The effect of dietary inclusions of guanidinoacetic acid on D1-42 broiler performance and processing yields
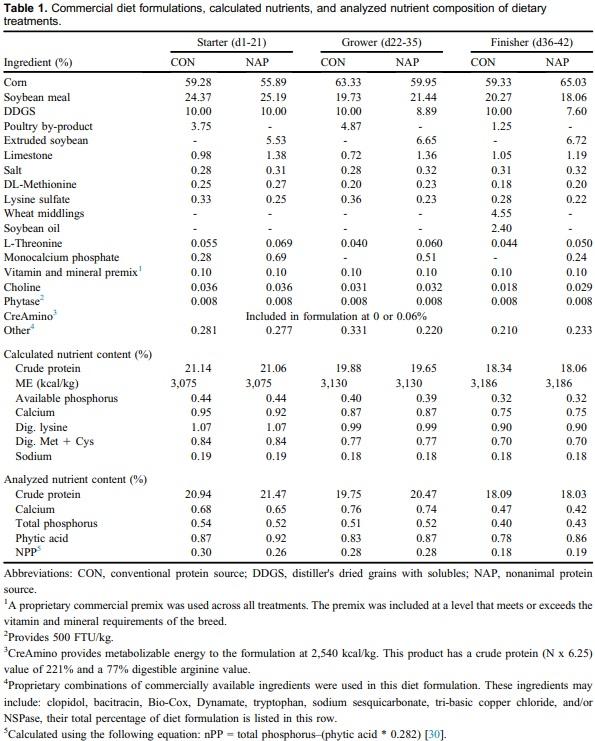
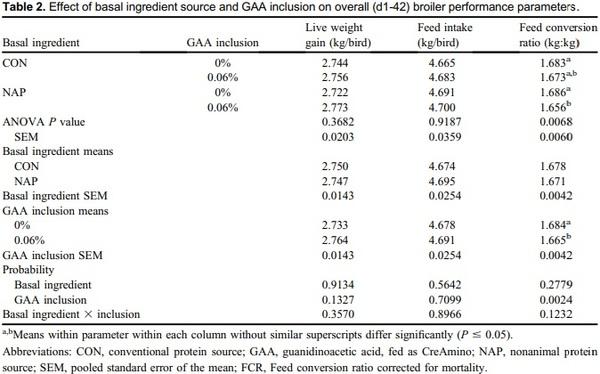
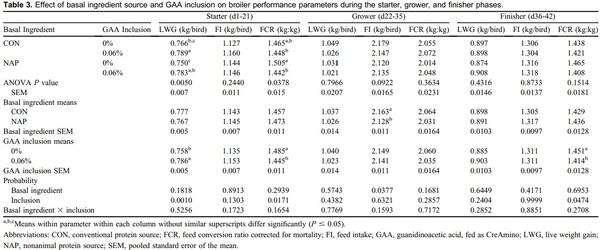
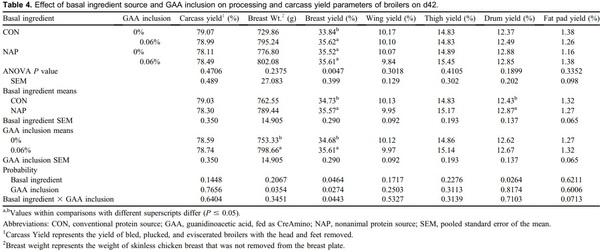
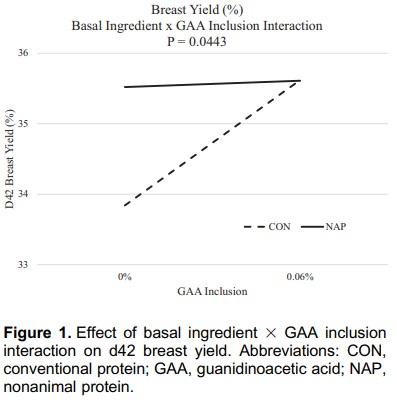
Creatine is a central constituent in energy metabolism, especially in cells with variable energy demand, such as muscle cells. Animal proteins such as fish meal, poultry meal, and meat meal may provide adequate amounts of creatine when included in diet formulation. Guanidinoacetic acid (GAA) is an immediate precursor of creatine and should be considered for use in animal nutrition. An experiment was conducted to describe broiler performance and carcass yield effects when GAA was provided to broilers consuming diets containing either animal protein or nonanimal protein. Treatments were arranged in a 2 x 2 factorial varying in basal ingredients (conventional [CON] or nonanimal protein [NAP]) and GAA inclusion (0 or 0.06%), provided as CreAmino, in a randomized complete block design. Treatments were applied to 12 replicate pens of 36 Hubbard x Cobb 500 straight-run broilers from hatch through d 42. Overall (d1-42) results indicate 0.019 FCR improvement when GAA was included to broiler diets (P = 0.0024). Similar results were apparent in each feed phase period (P < 0.05). Processing yield results indicate a 45-g increase in breast weight when GAA was included in broiler diets (P = 0.0354). Basal ingredients and GAA inclusion effects interacted, which affected d42 breast yield (BY) (P = 0.0443). The inclusion of GAA in NAP diets did not affect BY; however, BY was reduced by 1.78 percentage points when broilers were provided the CON diet devoid of GAA. GAA has the potential to improve broiler performance and BY when included in broiler diets with basal ingredients including conventional animal protein.
Key words: energy metabolism, all vegetable, breast yield.
1. Michiels, J., L. Maertens, J. Buyse, A. Lemme, M.
Rademacher, N. A. Dierick, and S. De Smet. 2012. Supplementation of guanidinoacetic acid to broiler diets: effects on performance, carcass characteristics, meat quality, and energy metabolism. Poult. Sci. 91:402–412.
2. Brosnan, J. T., E. P. Wijekoon, L. Warford-Woolgar,
N. L. Trottier, M. E. Brosnan, J. A. Brunton, and R. F. P.
Bertolo. 2009. Creatine synthesis is a major metabolic process in Neonatal Piglets and has important Implications for amino acid metabolism and methyl Balance. J. Nut. 139:1292–1297.
3. Wyss, M., and R. Kaddurah-Daouk. 2000. Creatine and creatinine metabolism. Phys. Rev. 80:1107–1213.
4. Walker, J. B. 1979. Creatine: biosynthesis, regulation, and function. Adv. Enzymol. Relat. Areas Mol. Biol.
50:177–242.
5. Lemme, A., J. Ringel, H. S. Rostagno, and M. S.
Redshaw. 2007. Supplemental guanidino acetic acid improved feed conversion, weight gain, and breast meat yield in male and female broilers. In: Proc. 16th Eur. Symp.
Poult. Nutr., Strasbourg, France. World’s Poult. Sci. Assoc.,
Beekbergen, the Netherlands, pp. 335–338.
6. Vieira, S. L., and I. L. Lima. 2005. Live performance, water intake and excreta characteristics of broilers fed all vegetable diets based on corn and soybean meal. Int. J.
Poult. Sci. 4:365–368.
7. Jayathilakan, K., K. Sultana, K. Radhakrishna, and
A. S. Bawa. 2011. Utilization of byproducts and waste materials from meat, poultry and fish processing industries: a review. J. Food Sci. Technol. 49:278–293.
8. Dobenecker, B., and U. Braun. 2015. Creatine and creatinine contents in different diet types for dogs – effects of source and processing. J. Anim. Physiol. Anim. Nutr.
99:1017–1024.
9. Khan, A. W., and D. C. Cowen. 1977. Rapid estimation of muscle proteins in beef-vegetable protein mixtures. J. Agric. Food Chem. 25:236–238.
10. Gabor, E., O. Gaspar, and E. Vamos. 1984. Quantitative determination of muscle protein in meat products by measuring creatine content. Acta Alimentaria. 13:13–22.
11. Baker, D. H. 2009. Advances in protein–amino acid nutrition of poultry. Amino Acids. 37:29–41.
12. AlzChem Group AG, Trostberg, Germany.
13. Córdova-Noboa, H. A., E. O. Oviedo-Rondón,
A. H. Sarsour, J. Barnes, P. Ferzola, M. RademacherHeilshorn, and U. Braun. 2018. Performance, meat quality, and pectoral myopathies of broilers fed either corn or sorghum based diets supplemented with guanidinoacetic acid. Poult. Sci. 97:2479–2493.
14. Ringel, J., A. Lemme, A. Knox, J. Mc Nab, and M.
S. Redshaw. 2007. Effects of graded levels of creatine and guanidinoacetic acid in vegetable-based diets on performance and biochemical parameters in muscle tissue. In:
Proc. 16th Eur. Symp. Poult. Nut., Strasbourg, France, pp. 387–390.
15. Longenecker’s Hatchery, Elizabethtown, PA.
16. SAS Institute. 2017. The SAS System for Windows
2017. Release 9.4. SAS Inst. Inc., Cary, NC.
17. Mousavi, S. N., A. Afsar, and H. Lotfollahian. 2013.
Effects of guanidinoacetic acid supplementation to broiler diets with varying energy contents. J. Appl. Poult. Res.
22:47–54.
18. Córdova-Noboa, H. A., E. O. Oviedo-Rondón,
A. H. Sarsour, J. Barnes, D. Sapcota, D. López, L. Gross,
M. Rademacher-Heilshorn, and U. Braun. 2018. Effect of guanidinoacetic acid supplementation on live performance, meat quality, pectoral myopathies and blood parameters of male broilers fed corn-based diets with or without poultry by-products. Poult. Sci. Vol 97:2494–2505.
19. Tossenberger, J., M. Rademacher, K. Németh, V.
Halas, and A. Lemme. 2016. Digestibility and metabolism of dietary guanidino acetic acid fed to broilers. Poult. Sci.
95:2058–2067.
20. Lemme, A., J. Ringel, A.-R. Sterk, and J. F. Young.
2007. Supplemental guanidino acetic acid affects energy metabolism of broilers. In: Proc. 16th Eur. Symp. On Poult.
Nut., Strasbourg, France, pp. 339–342.
21. Dilger, R. N., K. Bryant-Angeloni, R. L. Payne, A.
Lemme, and C. M. Parsons. 2013. Dietary guanidino acetic acid is an efficious replacement for arginine in young chicks.
Poult. Sci. 92:171–177.
22. DeGroot, A. A., U. Braun, and R. N. Dilger. 2019.
Guanidinoacetic acid is efficacious in improving growth performance and muscle energy homeostasis in broiler chicks fed arginine-deficient or arginine-adequate diets.
Poult. Sci. 0:1–10.
23. Chamruspollert, M., G. M. Pesti, and R. I. Bakalli.
2002. The influence of labile dietary methyl donors on the arginine requirement of young broiler chicks. Poult. Sci.
81:1142–1148.
24. Lemme, A., R. Gobbi, and E. Esteve-Garcia. 2010b.
Effectiveness of Creatine Sources on Performance of
Broilers at Deficient or Adequate Methionine Supply. 13th
Eur. Poult. Con., Tours, France.
25. Ziegenfuss, T. N., L. M. Lowery, and P. W. R.
Lemon. 1998. Acute fluid changes in men during three days of creatine supplementation. Online J. Exerc.
Physiol. Vol. 3.
26. Häussinger, D. 1996. The role of cellular hydration in the regulation of cell function. Biochem. J.
313:697–710.
27. Prevost, M. C., A. G. Nelson, and G. S.
Morris. 1997. Creatine supplementation enhances intermittent work performance. Res. Q. Exerc. Sport.
68:233–240.
28. von Lengerken, G., S. Maak, and M. Wicke. 2002.
Muscle metabolism and meat quality of pigs and poultry.
Veterinarija Ir Zootechnika. 20:82–86.
29. Stahl, C. A., M. W. Greenwood, and E. P. Berg.
2003. Growth parameters and carcass quality of broilers fed a corn-soybean diet supplemented with creatine monohydrate. Int. J. Poult. Sci. 2:404–408.
30. Angel, R., N. M. Tamim, T. J. Applegate, A. S.
Dhandu, and L. E. Ellestad. 2002. Phytic acid chemistry: influence on phytin-phosphorus availability and phytase efficacy. J. Appl. Poult. Res. 11:471–480.












")
