Optimizing Calcium, Phosphorus and Phytase Formulation in layer Diets
Published: November 4, 2014
By: Stuart Wilkinson, Aaron Cowieson and Emma Bradbury (Poultry Research Foundation, University of Sydney)
BACKGROUND
Calcium (Ca) is an essential nutrient for poultry that is generally provided to laying hens at approximately 4.6% of the total feed volume. Calcium is usually supplied to layers as a calcite grit or flour i.e. limestone (CaCO3) and also as part of the inorganic phosphorus (P) supplye.g. dicalcium or monocalcium phosphate. Though, in some markets, meat and bone meal may also contribute substantially to this. Only a minor part of the birds’ Ca requirement is met by vegetable ingredients such as wheat, canola or soyabean meal. Though Ca is an essential dietary requirement, high concentrations of Ca, particularly from limestone in poultry diets, results in a dramatic reduction in the digestibility of the diet. Paradoxically, poultry diets are formulated on a total, rather than a digestible Ca basis. This has led to some confusion regarding the birds’ requirement for Ca and most probably suggests that current recommendations may be in excess to the requirements of layers. Excess Ca can lead to poor phosphorus (P) digestibility and contribute to the formation of insoluble mineral-phytate complexes. Therefore utilisation of phytate-P can be improved by feeding Ca strictly to minimum requirements (Underwood and Suttle, 2001a). This paper presents an update on the findings of an ongoing project investigating the calcium and phosphorus requirements of layers that is funded by the Australian Egg Corporation Ltd.
CALCIUM AND PHOSPHORUS IN POULTRY NUTRITION
Much has been written on this subject so only a brief description will be provided herein. Calcium is the most prevalent mineral in the body with approximately 99% found in the skeleton (Underwood and Suttle, 2001a). Hence, the predominant role of Ca is to provide structural strength and support in bones. However, Ca is also critical to many biochemical pathways such as muscle and nerve conduction, blood coagulation, control of hormone secretions such as Vitamin D3 and parathyroid hormone and egg shell calcification (de Matos, 2008). Deficiencies of Ca lead to the development of skeletal abnormalities and reduced egg shell quality.
Phosphorus is the second most abundant mineral in the body with 80% found in bones. Importantly, P is a critical component of phospholipids which contribute to cell membrane fluidity and integrity as well as the phosphate buffer system which helps maintain osmotic and acid-base balance (Underwood and Suttle, 2001b). Deficiencies of P in poultry lead to similar outcomes as for Ca deficiency including reduced skeletal integrity, appetite, subnormal growth in young animals and weight loss in older animals (Christensen et al., 2003; Narcy et al., 2009).
PHYTAT
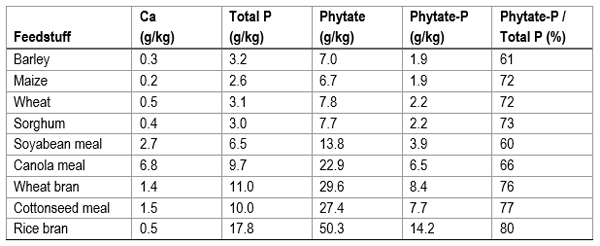
Phytate (myo-inositol hexaphosphate: IP-6) serves as the storage form of P in plants which is predominantly stored in seeds. This has implications for pig and poultry nutrition since diets fed to both species use seeds extensively. Phytate-P is considered to have a low bioavailability to monogastric animals, at least when fed in mixed rations (more on this later). Other points to consider are that feedstuffs vary in their concentration of phytate-P as well as in the proportion of total P that is present as phytate-P (Table 1). The concentration of phytate-P varies from 0.2% in maize to approximately 1.4% in rice bran whilst the proportion of P as phytate-P ranges from 60% in soybean meal to 80 in rice bran. Therefore dietary ingredients greatly influence the amount of phytate, phytate-P and non-phytate in the diet. To overcome this, diets include inorganic sources of P to meet the nutritional requirements of the bird.
Table 1 Concentrations of calcium (Ca), total phosphorus (P), phytate-phosphorus and the proportion of phytate-phosphorus in common feedstuffs (Selle et a l., 2009)
INTERACTIONS OF CALCIUM, PHOSPHORUS AND PHYTATE
Conventionally it is believed that phytate-P is not able to be hydrolysed by poultry despite mucosal phytases being present in the small intestines of poultry. However, their efficacy is thought to be limited by the high concentration of Ca in poultry diets, which renders phytate insoluble at the pH of the small intestine (Tamim et al., 2004). The relationship between dietary Ca and phytate-P was proposed as early as the 1940’s and in later work by Taylor (1965) who demonstrated when dietary Ca is kept to a minimum, phytate-P utilisation is significantly improved. Similarly, work from the University of Maryland has shown that the addition of 0.5% Ca to a broiler diet without phytase, reduces the birds’ endogenous capacity to hydrolyse phytate-P from around 70 % to 25 % (Tamim et al., 2004). This is interpreted as a calcium-led reduction in the solubility of phytate-P via chelation and precipitation as a calcium 'phytin' salt.
Limestone also has a significant acid binding capacity and may prevent the young chick, pullet or adult bird from satisfactorily acidifying the diet in the gastric gut, compromising protein digestibility. It is important to note that the improved digestibility of phytate-P and other nutrients in poultry diets is thought to be the result of the physical separation of Ca and phytate and not simply that of feeding reduced amounts of Ca. Thus, though Ca is necessary for optimal performance and bone strength its addition (notably as limestone) is contrary to minimal FCR and maximal egg production, persistency and reduced fatigue. This 'calcium conundrum' is the genesis of the current project as the separate supply of calcium as a choice-fed grit may allow appropriate calcium intake (for skeletal health and shell quality) whilst mitigating risk associated with poor P and amino acid digestibility. Further, as poultry have a Ca specific appetite, the use of a choice-feeding model will allow both the assessment of its usefulness viz. P digestibility and performance effects (per se) as well as the establishment of more accurate Ca requirement figures, especially in concert with varying digestible P and phytase regimes.
Previous studies have confirmed that poultry are able to select their own diet from various feedstuffs in order to meet their nutritional requirements (Malheiros et al., 2003). There is evidence that this ‘nutritional wisdom’ applies for several essential nutrients including lysine (Newman and Sands, 1983), methionine (Steinruck et al., 1990), total protein (Forbes and Shariatmadari, 1994; Delezie et al., 2009), selenium (Zuberbuehler et al., 2002) and Ca (Wood-Gush and Kare, 1966; Hughes and Wood-Gush, 1972; Joshua and Mueller, 1979). Work in laying hens has shown that egg weight, shell
thickness and FCR were all significantly improved in choice-fed hens compared to hens that were fed a conventional mash diet (Olver and Malan, 2000). A review by Henuk and Dingle (2002) reported that the practice of choice feeding was historically a by-product of the poultry management and infrastructure available in the early to mid-20th century. However, as poultry production moved from small holder farms to more intensive and mechanised systems, feed delivery systems changed from being manual operated to automated feeding systems capable of delivering complete diets. One consequence of this process has been to remove the ability of poultry to choice feed.
WORK IN BROILERS
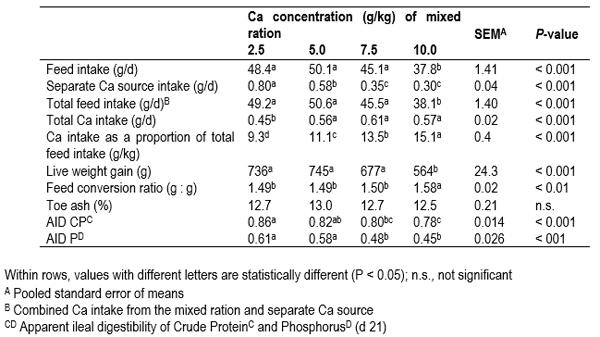
Work recently conducted at the Poultry Research Foundation in broilers investigated feeding broilers diets with varying Ca concentrations (2.5, 5.0, 7.5 and 10.0 g/kg) and a separate Ca source (CaCO3). The results from this study showed that modern broilers consumed significantly more of the separate Ca source as dietary Ca decreased (Table 2) and confirmed that the Ca specific appetite is still active in modern broilers. Similar to previous findings, P digestibility decreased with higher dietary Ca concentrations. Interestingly, feeding birds reduced Ca concentrations while providing a separate Ca source had no adverse effect on bird performance or bone mineralisation (toe ash). The significant improvement in CP and P digestibilities in birds fed reduced Ca diets are in keeping with earlier work. However, unlike previous studies this improvement was associated with consuming Ca separately from the mixed ration. These findings indicate that broilers were able to meet their Ca requirement when fed low Ca diets and a separate Ca source. The improvements in nutrient digestibility may be a result of feeding broilers low Ca diets as well as physically and temporally separating the consumption of phytate and Ca.
Table 2 The effect of dietary calcium (Ca) on the intake of a separate Ca source, total Ca intake broiler performance (d 1-21) and the apparent ileal digestibility (AID) of crude protein (CP) and phosphorus (P). Adapted from Wilkinson et al. (2014).
WORK IN LAYERS
Layer hens have been found to possess a significant Ca specific appetite. Presumably this originates from the ancestor of the modern bird, as historically they would not have consumed a nutritionally complete diet in one bolus (as modern layer hens do) but would rather have foraged, consuming seeds, roots, insects and soils/silicates to its nutritional satisfaction. Thus, the intake of phytate-P (in seeds) would have been quite separate from its intake of calcium (as soils, clays and silicates). This may explain why poultry are able to digest phytate-P in a low Ca environment as most of the P requirement of poultry may actually be met by the consumption of phytate-P. The poor (< 20%) retention of phytate-P by modern layers may be an artefact of the concerted provision of Ca as limestone or dicalcium phosphate with the vegetable component of the diet.
As a first step to investigate the optimisation Ca and P in layer diets, a study at the Poultry Research Foundation investigated whether modern layers possess the Ca-specific appetite as well as if this may be used to feed Ca separately to the mixed ration to improve nutrient digestibility. A total of 80 Isa Brown laying hens (approximately 35-42 weeks of age) were randomly allocated to one of four dietary treatments differing in Ca concentrations (10, 20, 30 or 40 g/kg Ca.) Diets were wheat-soyabean meal based and formulated to an AME of 11.72 MJ/kg. All birds were able to access a separate Ca source, approximately 95% CaCO3 with an average particle size of 1.4 mm (AB Grit, Omya Australia). Birds had ad libitum access to feed, water and the separate Ca source throughout the study. Birds were housed in individual cages (23cm W x 45cm D x 45cm H) in an environmentally controlled shed and were exposed to a lighting regime of 16h:8h (light:dark). Birds were acclimatised to their cage two weeks before the study commenced and fed the 40 g/kg Ca diet to minimise the effects of a change in diet during the study.
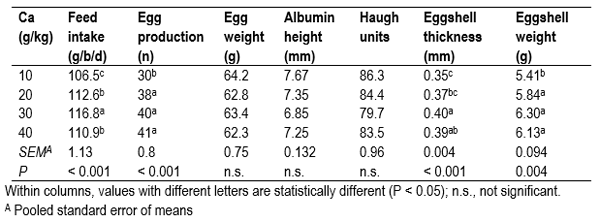
Data for feed intake and egg production are presented in Table 3. There was no difference between groups for egg weight, albumin height or Haugh units. Egg production increased with increasing dietary Ca and birds fed 10 g/kg Ca diets laid significantly less eggs than birds fed the other diets. Eggshells from birds fed 10 g/kg Ca diets were significantly thinner than those from birds fed diets containing 30 and 40 g/kg Ca. Dry egg shell weight was lowest in birds fed diets containing 10 g/kg Ca when compared to the other groups (P < 0.05). Calcium deficiency decreases egg production and reduces the thickness of the eggshell (Scott et al., 1971) and this would explain the results for birds fed the 10 g/kg Ca diet. In fact towards the end of the study, several birds in this study fed the 10 g/kg Ca diet ceased egg production and one bird was observed to regularly consume the egg shell, presumably in an attempt to obtain Ca.
Table 3 Effect of dietary calcium (Ca) concentration (g/kg) on feed intake and egg production in Isa Brown hens. Adapted from Bradbury et al. (2014)
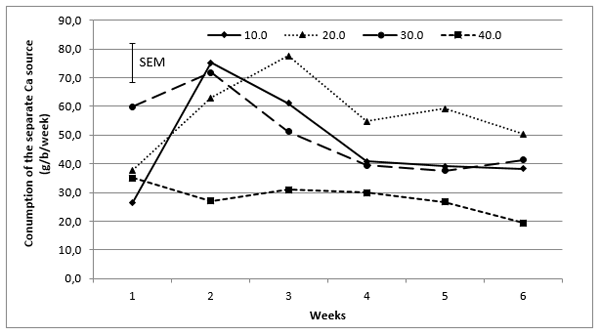
Overall, during the experimental period, birds consumed more of the separate Ca source (Figure 1) as dietary Ca decreased (P < 0.05). However, the results of the present study showed there were differences between individual birds in their ability to adjust to the current feeding regime. Not all birds were able to adapt to being fed low Ca diets and a separate Ca source. Some birds fed low Ca diets were unable to adapt to separate Ca choice feeding and this was particularly apparent in the group fed 10 g/kg Ca diets. This suggests that feeding layers 10 g/kg Ca (and a separate Ca source) was too low in the context of this study. This is in agreement with Gleaves and Salim (1982) who reported some hens did not consume a Ca supplement when fed 5.0 g/kg Ca diets. In the current study, birds fed 20 g/kg Ca diets showed a better response to the separate feeding of Ca, however, birds may require additional time beyond the acclimation period provided in this study. It appears that most birds within the groups fed 10 - 30 g/kg Ca diets identified that the diets offered were lower in Ca than required and consumed the separate Ca source to compensate for this deficiency. The pattern of consumption of the separate Ca source indicates birds fed the 10 g/kg diet increased their consumption of the separate Ca source in week 2 and this may indicate that birds were still adjusting to the experimental feed. Generally speaking, if the 10 g/kg dietary Ca group is excluded from the results from week three onwards, the consumption of the separate Ca source follows the assumption that birds on a low Ca diet will increase their separate Ca intake accordingly.
Figure 1 The effect of dietary calcium (Ca) concentration on the consumption (g/b/week) of a separate Ca source by Isa Brown hens.
It can be concluded that the Ca-specific appetite is not universally or uniformly expressed in laying hens. Though some birds were able to regulate to a common Ca intake and maintain production when fed reduced dietary Ca, others were unable to adapt to this new feeding regime. Further work is required to explore the optimum time, ontogenetically, to introduce a separate Ca source and to consider groups of birds where birds with reduced ability to self-regulate may learn from the early adopters. The effect of separating Ca from phytate on nutrient digestibility in layers is currently being assessed. The effects of phytase will also be explored in future studies. With a better understanding, it may be possible to spatially separate Ca from the basal diet and exploit the Ca-specific appetite of layer hens to specifically improve the digestibility of phytate-P and amino acids. This would enhance the profitability and sustainability of the egg industry. This work is ongoing with the support of the Australian Egg Corporation Limited.
REFERENCES
Bradbury, E.J., Wilkinson, S.J., Cronin, G.M., Bedford, M.R. and Cowieson, A.J. (2014). Exploring the calcium appetite of layers with choice feeding. Proceedings of the 25th Annual Australian Poultry Science Symposium, Sydney, New South Wales, Australia, 16-19 February 2014,
Christensen, J.W., Nielsen, B.L., Young, J.F. and Nøddegaard, F. (2003). Effects of calcium deficiency in broilers on the use of outdoor areas, foraging activity and production parameters. Applied Animal Behaviour Science, 82, 229-240
De Matos, R. (2008). Calcium metabolism in birds. Veterinary Clinics of North America Exotic Animal Practice, 11, 59-82
Delezie, E., Maertens, L., Huyghebaert, G. and Lippens, M. (2009). Can choice feeding improve performances and n-retention of broilers compared to a standard three-phase feeding schedule? British Poultry Science, 50, 573-582
Forbes, J.M. and Shariatmadari, F. (1994). Diet selection for protein by poultry. Worlds Poultry Science Journal, 50, 7-24
Gleaves, E.W. and Salim, A.A. (1982). The effect of lactose supplementation and source on feed intake and production characteristics of laying hens. Poultry Science, 61, 2390-2397
Henuk, Y.L. and Dingle, J.G. (2002). Practical and economic advantages of choice feeding systems for laying poultry. Worlds Poultry Science Journal, 58, 199-208
Hughes, B.O. and Wood-Gush, D.G.M. (1972). Hypothetical mechanisms underlying calcium appetite in fowls. Revue du Comportement Animal, 6, 95-106
Joshua, I.G. and Mueller, W.J. (1979). The development of a specific appetite for calcium in growing broiler chicks. British Poultry Science, 20, 481-490
Malheiros, R.D., Moraes, V.M.B., Collin, A., Decuypere, E. and Buyse, J. (2003). Free diet selection by broilers as influenced by dietary macronutrient ratio and corticosterone supplementation. 1. Diet selection, organ weights, and plasma metabolites. Poultry Science, 82, 123-131
Narcy, A., Letourneau-Montminy, M.P., Magnin, M., Lescoat, P., Jondreville, C., Sauvant, D. and Nys, Y. 2009. Phosphorus utilisation in broilers. World Poultry Science Association (WPSA), 17th European Symposium on Poultry Nutrition, 23-27 August, 2009. 2009 Edinburgh, UK. Beekbergen Netherlands: World Poultry Science Association (WPSA), 14-20
Newman, R.K. and Sands, D.C. (1983). Dietary selection for lysine by the chick. Physiology & Behavior, 31, 13-19
Olver, M.D. and Malan, D.D. (2000). The effect of choice-feeding from 7 weeks of age on the production characteristics of laying hens. South African Journal of Animal Science, 30, 110-114
Scott, M.L., Hull, S.J. and Mullenho, P.A. (1971). Calcium requirements of laying hens and effects of dietary oyster shell upon egg shell quality. Poultry Science, 50, 1055-1063
Selle, P.H., Cowieson, A.J. and Ravindran, V. (2009). Consequences of calcium interactions with phytate and phytase for poultry and pigs. Livestock Science, 124, 126-141
Steinruck, U., Rothe, F.X. and Kirchgebner, M. (1990). Selective feed intake of broilers during methionine deficiency. Arch. Geflugelk, 54:173-183.
Tamim, N.M., Angel, R. and Christman, M. (2004). Influence of dietary calcium and phytase on phytate phosphorus hydrolysis in broiler chickens. Poultry Science, 83, 1358-1367
Taylor, T.C. (1965). The availability of the calcium and phosphorus of plant materials for animals. Proceedings of the Nutrition Society, 24, 105-112
Underwood, E.J. and Suttle, N.F. 2001a. Calcium. In: Underwood, E. J. and Suttle, N. F. (eds.) The mineral nutrition of livestock. 3rd ed. Wallingford UK: Cabi
Underwood, E.J. and Suttle, N.F. 2001b. Phosphorus. In: Underwood, E. J. and Suttle, N. F. (eds.) The mineral nutrition of livestock. 3rd ed. Wallingford UK: Cabi
Wilkinson, S.J., Selle, P.H., Bedford, M.R. and Cowieson, A.J. (2014). Separate feeding of calcium improves performance and ileal nutrient digestibility in broiler chicks. Animal Production Science, 54, 172-178
Wood-Gush, D.G.M. and Kare, M.R. (1966). The behaviour of calcium-deficient chickens. British Poultry Science, 7, 285-290
Zuberbuehler, C.A., Messikommer, R.E. and Wenk, C. (2002). Choice feeding of selenium-deficient laying hens affects diet selection, selenium intake and body weight. Journal of Nutrition, 132, 3411-3417
Content from the event:
Related topics:
Authors:



dsm-firmenich

Recommend
Comment
Share

Would you like to discuss another topic? Create a new post to engage with experts in the community.












