amino acid content of HP-DDG. In Exp. 1, a total of 144 pigs (body weight [BW] = 20.3 ± 1.6 kg) were fed either corn-soybean meal control diets (CON) or 30% HP-DDG diets (HP-DDG) containing 0.7 mg/kg deoxynivalenol (DON), 0.1 mg/ kg fumonisins (FUM), and 56 μg/kg zearalenone (ZEA) for 8 wk. On week 9, a mycotoxin mitigation additive (MA) was added to CON and HP-DDG diets, resulting in a 2 × 2 factorial arrangement of treatments consisting of: CON, CON + MA, HP-DDG, and HP-DDG + MA. Pigs fed HP-DDG had lower (P < 0.01) average daily gain (ADG) and average daily feed intake (ADFI) compared with those fed CON during the first 8 wk. After MA was added to diets, pigs fed HP-DDG diets without MA had lower (P < 0.05) overall ADG than those fed HP-DDG + MA and less (P < 0.05) final BW than pigs fed CON or CON + MA. Adding MA to HP-DDG diets containing relatively low concentrations of mycotoxins was effective in restoring growth performance comparable to feeding CON. In Exp. 2, a different source of HP-DDG was used, and mycotoxin MAs were added to all diets at the beginning of the trial. A total of 144 pigs (BW = 22.7 ± 2.3 kg) were fed either a corn-soybean meal control diet or a 30% HP-DDG diet containing 0.5 mg/kg DON and 0.8 mg/kg FUM for 16 wk. Pigs fed HP-DDG diets had less (P < 0.01) final BW and ADG than pigs fed CON, but there were no differences in ADFI. Feeding the HP-DDG diets reduced (P < 0.01) hot carcass weight, carcass yield, longissimus muscle area (LMA), and percentage of carcass fat-free lean compared with pigs fed CON but did not affect backfat (BF) depth. Pigs fed HP-DDG had less (P < 0.01) saturated fatty acid (SFA) and monounsaturated fatty acid (MUFA) content and greater (P < 0.01) poly-unsaturated fatty acid (PUFA) and iodine value in BF than pigs fed CON. These results suggest that feeding diets containing relatively low concentrations of co-occurring mycotoxins can be detrimental to growth performance, and the addition of MA alleviated the growth reduction. Feeding 30% HP-DDG reduced BW, ADG, carcass yield, LMA, and percentage of fat-free lean of grow-ing–finishing pigs but yielded acceptable pork fat quality.
Key words: carcass characteristics, growing–finishing pigs, growth performance, high-protein distillers dried grains, mycotoxins, pork fat quality.
Alizadeh, A., S. Braber, P. Akbari, J. Garssen, and J. Fink-Gremmels. 2015. Deoxynivalenol impairs weight gain and affects markers of gut health after low-dose, short-term exposure of growing pigs. Toxins (Basel). 7:2071–2095. doi:10.3390/toxins7062071.
AOAC. 2012. Official methods of analysis. 19th ed. Assoc. Off. Anal. Chem., Washington, DC.
AOCS. 1998. Official methods and recommended practices of the AOCS. 5th ed. American Oil Chemists’ Society, Champaign, IL.
Apple, J. K., C. V. Maxwell, D. C. Brown, K. G. Friesen, R. E. Musser, Z. B. Johnson, and T. A. Armstrong. 2004. Effects of dietary lysine and energy density on perfor-mance and carcass characteristics of finishing pigs fed ractopamine. J. Anim. Sci. 82:3277–3287. doi:10.2527/20 04.82113277x.
Berthiller, F., C. Crews, C. Dall’Asta, S. D. Saeger, G. Haesaert, P. Karlovsky, I. P. Oswald, W. Seefelder, G. Speijers, and J. Stroka. 2013. Masked mycotoxins: a review. Mol. Nutr. Food Res. 57:165–186. doi:10.1002/mnfr.201100764.
Boyd, R. D., M. E. Johnston, K. Scheller, A. A. Sosnicki, and E. R. Wilson. 1997. Relationship between dietary fatty acid profile and body fat composition in growing pigs. Memo 153. PIC Tech., Franklin, KY.
Bracarense, A. P., J. Lucioli, B. Grenier, G. Drociunas Pacheco, W. D. Moll, G. Schatzmayr, and I. P. Oswald. 2012. Chronic ingestion of deoxyni-valenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intes-tine of piglets. Br. J. Nutr. 107:1776–1786. doi:10.1017/ S0007114511004946.
Cemin, H. S., M. D. Tokach, S. S. Dritz, J. C. Woodworth, J. M. DeRouchey, and R. D. Goodband. 2019. Meta-regression analysis to predict the influence of branched-chain and large neutral amino acids on growth performance of pigs. J. Anim. Sci. 97:2505–2514. doi:10.1093/jas/skz118.
Chen, H. Y., A. J. Lewis, P. S. Miller, and J. T. Yen. 1999. The effect of infusion of urea into the vena cava on feed intake of finishing gilts. J. Anim. Sci. 77:3248–3252. doi:10.2527 /1999.77123248x.
Cromwell, G. L., T. R. Cline, J. D. Crenshaw, T. D. Crenshaw, R. C. Ewan, C. R. Hamilton, A. J. Lewis, D. C. Mahan, E. R. Miller, and J. E. Pettigrew. 1993. The dietary protein and(or) lysine requirements of barrows and gilts. NCR-42 Committee on Swine Nutrition. J. Anim. Sci. 71:1510– 1519. doi:10.2527/1993.7161510x.
Eriksen, G. S., and H. Pettersson. 2004. Toxicological evaluation of trichothecenes in animal feed. Anim. Feed Sci. Technol. 114:205–239. doi:10.1016/j.anifeedsci.2003.08.008.
Etienne, M., and Y. Wache. 2008. Biological and physio-logical effects of deoxynivalenol (DON) in the pig. In: I. P. Oswald and I. Taranu, editors, Mycotoxins in farm animals. Transworld Research Network, Kerala, India. p. 113–130.
Farnworth, E. R., and J. K. G. Kramer. 1987. Fat metabolism in growing swine: a review. Can. J. Anim. Sci. 67:301–318. doi: 10.4141/cjas87-029.
Frobose, H. L., J. A. Erceg, S. Q. Fowler, M. D. Tokach, J. M. DeRouchey, J. C. Woodworth, S. S. Dritz, and R. D. Goodband. 2016. The progression of deoxyniva-lenol-induced growth suppression in nursery pigs and the potential of an algae-modified montmorillonite clay to mitigate these effects. J. Anim. Sci. 94:3746–3759. doi:10.2527/jas.2016-0663.
Frobose, H. L., E. D. Fruge, M. D. Tokach, E. L. Hansen, J. M. DeRouchey, S. S. Dritz, R. D. Goodband, and J. L. Nelssen. 2015. The effects of deoxynivalenol-con-taminated corn dried distillers grains with solubles in nursery pig diets and potential for mitigation by commer-cially available feed additives. J. Anim. Sci. 93:1074–1088. doi:10.2527/jas.2013-6883.
Grenier, B., and I. Oswald. 2011. Mycotoxin co-contamination of food and feed: meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 4:285–313. doi:10.3920/WMJ2011.1281.
Gutierrez, N. A., D. Y. Kil, Y. Liu, J. E. Pettigrew, and H. H. Stein. 2014. Effects of co-products from the corn-ethanol industry on body composition, retention of protein, lipids and energy, and on the net energy of diets fed to growing or finishing pigs. J. Sci. Food Agric. 94:3008–3016. doi:10.1002/jsfa.6648.
Hendel, E. G., P. N. Gott, G. R. Murugesan, and T. Jenkins. 2017. Survey of mycotoxins in 2016 United States corn. J. Anim. Sci. 95:16–17. doi:10.2527/asasann.2017.033.
Htoo, J. K., K. Männer, and J. Zentek. 2017. Excess dietary leucine level increases the optimal dietary isoleucine-to-ly-sine ratio in 8- to 21-kilogram pigs. J. Anim. Sci. 95:195– 196. doi:10.2527/asasann.2017.396.
Huff, W. E., L. F. Kubena, R. B. Harvey, and J. A. Doerr. 1988. Mycotoxin interactions in poultry and swine. J. Anim. Sci. 66:2351–2355. doi:10.2527/jas1988.6692351x.
Jacela, J. Y., H. L. Frobose, J. M. DeRouchey, M. D. Tokach, S. S. Dritz, R. D. Goodband, and J. L. Nelssen. 2010. Amino acid digestibility and energy concentration of high-protein corn dried distillers grains and high-protein sorghum dried distillers grains with solubles for swine. J. Anim. Sci. 88:3617–3623. doi:10.2527/jas.2010-3098.
Just, A. 1982. The influence of crude fibre from cereals on the net energy value of diets for growth in pigs. Livest. Prod. Sci. 9:569–580. doi:10.1016/0301-6226(82)90004–5.
Kass, M. L., P. J. Van Soest, W. G. Pond, B. Lewis, and R. E. McDowell. 1980. Utilization of dietary fiber from alfalfa by growing swine. I. Apparent digestibility of diet components in specific segments of the gastroin-testinal tract. J. Anim. Sci. 50:175–191. doi:10.2134/ jas1980.501175x.
Kim, B. G., G. I. Petersen, R. B. Hinson, G. L. Allee, and H. H. Stein. 2009. Amino acid digestibility and energy con-centration in a novel source of high-protein distillers dried grains and their effects on growth performance of pigs. J. Anim. Sci. 87:4013–4021. doi:10.2527/jas.2009-2060.
Kwon, W. B., K. J. Touchette, A. Simongiovanni, K. Syriopoulos, A. Wessels, and H. H. Stein. 2019. Excess dietary leucine in diets for growing pigs reduces growth performance, biological value of protein, protein reten-tion, and serotonin synthesis1. J. Anim. Sci. 97:4282–4292. doi:10.1093/jas/skz259.
Nagl, V., B. Woechtl, H. E. Schwartz-Zimmermann, I. Hennig-Pauka, W. D. Moll, G. Adam, and F. Berthiller. 2014. Metabolism of the masked mycotoxin deoxynivale-nol-3-glucoside in pigs. Toxicol. Lett. 229:190–197. doi:10.1016/j.toxlet.2014.06.032.
NGFA. 2019. A guide for grain elevators, feed manufactur-ers, grain processors and exporters. FDA Mycotoxin Regulatory Guidance. Arlington (VA): National Grain and Feed Association. https://www.ngfa.org/newsletter [Accessed January 5, 2020].
NPPC. 2000. Pork composition & quality assessment proce-dures. National Pork Producers Council, Des Moines, IA NRC. 2012. Nutrient requirements of swine. 11th rev. ed. Natl. Acad. Press, Washington, DC.
Patience, J. F., A. J. Myers, S. Ensley, B. M. Jacobs, and D. Madson. 2014. Evaluation of two mycotoxin mitiga-tion strategies in grow-finish swine diets containing corn dried distillers grains with solubles naturally contam-inated with deoxynivalenol. J. Anim. Sci. 92:620–626. doi:10.2527/jas.2013-6238.
Pierron, A., S. Mimoun, L. S. Murate, N. Loiseau, Y. Lippi, A. P. Bracarense, L. Liaubet, G. Schatzmayr, F. Berthiller, W. D. Moll, et al. 2016. Intestinal toxicity of the masked mycotoxin deoxynivalenol-3-β-D-glu-coside. Arch. Toxicol. 90:2037–2046. doi:10.1007/ s00204-015-1592-8.
Rho, Y., C. Zhu, E. Kiarie, and C. F. M. de Lange. 2017. Standardized ileal digestible amino acids and digestible energy contents in high-protein distiller’s dried grains with solubles fed to growing pigs. J. Anim. Sci. 95:3591–3597. doi:10.2527/jas.2017.1553.
Schaafsma, A. W., V. Limay-Rios, D. E. Paul, and J. David Miller. 2009. Mycotoxins in fuel ethanol co-prod-ucts derived from maize: a mass balance for deoxyniva-lenol. J. Sci. Food Agric. 89:1574–1580. doi: 10.1002/ jsfa.3626.
Seabolt, B. S., E. van Heugten, S. W. Kim, K. D. Ange-van Heugten, and E. Roura. 2010. Feed preferences and performance of nursery pigs fed diets containing various inclusion amounts and qualities of distillers coprod-ucts and flavor. J. Anim. Sci. 88:3725–3738. doi:10.2527/ jas.2009-2640.
Shawk, D. J., S. S. Dritz, R. D. Goodband, M. D. Tokach, J. C. Woodworth, and J. M. DeRouchey. 2019. Effects of sodium metabisulfite additives on nursery pig growth. Transl. Anim. Sci. 3:143–152. doi: 10.1093/tas/txy098.
Shawk, D. J., S. S. Dritz, M. D. Tokach, J. C. Woodworth, B. D. Goodband, and J. M. DeRouchey. 2018. Effects of added defusion or feed aid on finishing pig growth perfor-mance from 20 to 110 kg. J. Anim. Sci. 96:123–124. doi: 10.1093/jas/sky073.229.
Smith, M. C., S. Madec, E. Coton, and N. Hymery. 2016. Natural co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins (Basel). 8:94. doi:10.3390/toxins8040094.
Smith, J. W. II, M. D. Tokach, P. R. O’Quinn, J. L. Nelssen, and R. D. Goodband. 1999. Effects of dietary energy den-sity and lysine: calorie ratio on growth performance and carcass characteristics of growing-finishing pigs. J. Anim. Sci. 77:3007–3015. doi:10.2527/1999.77113007x.
Speijers, G. J. A., and M. H. M. Speijers. 2004. Combined toxic effects of mycotoxins. Toxicol. Lett. 153:91–98. doi: 10.1016/j.toxlet.2004.04.046.
Stein, H. H., and G. C. Shurson. 2009. Board-invited review: The use and application of distillers dried grains with solubles in swine diets. J. Anim. Sci. 87:1292–1303. doi:10.2527/jas.2008-1290.
Thaler, B., and D. E. Reese. 2010. Utilization of weather stressed feedstuffs in swine. National Swine Nutrition Guide. U.S. Pork Center of Excellence. Ames, IA. p. 207–213.
Thanh, B. V. L., M. Lessard, Y. Chorfi, and F. Guay. 2015. The efficacy of anti-mycotoxin feed additives in prevent-ing the adverse effects of wheat naturally contaminated with Fusarium mycotoxins on performance, intestinal barrier function and nutrient digestibility and retention in weanling pigs. Can. J. Anim. Sci. 95:197–209. doi:10.4141/ cjas-2014–126.
Varga, E., T. Glauner, R. Köppen, K. Mayer, M. Sulyok, R. Schuhmacher, R. Krska, and F. Berthiller. 2012. Stable isotope dilution assay for the accurate determi-nation of mycotoxins in maize by UHPLC-MS/MS. Anal. Bioanal. Chem. 402:2675–2686. doi:10.1007/ s00216-012-5757-5.
Whitney, M. H., G. C. Shurson, L. J. Johnston, D. M. Wulf, and B. C. Shanks. 2006. Growth performance and carcass characteristics of grower-finisher pigs fed high-quality corn distillers dried grain with solubles originating from a modern Midwestern ethanol plant. J. Anim. Sci. 84:3356– 3363. doi:10.2527/jas.2006-099.
Widmer, M. R., L. M. McGinnis, D. M. Wulf, and H. H. Stein. 2008. Effects of feeding distillers dried grains with solu-bles, high-protein distillers dried grains, and corn germ to growing-finishing pigs on pig performance, carcass quality, and the palatability of pork. J. Anim. Sci. 86:1819–1831. doi:10.2527/jas.2007-0594.
Wiltafsky, M. K., M. W. Pfaffl, and F. X. Roth. 2010. The effects of branched-chain amino acid interactions on growth performance, blood metabolites, enzyme kinetics and transcriptomics in weaned pigs. Br. J. Nutr. 103:964– 976. doi:10.1017/S0007114509992212.
Wiseman, T. G., D. C. Mahan, S. J. Moeller, J. C. Peters, N. D. Fastinger, S. Ching, and Y. Y. Kim. 2007. Phenotypic measurements and various indices of lean and fat tissue development in barrows and gilts of two genetic lines from twenty to one hundred twenty-five kilograms of body weight. J. Anim. Sci. 85:1816–1824. doi:10.2527/ jas.2006-406.
Wood, J. D., R. I. Richardson, G. R. Nute, A. V. Fisher, M. M. Campo, E. Kasapidou, P. R. Sheard, and M. Enser. 2004. Effects of fatty acids on meat quality: a review. Meat Sci. 66:21–32. doi:10.1016/S0309-1740(03)00022-6.
Wu, F., L. J. Johnston, P. E. Urriola, and G. C. Shurson. 2016a. Pork fat quality of pigs fed distillers dried grains with sol-ubles with variable oil content and evaluation of iodine value prediction equations. J. Anim. Sci. 94:1041–1052. doi:10.2527/jas.2015-9593.
Wu, F., L. J. Johnston, P. E. Urriola, and G. C. Shurson. 2016b. Pork fat quality of pigs fed distillers dried grains with sol-ubles with variable oil content and evaluation of iodine value prediction equations. J. Anim. Sci. 94:1041–1052. doi:10.2527/jas.2015-9593.
Xu, G., S. K. Baidoo, L. J. Johnston, D. Bibus, J. E. Cannon, and G. C. Shurson. 2010. Effects of feeding diets contain-ing increasing content of corn distillers dried grains with solubles to grower-finisher pigs on growth performance, carcass composition, and pork fat quality. J. Anim. Sci. 88:1398–1410. doi:10.2527/jas.2008-1404.
Yang, Z., P. E. Urriola, A. M. Hilbrands, L. J. Johnston, and G. C. Shurson. 2019. Growth performance of nursery pigs fed diets containing increasing levels of a novel high-pro-tein corn distillers dried grains with solubles. Transl. Anim. Sci. 3:38–47. doi: 10.1093/tas/txy101.
Zeng, Z. K., G. C. Shurson, and P. E. Urriola. 2017. Prediction of the concentration of standardized ileal digestible amino acids and safety margins among sources of distillers dried grains with solubles for growing pigs: A meta-analysis approach. Anim. Feed Sci. Technol. 231:150–159. doi: 10.1016/j.anifeedsci.2017.07.010.
Zhang, S., T. J. Knight, K. J. Stalder, R. N. Goodwin, S. M. Lonergan, and D. C. Beitz. 2009. Effects of breed, sex and halothane genotype on fatty acid composi-tion of triacylglycerols and phospholipids in pork lon-gissimus muscle. J. Anim. Breed. Genet. 126:259–268. doi:10.1111/j.1439-0388.2008.00782.x.








