Introduction
Increasing atmospheric concentrations of greenhouse gases (GHG) is a major worldwide concern. It is estimated that, globally, direct emissions from animal agriculture account for 7-10% of human derived GHG emissions (O’Mara, 2011), with this estimate increasing to 14.5% if indirect sources, such as feed production and deforestation for pasture or crop expansion are accounted for (Gerber et al., 2013). Beef and dairy production contribute directly to GHG emissions due to methane (CH4) generated during feed digestion (i.e., enteric CH4), and CH4 and nitrous oxide (N2O) from manure, as well as indirectly through N2O and CO2 emissions from crop production and fuel use, as well as carbon loss/gain in soil due to changes in land use. However, the largest single GHG source from ruminant production is enteric CH4. Methane is also a loss of potential energy from the animal with up to 12% of the total gross energy consumed by ruminants converted to CH4 and released via the breath. There has been a lot of interest in recent years in developing mitigation options to reduce GHG emissions from livestock production, and several excellent comprehensive reviews commissioned by the Food and Agricultural Organization (FAO) have recently been published (animal: Hristov et al., 2013a,b; manure: Montes et al., 2013). The present review summarizes the curent knowledge in the area of enteric CH4 reduction with a focus on the most promising dietary mitigation options available, or soon to be available, to livestock producers in Canada.
Sources of Emissions on the Farm
Methane accounts for 40–75% of total GHG emissions from beef and milk production in Canada when estimated using a “cradle-to-farm gate” life cycle analysis (Beauchemin et al., 2010; Mc Geough et al., 2012). This type of life cycle approach accounts for all CH4, N2O, and CO2 emissions arising from producing and transporting inputs used on the farm, livestock production on the farm, manure management, and growing and producing feed for the animals. Using a whole systems approach to evaluate a prospective mitigation practice ensures that a change in management aimed at reducing enteric CH4 production does not lead to an increase in emissions from other parts of the production cycle.
Methane arises primarily from enteric fermentation in the rumen and to a lesser extent, from manure storage. Life cycle assessment of confinement meat (beef) and milk (dairy) production systems indicate that more than 75% of the total CH4 is from enteric fermentation and less than 25% is from manure management (Beauchemin et al., 2010; Mc Geough et al., 2012). Emissions of CO2 from fossil fuel use are typically 5–10% of total GHG emissions. The remaining emissions (20–55% beef; 15–47% dairy) are N2O largely from denitrification of nitrogen in soils arising from manure and inorganic fertilizers (and urinary deposits on pasture), and to a lesser extent from sources of nitrogen resulting from leaching, runoff and volatilization.
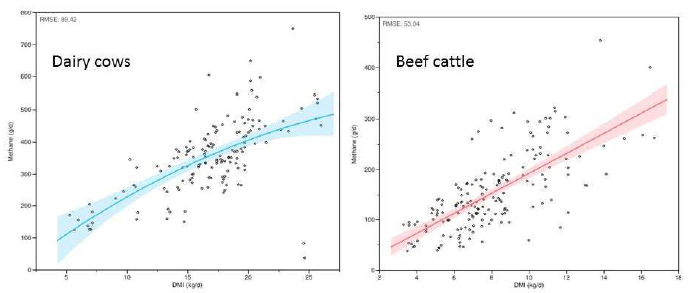
Enteric CH4 emission is proportional to the animal’s dry matter (DM) intake (DMI) (Figure 1), so expressing emissions on the basis of DMI, as g CH4/kg DMI or as a percentage of gross energy intake (GEI), accounts for differences in intake amongst animals. Emissions typically range from 200 to 500 g/d for lactating dairy cows and from 75 to 300 g/d for beef cattle, corresponding to 15 to 35 g CH4/kg DMI. The International Panel on Climate Change (IPCC, 2006) Tier 2 approach calculates CH4 emissions (55.6 MJ/kg) as 6.5 ± 1% of GEI except if diets contain >90% grain, in which case 3 ± 1% of GEI is lost as CH4.
Figure 1. A meta-analysis of the relationship between enteric methane emissions and dry matter intake (DMI) for dairy cows and beef cattle (each point is a treatment mean from the literature; Escobar, Beauchemin and Oba, unpublished data).
General Approaches to Lowering GHG Emissions from Livestock Production
The aim in reducing GHG emissions from livestock is to reduce the total CO2 equivalents (CO2e) per kilogram of milk or meat produced (referred to as GHG intensity or carbon footprint). Total CO2e are calculated by summing the individual GHGs after accounting for their respective global warming potential (100 year time horizon): CH4, kg × 25 + N2O, kg × 298 + CO2, kg (IPCC, 2007). In a global analysis of dairy production using a “cradle-to-farm gate” approach, Gerber et al. (2013) reported that GHG intensity (kg CO2e/kg fat and protein corrected milk [FPCM]) ranged from 1 to 2 for the industrialized regions of the world to a high of 7.5 for sub-Saharan Africa. The GHG intensity of milk production in Canada is at the very low end of this range at 0.8-1.2 kg CO2e/kg FPCM (Mc Geough et al., 2012; Vergé et al., 2013). Furthermore, GHG intensity of milk production tends to be lower in the western provinces (0.93 kg CO2e/L milk) than in the eastern provinces (1.12 kg of CO2e/L milk) due to climate conditions and management (Vergé et al., 2013).
Estimates of GHG emissions from beef production also vary widely depending upon production system, ranging from 17 to 37 kg CO2 eq/kg carcass. Globally, GHG intensities are lowest in Canada and the U.S. (17–23 kg CO2e/kg carcass), where beef production scenarios include a cow–calf suckling/grazing period followed by a growing phase and then by a high-grain finishing period in confined feedlots (Johnson et al., 2003; Beauchemin et al., 2010; Pelletier et al., 2010). Beef production systems that are extensively grass based or subject to climatic stress and feed shortages have much higher GHG intensity because of the slower growth of cattle (i.e, more days on feed, lower feed conversion efficiency) and higher CH4 emissions from fibrous, low quality feed.
As GHG intensity is a ratio between CO2 emissions and product output, intensity can be lowered through increased productivity and/or direct reduction of emissions. There are many examples globally whereby increased production has decreased GHG intensity. Capper et al. (2009) reported that the U.S. dairy industry reduced it carbon footprint between 1944 and 2007 by 63% due mainly to increased milk production per cow. Likewise, the Canadian dairy industry reduced its carbon footprint by 29% between 1981 and 2006, mainly due to an increase in milk production per cow and a reduction in animal numbers. Globally, there is tremendous potential to reduce the GHG intensity of meat and milk production through improved animal production. Increased animal productivity can be achieved through improvements in animal genetics, nutrition, health and overall management of the farm. The IPCC (2007) estimates that about 70% of the global GHG mitigation potential from agriculture lies in developing countries, where livestock production is mainly from smallholder farms with low-producing animals, and hence tremendous potential for increased productivity. While animal productivity is expected to continue to increase in North America, further decreases in GHG intensity on an industry-wide basis may be marginal as the limits of biological efficiency are reached. Increasing the productivity of lower producing animals has a relatively large impact on reducing GHG emission intensity, whereas a further increase in the productivity of highproducing animals has a relatively small impact. For example, increased milk production in Canada has occurred at the expense of cow longevity, adding to the need for additional replacement heifers that elevate GHG intensity of milk production because heifers consume feed, produce CH4, but do not contribute to milk production. We estimate in Canada that farms in the 10th percentile for lowest replacement rate (22%) have potentially a 6% lower GHG intensity of milk production compared with farms with average replacement rate (50th percentile, 37%) (Mc Geough, Little and Beauchemin, unpublished). Improving farm efficiencies, such as reproductive and feed efficiencies, are extremely important in lowering the carbon footprint of milk, especially in developed production systems. Because CH4 emissions are proportional to DMI, genetic selection of cattle for feed efficiency (e.g., residual feed intake) lowers emissions because feed requirements are reduced (Basarab et al., 2013). In highly productive livestock systems such as those in Canada, there is also opportunity to apply nutritional mitigation options that directly decrease methanogenesis in the rumen.
Decreased Methanogenesis through Nutrition
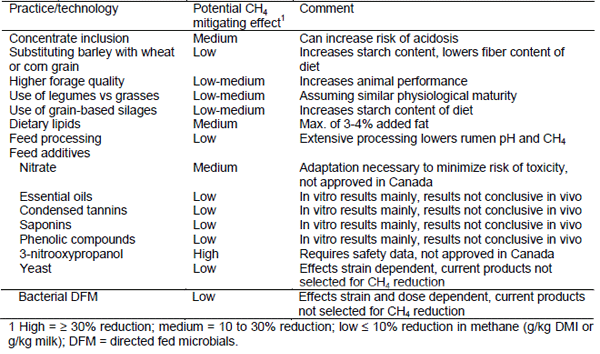
Enteric CH4 is a natural byproduct of microbial fermentation of feed in the rumen. Methane-producing Archaea (methanogens) prevent hydrogen from accumulating in the rumen by using it to reduce CO2 to CH4. The rumen microbial populations ferment feed to produce volatile fatty acids (VFA), including acetate, propionate and butyrate. Production of acetate and butyrate liberates metabolic hydrogen whereas propionate serves as a net hydrogen sink. Consequently, diets that increase propionate and decrease acetate in the rumen are often associated with a reduction in CH4 production. Furthermore, methanogens have a distinct association with protozoa, so defaunation of protozoa can also lower CH4 production. However, inhibiting methanogens without providing alternative electron acceptors can decrease feed digestibility and animal productivity. Development of strategies that reduce CH4 production without negative consequences on feed digestion and animal productivity is a significant challenge. Some dietary mitigation strategies that may reduce enteric CH4 production are listed in Table 1.
Table 1. Nutritional methane mitigation (CH4) practices and their expected effect (adapted from Gerber et al., 2013)
Grain content of diet
Feeding grain-based diets gives rise to less enteric CH4 (g/d and g/kg DMI) as compared to feeding forage-based diets because starch fermentation promotes propionate production in the rumen, which is a net hydrogen sink (Johnson and Johnson, 1995). Intensifying ruminant production by feeding more grain reduces enteric CH4 production, but whether net farm GHG emissions increase or decrease as a result of this practice depends on the particular farm. Furthermore, grain feeding ignores the importance of ruminants in converting fibrous feeds, unsuitable for human consumption, to high-quality protein sources (i.e., milk and meat).
Forage type and quality
Considerable variation in CH4 production occurs amongst forages. In most cases CH4 emissions (g/kg DMI) are lower from ruminants fed legumes than from those consuming grasses (McCaughey et al., 1999; Waghorn et al., 2002), because of the faster rate of passage from the rumen of legumes. However, plant maturity at the time of harvest can confound the impact of forage type on CH4 emissions. Advancing maturity causes reductions in soluble carbohydrate content and increased lignification of plant cell walls, which promote the production of acetate in the rumen, thereby increasing the amount of CH4 produced per unit of forage digested (Pinares-Patiño et al., 2003). Because reductions in forage quality are frequently accompanied by a reduction in ruminal digestibility, in many cases the amount of CH4 produced (g/kg DMI or as a percentage of GEI) is not influenced by forage quality (Pinares-Patiño et al., 2003; Molano et al., 2008). Nonetheless, improvements in forage quality usually lower emissions per kilogram of animal product as a result of enhanced animal productivity.
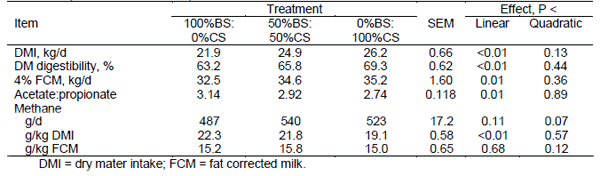
Cereal forages contain higher starch concentration than grass forages, which favours production of propionate over acetate and reduces CH4 production in the rumen. Corn silage and barley silage are the main cereal forages in eastern and western Canada, respectively, with starch concentrations typically higher in corn silage than barley silage. A study by Benchaar et al. (2014) showed that increasing corn silage proportion in the diet of dairy cows at the expense of barley silage increased dietary starch content (16.6 to 25.6% DM) and lowered NDF content (37.8 to 32.4% DM). Total tract digestibility of DM increased, as did DMI and fat corrected milk yield (Table 2). Although total CH4 production (g/d) increased due to higher DMI, CH4 g/kg DMI decreased, likely as a result of increased ruminal propionate proportion. Urinary N losses also decreased as the amount of corn silage included in the diet was increased at the expense of barley silage, reflecting enhanced efficiency of N use by the animal suggesting a low potential for N2O emissions. Thus, the increasing use of corn silage in beef and dairy production across Canada due to increasing availability of high yielding short season hybrids is expected to decrease GHG emissions from livestock.
Table 2. Effects of increasing corn silage (CS) proportion at the expense of barley silage (BS) in dairy cow diets (Benchaar et al., 2014).
Lipid supplementation of diets
Supplementation of diets with lipids is one of the most effective ways of lowering enteric CH4 emissions by ruminants, as reviewed by Grainger and Beauchemin (2011). The challenge is to lower CH4 emissions without impairing animal production, given that adding unprotected lipids to the diet can have negative effects on fiber digestion in the rumen. To prevent negative effects, the amount of lipid added to the diet must be limited to 2–4% of the dietary DM, such that total lipid content does not exceed 5-6% of dietary DM. The mechanism by which feeding lipids reduces CH4 production is multi-faceted. Replacing dietary carbohydrates with lipid decreases the amount of organic matter fermented in the rumen, lipids decrease protozoa numbers and associated methanogens, some fatty acids are toxic to rumen methanogens, and the biohydrogenation of fatty acids functions as a hydrogen sink.
When examined across studies with dairy cows, beef cattle and sheep, using different diets and experimental conditions, the reduction in CH4 (g/kg DMI) ranges from 2.5-6% per 1% dietary lipid, as summarized by Giger-Reverdin et al. (2003), Beauchemin et al. (2008), and Grainger and Beauchemin (2011). Furthermore, the reduction in CH4 is usually sustained with continued feeding of lipid. Thus, feeding lipids can be an effective CH4 mitigation practice but responses are variable depending upon the level of supplementation, fatty acid profile of the lipid source, form in which the lipid is administered (i.e., refined oil, full-fat oilseeds), and the type of diet (high forage diets are less responsive). Fat sources can be costly so improved animal performance is necessary to offset higher costs associated with feeding lipids for CH4 mitigation.
Plant bioactive compounds
A number of plant bioactive compounds, specifically tannins, saponins, and essential oils, have been explored for their ability to suppress CH4 emissions. Condensed tannins (CT) are secondary phenolic compounds in plants that vary widely in chemical structure, but have high affinity for protein. Tannins bind with proteins forming tannin-protein complexes, which reduce degradation of plant protein in the rumen, thereby enhancing the flow of feed protein to the intestines. For diets containing excess nitrogen, the protein-binding effect of CT can be beneficial for the environment as reduced nitrogen digestion reduces the excretion of nitrogen in urine, the form that is highly volatile.
However, reductions in CH4 emissions with CT appear to depend on the tannin source and ruminant species. Studies with sheep (Carulla et al., 2005; Hess et al., 2006) have shown decreased CH4 (g/kg DMI) (up to 20%) with supplemental CT, but our research in beef cattle with CT from quebracho trees (Beauchemin et al., 2007) and sainfoin (Chung et al., 2013) has found no reduction in CH4 with up to 2.7% dietary CT. It appears that the level of CT needed to reduce CH4 may be considerably higher than the level needed to decrease protein digestibility, and thus care must be taken to ensure that CH4 suppression using CT does not occur because of a reduction in feed digestion or feed intake. It is important that CH4 is not reduced at the expense of animal productivity, such as in the study conducted by Grainger et al. (2009) with dairy cows on pasture supplemented with grain. In that study, CH4 emission was reduced by up to 30%, but milk production of the cows was also reduced by about 10%.
Saponins are of interest for CH4 mitigation because they inhibit rumen ciliate protozoa. The two major commercial sources of saponins are the desert plants Yucca schidigera (steroidal saponins) from Mexico and Quillaja saponaria (triterpenoid saponins) from Chile. Many in vitro studies report a decrease in CH4 production when saponins are added, but the in vitro effects are rarely seen in vivo. Holtshausen et al. (2009) reported that feeding powdered Y. schidigera (containing 6% saponin) or Q. saponaria (3% saponin) to lactating dairy cows at 10 g/kg diet DM did not affect CH4 production, even though the same products reduced CH4 production in vitro. The differences between results in vitro and in vivo may indicate that the rumen microflora adapt to saponins. Clearly, additional animal feeding studies are necessary to establish the efficacy of saponins as a CH4 mitigation option.
Essential oils are the volatile aromatic compounds in plants that are used for their medicinal and antimicrobial properties. While many essential oils (e.g., garlic, cinnamon, oregano) have been shown in vitro to decrease CH4 production (as reviewed by Benchaar and Greathead, 2011), very few animal feeding studies have been conducted to substantiate these results. The favorable responses attained to date in vitro may not be as marked in vivo as a result of microbial adaptation. Furthermore, many of the concentrations of essential oils that elicit favorable responses in vitro are too high for in vivo applications, due to issues with palatability and possible toxicity. We found that feeding a commercial blend of essential oils to beef cattle had no effects on CH4 emissions (Beauchemin et al., 2006). However, recent studies with cashew nut shell liquid, the liquid obtained by pressing the shells of cashew nuts, show promise both in vitro and in vivo. The oil contains the alkenyl phenol anacardic acid, which has antibacterial activities against gram-positive bacteria. Cashew nut oil has been shown to inhibit CH4 production in vitro (Watanabe et al., 2010), but more importantly, in two small studies with three dairy cows, cashew nut decreased CH4 (g/kg DMI) by 19 to 38%. Methane reduction was accompanied by a decrease in the acetate to propionate ratio in rumen fluid (Shinkai et al., 2012). Larger scale and longer term studies are needed to substantiate these results.
Direct-fed microbials (DFM)
Commercial yeast products sold in Canada are from Saccharomyces cerevisiae and are generally categorized as active dry yeast which contains viable cells (>10 billion colony forming units per gram [cfu/g]) and yeast cultures that contain yeast plus culture. Many reviews have examined the beneficial effects of feeding yeast on milk production of dairy cows and weight gain of beef cattle (e.g., Beauchemin, 2012). Not unexpectedly, the results vary among studies, even when the same product is tested. However, when examined as a whole, there is an increasing body of literature that demonstrates their beneficial effects on animal productivity. However, their effects on methanogenesis are likely very small. We evaluated two commercially available strains of active dry yeast for beef cattle and reported no effects on CH4 production (McGinn et al., 2004). Subsequently, we fed a commercial strain of yeast and a new strain of yeast (selected for its ability to increase in vitro rate of fibre digestibility) to cattle and reported that enteric CH4 (g/kg DMI) was reduced by 10% using the new strain (Chung et al., 2011). The mechanism whereby yeast can decrease CH4 production is thought to be related to the increase in bacterial numbers that typically occurs as a result of added yeast. The partitioning of degraded carbohydrate between microbial cells and fermentation products may alter the production of hydrogen, thereby decreasing CH4 yield. While it may be possible to select yeasts that lower CH4 emissions, the commercial products currently available have not been selected for this purpose.
A wide variety of bacterial DFM are also registered for feeding to cattle in Canada. Those containing Propionibacterium strains are of interest because they may offer a means of increasing ruminal propionate production at the expense of CH4 production. As propionate is the major precursor for gluconeogenesis in ruminants, an increase in propionate production would be expected to increase feed conversion efficiency or animal production. We conducted a study (Vyas et al., 2014) in which beef heifers were assigned to a control treatment, or one of three Propionibacterium strains (P. acidipropionici P169; P. acidipropionici P5, and P. jensenii P54) dosed into the rumen once daily at 5 × 109 cfu. The heifers were fed a diet containing 70% forage. There were no treatment differences for total VFA, molar proportion of individual VFA, or CH4 production (g/d), although g CH4/kg DMI was reduced by 12, 8, and 13% with P169, P5, and P54 as compared to Control, respectively. The reduction in CH4 intensity (g CH4/kg of DMI) was a function of improved DMI with Propionibacterium strains, rather than a reduction in CH4. The relative abundance of Propionibacterium P169 in rumen fluid was greater 3 h post-dosing; however, the levels returned back to pre-treatment levels by 9 h post-treatment, suggesting lack of persistence of the supplemented bacterial strains. We concluded that these strains have a low capacity to survive and integrate into the ruminal microbial community. For bacterial additives to play a role in CH4 reduction, the challenge is to improve their survival and function within the rumen microbial community.
Alternative hydrogen sinks (nitrate)
Rumen fermentation converts carbohydrates into VFA and produces metabolic hydrogen. The main hydrogen sink in the rumen is methanogenesis; the methanogens reduce CO2 to CH4 which utilizes hydrogen (Janssen, 2012). However, there are also other pathways in the rumen that ensure that hydrogen does not accumulate; sulfates and nitrates are reduced to sulfides and ammonia, and unsaturated fatty acids are saturated. Synthesis of microbial cells also provides a sink for hydrogen.
When nitrate is present in the rumen, nitrite and ammonia formation are favoured over CH4 production (Ungerfeld and Kohn 2006). Thus, feeding nitrate can be a means of reducing CH4 production. Nitrate reduction has the added benefit of contributing to the rumen ammonia pool, which is often limiting in ruminants fed low quality diets. There is now overwhelming data to support the concept that feeding nitrate reduces CH4 emissions (Nolan et al., 2010; van Zijderveld et al., 2010, 2011; Hulshof et al., 2012; Li et al., 2012; El-Zaiat et al., 2014). In a dairy study, cows (33.2 kg/d of milk; 104 days in milk at the start) were fed a total mixed ration containing either urea or nitrate (21 g of nitrate/kg DM) during 4 successive 24-d periods (van Zijderveld et al., 2011). At the start of the study, the cows were adapted to the treatments over 4 weeks. Nitrate decreased CH4 production by 16% (g/d, g/kg DMI, % GEI), and the reduction persisted for the entire experiment. There were no negative effects on diet digestibility, but the energetic benefit from the decreased CH4 production did not appear to benefit the animal, as milk production and energy balance were not affected.
The major concern when feeding nitrate is the potential for toxicity if the nitrite level in the rumen exceeds the capacity of the microbes to convert it to ammonia. In that case, nitrate and nitrite are absorbed into the bloodstream and the nitrite causes toxicity by combining with hemoglobin to form methemoglobin. Thus, ferrous iron (Fe2+) in hemoglobin is transformed into ferric iron (Fe3+), rendering the hemoglobin molecule incapable of transporting oxygen to the tissues. If high levels of methemoglobin are formed (30– 40%), the animal begins to suffer from oxygen starvation, and must be treated immediately with methylene blue. The key is to adapt the animals to nitrate slowly so the population of nitrite-reducing bacteria increases in size and capacity to reduce nitrite. Another critical point is that the animals should not be restrictively fed. Feeding ad libitum decreases the rate of feed intake and helps ensure nitrate is converted to ammonia, without a build-up of nitrite in the rumen (Lee and Beauchemin, unpublished data). We are currently conducting studies to examine the potential benefits of using an encapsulated form of nitrate as a means of reducing its rate of availability in the rumen. With further evaluation and development of protocols to reduce toxicity risk, nitrate feeding could become a viable strategy to lower CH4 emissions. However, use of nitrate products for CH4 reduction would first need to be approved by Health Canada.
Inhibitors (3-nitrooxypropanol)
Some inhibitory compounds such as bromochloromethane, 2-bromo-ethane sulfonate, chloroform and cyclodextrin, have been shown to reduce CH4 emissions, but their safety as feed additives is a major limitation to their commercial use (McAllister and Newbold, 2008). Recently, the compound 3- nitrooxypropanol (NOP) from DSM Nutritional Products (Kaiseraugst, Switzerland), was shown to reduce CH4 emissions (Duval and Kindermann, 2012), but unlike other inhibitors, preliminary studies indicate that NOP has a high degree of safety although further study is needed. NOP is a methyl-coenzyme M analog, and thus is thought to inhibit the last step of methanogenesis.
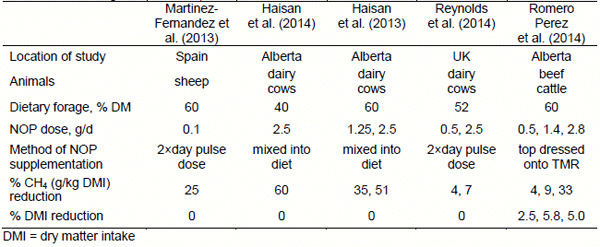
3-Nitrooxypropanol has been shown to reduce CH4 production (g/kg DMI) when supplemented to sheep (Martinez-Fernandez et al., 2013), lactating dairy cows (Haisan et al., 2013, 2014; Reynolds et al., 2014), and growing beef cattle (Romero Perez et al., 2014) with no signs of animal illness or intoxication (Table 3). Although NOP reduced CH4 production in all studies, the magnitude of the response was variable (from 4 to 60% reduction), depending upon animal type, diet, and in particular, method of dosing the product. Either no effect, or very minor effects, on DMI were reported. As those studies were short term (< 1 month feeding periods), we recently examined the effect of feeding NOP over a 4 month period in beef cattle with NOP included in the total mixed ration (Romero Perez et al., unpublished data). Methane production was consistently reduced by 60% with no signs of adaptation. We are currently conducting a larger scale feedlot trial to evaluate whether CH4 is lowered both during the backgrounding and finishing period, and to establish effects on animal performance and safety. The results to date for NOP have been very promising, but obviously, further research is required to fully establish its efficacy and safety before it can be registered for animal feeding in Canada.
Table 3. Summary of the effects of the methane inhibitor 3-nitrooxypropanol (NOP; DSM Nutritional Products, Kaiseraugst, Switzerland)
Conclusions
Interventions to reduce CH4 emissions from beef and dairy production currently available in Canada are limited to mainly those practices that improve production efficiency at the animal and herd levels. Improved diet formulation, herd health, and animal management can increase product output, which reduces enteric emissions relative to the quantity of meat/milk produced by the farm. These types of improvement in management can also increase farm profitability. It is also possible to apply mitigation strategies that directly reduce methanogenesis, such as the use of high starch forages and lipid supplementation of diets. In the future, it may be possible to use feed additives to reduce methanogenesis, as a number of very promising compounds have been identified. However, additional animal studies are needed to confirm these products are effective and safe. Although substantial research is required (especially long-term studies) to bring these feed additives forward, the potential for reducing the carbon footprint of meat and milk in Canada, and globally, is a tremendous incentive.
References
Basarab, J.A., K.A. Beauchemin, V.S, Baron, K.H. Ominski, L.L. Guan, S.P. Miller, and J.J. Crowley. 2013. Reducing GHG emissions through genetic improvement for feed efficiency: effects on economically important traits and enteric methane production. Animal 7:303–315.
Beauchemin, K.A. 2012. Use of yeast in ruminant diets. Pages 122-137 in the Proc. of the 33th Western Nutrition Conference, Winnipeg, Manitoba.
Beauchemin, K.A., and S.M. McGinn. 2006. Methane emissions from beef cattle: effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 84:1489–1496.
Beauchemin, K.A., S.M. McGinn, T.F. Martinez, and T.A. McAllister. 2007. Use of condensed tannin extract from quebracho trees to reduce methane emissions from cattle. J. Anim. Sci. 85:1990–1996.
Beauchemin, K.A, M. Kreuzer, F. O’Mara, and T.A. McAllister. 2008. Nutritional management for enteric methane abatement: a review. Australian J. Exp. Agric. 48:21–27.
Beauchemin, K.A., H.H. Janzen, S.M. Little, T.A. McAllister, and S.M. McGinn. 2010. Life cycle assessment of greenhouse gas emissions from beef production in western Canada: A case study. Agric. Syst. 103:371–379.
Benchaar, C., and H. Greathead. 2011. Essential oils and opportunities to mitigate enteric methane emissions from ruminants. Anim. Feed Sci. Technol. 166–167:338–355.
Benchaar, C., F. Hassanat, R. Gervais, P.Y. Chouinard, H.V. Petit, and D.I. Massé. 2014. Methane production, digestion, ruminal fermentation, nitrogen balance, and milk production of cows fed corn silage- or barley silage-based diets. J. Dairy Sci. 97:1–14.
Capper,J.L., R.A. Cady, and D.E. Bauman. 2009. The environmental impact of dairy production: 1944 compared with 2007. J. Anim. Sci. 87:2160-2167.
Carulla, J.E., M. Kreuzer, A. Machmuller, and H.D. Hess. 2005. Supplementation of Acacia mearnsii tannins decreases methanogenesis and urinary nitrogen in forage-fed sheep. Aust. J. Agric. Res. 56:961– 970.
Chung, Y.-H., N.D. Walker, S.M. McGinn, and K.A. Beauchemin. 2011. Differing effects of 2 active dried yeast (Saccharomyces cerevisiae) strains on ruminal acidosis and methane production in nonlactating dairy cows. J. Dairy Sci. 94:2431–2439.
Chung, Y.-H., E.J. Mc Geough, S. Acharya, T.A. McAllister, S.M. McGinn, O.M. Harstad, and K.A. Beauchemin. 2013. Enteric methane emission, diet digestibility, and nitrogen excretion from beef heifers fed sainfoin or alfalfa. J. Anim. Sci. 91:4861-4874.
Duval, S., and M. Kindermann. 2012. Use of nitooxy organic molecules in feed for reducing enteric methane emissions in ruminants, and/or to improve ruminant performance, International Patent Application WO 2012/084629 A1, World Intellectual Property Organization.
El-Zaiat, H.M., R.C. Araujo, Y.A. Soltan, A.S. Morsy, H. Louvandini, A.V. Pires, H.O., Patino, P.S. Correa, and A.L. Abdalla. 2014. Encapsulated nitrate and cashew nut shell liquid on blood and rumen constituents, methane emission, and growth performance of lambs. J. Anim. Sci. http://www.journalofanimalscience.org/content/early/2014/03/18/jas.2013-7084
Gerber, P.J., H. Steinfeld, B. Henderson, A. Mottet, C. Opio, J. Dijkman, A. Falcucci, and G. Tempio. 2013. Tackling climate change through livestock – A global assessment of emissions and mitigation opportunities. Food and Agriculture Organization of the United Nations (FAO), Rome.
Giger-Reverdin, S., P. Morand-Fehr, and G. Tran. 2003. Literature survey of the influence of dietary fat composition on methane production in dairy cattle. Livest. Prod. Sci. 82:73–79.
Grainger, C., and K.A. Beauchemin. 2011. Can enteric methane emissions from ruminants be lowered without lowering their production? Anim. Feed Sci. Technol. 166–167:308–320.
Grainger, C., T. Clarke, M.J. Auldist, K.A. Beauchemin, S.M. McGinn, G.C. Waghorn, and R.J. Eckard. 2009. Potential use of Acacia mearnsii condensed tannins to reduce methane emissions and nitrogen excretion from grazing dairy cows. Can. J. Anim. Sci. 89:241-251.
Haisan, J., Y. Sun, K. Beauchemin, L. Guan, S. Duval, D.R. Barreda, and M. Oba. 2013. Effects of feeding 3-nitrooxypropanol, at varying levels, on methane emissions and rumen fermentation in lactating dairy cows. Adv. Anim. Biosci. 4:326 (Abstr.).
Haisan, J., Y. Sun, L.L. Guan, K.A. Beauchemin, A. Iwaasa, S. Duval, D.R. Barreda, and M. Oba. 2014. The effects of feeding 3-nitrooxypropanol on methane emissions and productivity of Holstein cows in mid lactation. J. Dairy Sci. 97:1–10.
Hess, H.D., T.T. Tiemann, F. Noto, J.E. Carulla, and M. Kruezer. 2006.Strategic use of tannins as means to limit methane emission from ruminant livestock. In: Soliva, C.R., J. Takahashi, and M. Kreuzer. (Eds.) 2nd International Conference of Greenhouse Gases and Animal Agriculture. ETH Zurich, Zurich, Switzerland; p. 94–97.
Holtshausen, L., A.V. Chaves, K.A. Beauchemin, S.M. McGinn, T.A. McAllister, N.E. Odongo, P.R. Cheeke, and C. Benchaar. 2009. Feeding saponin-containing Yucca schidigera and Quillaja saponaria to decrease enteric methane production in dairy cows. J. Dairy Sci. 92:2809–2821.
Hristov, A.N., J. Oh, J. Firkins, J. Dijkstra, E. Kebreab, G. Waghorn, A. Adesogan, W. Yang, J. Tricarico, C. Lee, P.J. Gerber, B. Henderson, and H.P.S. Makkar. 2013a. Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 91:5045-5069.
Hristov, A.N., T. Ott, J. Tricarico, A. Rotz, G. Waghorn, A. Adesogan, J. Dijkstra, F. Montes, J. Oh, E. Kebreab, S.J. Oosting, P.J. Gerber, B. Henderson, H.P.S. Makkar and J. Firkins. 2013b. Mitigation of methane and nitrous oxide emissions from animal operations: III. A review of animal management mitigation options. J. Anim. Sci. 91:5095-5113.
Hulshof, R.B.A., A. Berndt, W.J.J. Gerrits, J. Dijkstra, S.M., van Zijderveld, J.R. Newbold, and H.B. Perdok. 2012. Dietary nitrate supplementation reduces methane emission in beef cattle fed sugarcane based diets. J. Anim. Sci. 90:2317-2323.
IPCC (Intergovernmental Panel on Climate Change). 2006. In: H.S. Eggleston, L. Buendia, K. Miwa, T. Ngara, and K. Tanabe, (Eds.), Guidelines for National Greenhouse Gas Inventories, Prepared by the National Greenhouse Gas Inventories Programme. IGES, Japan, www.ipccnggip.iges.or.jp/public/2006gl/index.htm.
IPCC (Intergovernmental Panel on Climate Change). 2007. Climate Change 2007: Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change Core Writing Team, R.K. Pachauri, and A. Reisinger (Eds.), IPCC, Geneva, Switzerland. p. 104.
Janssen, P.H. 2012. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim. Feed Sci. Technol. 160:1–22.
Johnson, K.A., and D.E. Johnson, 1995. Methane emissions from cattle. J. Anim. Sci. 73:2483–2492. Johnson, D.E., H.W. Phetteplace, A.F. Seidl, U.A. Schneider, and B.A. McCarl. 2003. Management variations for U.S. beef production systems: effects on greenhouse gas emissions and profitability. In: Proc. of the 3rd International Methane and Nitrous Oxide Mitigation Conference, November 17–21, Beijing, China. Coal Information Institute, Beijing, China, pp. 953–961.
Li, L., J. Davis, J. Nolan, and R. Hegarty. 2012. An initial investigation on rumen fermentation pattern and methane emission of sheep offered diets containing urea or nitrate as the nitrogen source. Anim. Prod. Sci. http://dx.doi.org/10.1071/AN11254
Martinez-Fernandez, G., A. Arco, L. Abecia, G. Cantalapiedra-Hijar, E. Molina-Alcaide, A.I. Martin-Garcia, M. Kindermann, S. Duval, and D.R. Yanez-Ruiz. 2013. The addition of ethyl-3-nitrooxy propionate and 3- nitrooxypropanol in the diet of sheep substantially reduces methane emissions and the effect persists over a month. Adv. Anim. Biosci. 4:368 (Abstr.).
McAllister, T.A., and C.J. Newbold. 2008. Redirecting rumen fermentation to reduce methanogenesis. Austr. J. Exp. Agric. 48:7–13.
McCaughey, W. P., K. Wittenberg, and D. Corrigan. 1999. Impact of pasture type on methane production by lactating beef cows. Can. J. Anim. Sci. 79:221–226.
Mc Geough, E.J., S.M. Little, H.H. Janzen, T.A., McAllister, S.M. McGinn, and K.A. Beauchemin. 2012. Life-cycle assessment of greenhouse gas emissions from dairy production in Eastern Canada: A case study. J. Dairy Sci. 95:5164-5175.
McGinn, S.M., K.A. Beauchemin, T. Coates, and D. Colombatto. 2004. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and furmaric acid. J. Anim. Sci. 82: 3346-3356.
Molano, G., and H. Clark, 2008. The effect of intake and forage quality on methane production by sheep. Austr. J. Exp. Agric. 48:219–222.
Montes, F., R. Meinen, C. Dell, A. Rotz, A.N. Hristov, J. Oh, G. Waghorn, P.J. Gerber, B. Henderson, and H.P.S. Makkar. 2013. Mitigation of methane and nitrous oxide emissions from animal operations: II. A review of manure management options. J. Anim. Sci. 91:5070-5094.
Nolan, J.V., R.S. Hegarty, J. Hegarty, I.R. Godwin, and R. Woodgate. 2010. Effects of dietary nitrate on fermentation, methane production and digesta kinetics in sheep. Anim. Prod. Sci. 50:801–806.
O’Mara, F.P. 2011. The significance of livestock as a contributor to global greenhouse gas emissions today and in the near future. Anim. Feed Sci. Technol. 166-167:7-15.
Pelletier, N., R. Pirog, R. Rasmussen. 2010. Comparative life cycle environmental impacts of three beef production strategies in the Upper Midwestern United States. Agric. Syst. 103:380–389.
Pinares-Patiño, C.S., R. Baumont, and C. Martin. 2003. Methane emissions by Charolais cows grazing a monospecific pasture of timothy at four stages of maturity. Can. J. Anim. Sci. 83:769–777.
Reynolds, C.K., D.J. Humphries, P. Kirton, M. Kindermann, S. Duval, and W. Steinberg. 2014. Effects of 3-nitrooxypropanol on methane emission, digestion, and energy and nitrogen balance of lactating dairy cows. J. Dairy Sci. 97:1-13.
Romero Perez, A., K.A. Beauchemin, S.M. McGinn, E.K. Okine, L.L. Guan, M. Oba and S.M. Duval. 2014. The potential of 3-nitrooxypropanol to lower enteric methane emissions from beef cattle. J. Anim. Sci. (submitted)
Shinkai, T., O. Enishi, M. Mitsumori, K. Higuchi, Y. Kobayashi, A. Takenaka, K. Nagashima, M. Mochizuki, and Y. Kobayashi. 2012. Mitigation of methane production from cattle by feeding cashew nut shell liquid. J. Dairy Sci. 95:5308–5316.
Ungerfeld, E.M., and R.A. Kohn. 2006. Pages 55-85 In: Sejrsen, K., T. Hvelplund, and M.O. Nielson (Eds.) Ruminant physiology. Digestion, metabolism and impact of nutrition on gene expression, immunology and stress. Wageningen Academic Publishers, Wageningen, The Netherlands.
van Zijderveld, S.M., W.J.J. Gerrits, J.A. Apajalahti, J.R. Newbold, J. Dijkstra, R.A. Leng, and H.B. Perdok. 2010. Nitrate and sulfate: Effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep J. Dairy Sci. 93:5856–5866.
van Zijderveld, S.M., W.J.J. Gerrits, J. Dijkstra, J.R. Newbold, R.B.A. Hulshof, and H.B. Perdok. 2011. Persistency of methane mitigation by dietary nitrate supplementation in dairy cows. J. Dairy Sci. 94:4028– 4038.
Vergé, X.P.C., D. Maxime, J.A. Dyer, R.L. Desjardins, Y. Arcand, and A. Vanderzaag. 2013. Carbon footprint of Canadian dairy products: Calculations and issues. J. Dairy Sci. 96 :6091–6104.
Vyas D., E.J. McGeough, S.M. McGinn, T.A. McAllister, and K.A. Beauchemin. 2014. Effect of Propionibacterium spp. on ruminal fermentation, nutrient digestibility, and methane emissions in beef heifers fed a high forage diet. J. Anim. Sci. jas.2013-7492; published ahead of print March 18, 2014, doi:10.2527/jas.2013-7492.
Waghorn, G.C., M.H. Tavendale, and D.R. Woodfield. 2002. Methanogenesis from forages fed to sheep. Proc. N.Z. Grassland Assoc. 64:159–165.
Watanabe, Y., R. Suzuki, S. Koike, K. Nagashima, M. Mochizuki, R.J. Forster, and Y. Kobayashi. 2010. In vitro evaluation of cashew nut shell liquid as a methane-inhibiting and propionate-enhancing agent for ruminants. J. Dairy Sci. 93:5258–5267.