I. INTRODUCTION
Broiler diets are formulated on the basis of ideal protein ratios (IPR) where selected amino acid requirements are expressed relative to lysine (100). However, IPR have been developed over decades for standard diets rather than diets with reduced-CP. This has led to the possibility that reduced-CP diets require modified IPR. For this reason, two sets of ratios were evaluated in diets with 210 and 180 g/kg CP in a 2 × 2 factorial design.
II. MATERIAL AND METHODS
This feeding study was conducted in compliance with the guidelines of the Animal Ethics Committee of The University of Sydney. Two dietary CP levels (210 and 180 g/kg) were tested under two ideal protein ratios (A and B) as shown in Table 1. All diets were formulated to have an energy density of 13.0 MJ/kg and 11 g/kg of digestible lysine (Table 2). The diets were cold-pelleted and contained xylanase enzyme. A total of 420 off-sex male Ross 308 broilers at 14 days of post-hatch were distributed to 28 floor pens (15 birds/pen) to have even distribution of average body weights and each of the four diet had seven replications. Weight gain, feed intake and FCRs were determined from 14 to 35 days post-hatch and carcass parameters were measured on 35 days post-hatch. JMP Pro 14 was used to perform analyses of variance and significance was determined at P < 0.05 using LSD student t-test.
Table 1 - Ideal protein ratios (IPR) used in the study
III. RESULTS
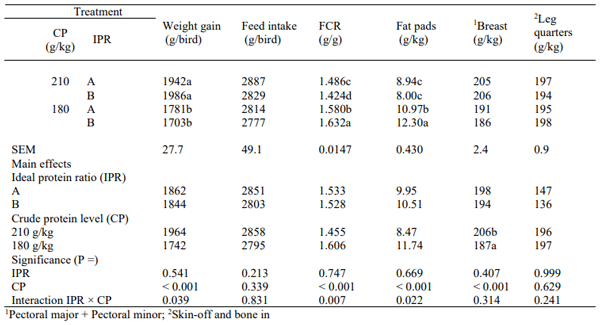
Growth performance from 14 to 35 days post-hatch and carcass parameters in broilers on 35 days post-hatch are shown in Table 3 and mortality rate was recorded as 2.4 % and not relevant to treatments. Overall weight gain was comparable to the Ross 308 male performance objectives (1853 versus 1849 g/bird) but feed conversion exceeded by 3.10% (1.531 versus 1.580) and reduced the feed intake by 3.3% (2826 versus 2921 g/bird). Significant interactions of dietary CP and ideal protein ratios were observed for weight gain (P = 0.039), FCR (P = 0.007) and relative fat pad weights (P = 0.022).
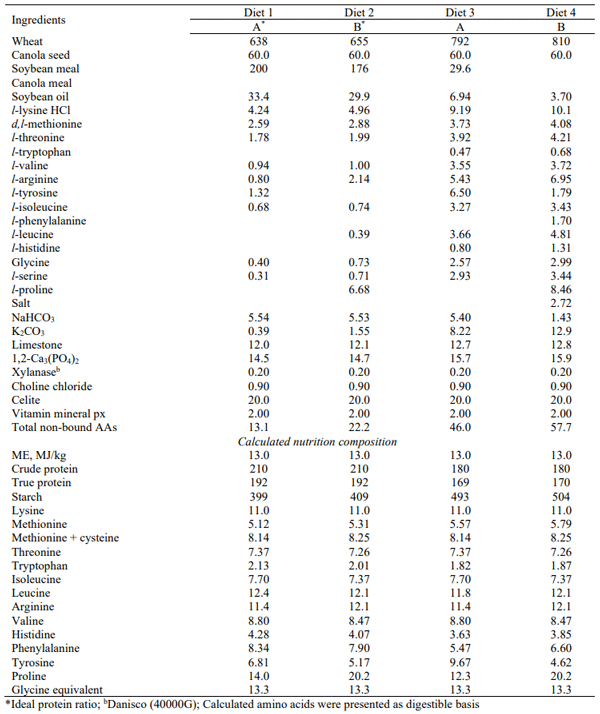
Table 2 - Composition and calculated nutrient specification in experimental diets (g/kg)
Table 3 - Growth performance from 14 to 35 days post-hatch and relative carcass parameters in broilers on 35 days post-hatch
Broilers offered ideal protein ratio B improved weight gain by 2.27% (1942 versus 1986 g) in comparison to ratio A in 210 g/kg CP diets and reduced weight gain by 4.58% (1781 versus 1703 g) in 180 g/kg CP diets. Ideal protein ratio B improved FCR by 4.35% in comparison to ratio A (1.424 versus 1.486) in 210 g/kg CP diets; whereas, ratio B compromised FCR by 3.29% (1.632versus 1.580) in 180 g/kg CP diets. Ideal protein ratios did not influence the relative fat pad weights in 210 g/kg CP diets, but ideal ratio B generated a 12.1% (12.30 versus 10.97 g/kg) increment in 180 CP diets. Broilers offered 210 g/kg CP diets showed a 10.2% higher (P < 0.001) breast muscle yield than those offered 180 CP diets (206 versus 187 g/kg) regardless of ideal protein ratios.
III. DISCUSSION
Treatment interactions between dietary crude protein level and ideal protein ratio were observed for weight gain (P < 0.05), FCR (P < 0.01) and relative abdominal fat-pad weights (P < 0.05). This indicates that IPR A was superior to B when birds were offered 180 g/kg CP diets by 4.58%, 3.29% and 10.8%, for weight gain, FCR, and relative fat-pad weights respectively. The situation was reversed when birds were offered 210 g/kg CP diets. Therefore, the proposition that appropriate IPRs are dependent on dietary CP levels was validated. From Table 1, it is evident that the only real discrepancy between the two ideal protein ratios for the nominated amino acids was for phenylalanine plus tyrosine at 116 and 102, relative to lysine. This meant, based on calculated nutrient specifications (Table 2), that 180 g/kg CP IPR A diets contained 15.14 g/kg phenylalanine plus tyrosine (138 relatives to lysine) as opposed to 11.22 g/kg (102 relatives to lysine) in IPR B diets.
Phenylalanine and tyrosine are interrelated as phenylalanine may be efficiently converted to tyrosine in poultry (Sasse and Baker, 1972). In addition to being incorporated into protein, both amino acids are related to circulating thyroid hormone levels in poultry (Elkin et al., 1980) and the synthesis of catecholamines, including dopamine, in brain tissue (Fernstrom and Fernstrom, 2017). Numerous recommendations have been put forward; however, Dorigam et al. (2013) concluded that the ideal ratio for phenylalanine plus tyrosine should be 115, relative to lysine. This suggests that the ratio of 102 in the IPR B reduced-CP diets may have been inadequate in comparison to the 138 ratio in IPR A diets in the present study. The analysed dietary amino acid concentrations should be more instructive when they come to hand. Nevertheless, the likelihood is that this differential contributed to the superiority of IPR A in reduced-CP diets.
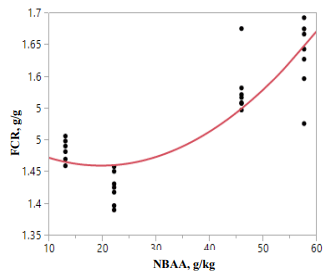
While speculative, there is another factor that may have contributed to the superiority of IPR A in reduced-CP diets. The 180 g/kg CP diets contained concentrations of either 46.0 g/kg (IPR A) or 57.7 g/kg (IPR B) non-bound amino acids. In Chrystal et al. (2020), 165 g/kg CP diets based on maize contained 38.5 g/kg as opposed to 49.4 g/kg non-bound amino acids in the corresponding wheat-based diets and the performance of the birds offered maize-based diets was clearly superior. However, across all diets in this study there was a highly significant quadratic relationship (r = 0.933; P < 0.0001), which indicated that FCR began to deteriorate once a concentration of 17.0 g/kg non-bound amino acids was exceeded. Moreover, in the present study there was a quadratic relationship (r = 0.861; P < 0.0001) between FCR and concentrations of non-bound amino acids (NBAA) in birds offered diets 1 to 4, inclusive, where:
y(FCR) = 1.353 + 0.00391*NBAA(g/kg) + 0.0001306*NBAA(g/kg)2 .
As shown in Figure 1, this indicates that FCR started to deteriorate when a concentration of 14.97 g/kg NBAA was exceeded.
These relationships may not be meaningful, but they could indicate that broiler chickens may not be able to accommodate high non-bound amino acid dietary inclusions. This contention was advanced by Pinchasov et al. (1990) who argued that there may be a need for a minimum quantity of ‘intact’ protein because intestinal uptakes of oligopeptides and non-bound amino acids are independent, but oligopeptides are absorbed more rapidly (Matthews and Abidi, 1976). Pinchasov et al. (1990) also raised the possibility that the relevant transport systems for non-bound amino acids may become saturated and that the difference in rates of intestinal uptakes may lead to amino acid imbalances at sites of protein synthesis. This is a real possibility as the digestive dynamics of non-bound versus protein-bound amino acids are inherently different (Liu and Selle, 2017).
Figure 1 - Quadratic relationship (r = 0.861; P < 0.0001) between concentrations of non-bound amino acids (NBAA) and FCR in birds offered diet 2 to 4, inclusive.
ACKNOWLEDGMENTS: This research was supported by an Australian Government Research Training Programme Scholarship (International) which was awarded to Shemil Priyan Macelline.
Presented at the 32th Annual Australian Poultry Science Symposium 2021. For information on the next edition, click here. 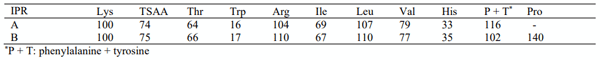





")









")