Introduction
Avian coccidiosis is one of the most costly infectious diseases affecting the commercial poultry industry [1]. Coccidia that infect chickens include Eimeria acervulina, E. tenella, E. maxima, E. brunetti, E. necatrix, E. praecox, and E. mitis. These apicomplexan protists invade cells of the intestinal epithelium, evoking necrotic tissue destruction and resulting in reduced body weight gain in broilers, decreased egg production in layers, and fecal shedding of viable parasites that re-infect susceptible animals upon ingestion. Over the preceding 40 years, avian coccidiosis has been mainly controlled by prophylactic chemotherapeutic drugs. More recently, the use of coccidia vaccines has reduced the need for in-feed medication [2]. Because host immunity to Eimeria infection is species-specific, currently available live, attenuated vaccines consist of mixtures of four or more Eimeria species. The basis of using live coccidia vaccines involves a continuous excretion/reingestion cycle of an initial low dose of parasites, which progressively induces protective flock immunity. However, live vaccines often lead to an early reduction in weight gain and may not be effective against regional antigenic variants absent from the formulation. Therefore, novel approaches are needed to more effectively control coccidiosis in commercial poultry flocks.
Vaccine delivery in conjunction with an adjuvant offers one means to increase potency. By definition, an adjuvant is an agent that stimulates the immune system and increases the host response to an antigen without itself conferring a specific antigenic effect [3]. Some adjuvants act by sequestering antigens in physically restricted areas, termed depots, to provide an extended time period of antigenic stimulation. This depot effect is essential for the efficacy of the majority of human and veterinary vaccine adjuvants, particularly with vaccines consisting of pathogen subunits (proteins, nucleic acids, and carbohydrates). Currently, aluminum hydroxide (alum) is only adjuvant approved for human use in the U.S. and Canada. Other adjuvants have been licensed for human use in Europe, including the water-in-oil (W/O) emulsions, MF59 and AS03, and the TLR4 agonist, monophosphoryl lipid A in alum [4]. A larger list of adjuvants is available for veterinary use [5].
The Montanide ISA series of adjuvants include the W/O emulsions, MontanideTM ISA 70 VG (ISA 70) and MontanideTM ISA 71 VG (ISA 71). Both formulations are mineral oil-based solutions incorporating a highly refined mannitol/oleic acid emulsifier [6]. ISA 71 is similar to ISA 70 except that it contains an improved mineral oil enabling the preferential stimulation of Th1-type cell-mediated immunity. ISA 70 and ISA 71 have been successfully applied to enhance immune response against pathogens of poultry, cattle, and small ruminants [7]. Our previous studies demonstrated that either ISA 70 or ISA 71 in conjunction with the Eimeria recombinant protein, profilin, was associated with enhanced protective immunity against experimental avian coccidiosis, as measured by increased body weight gain and/or decreased fecal oocyst shedding compared with profilin alone [8,9,10]. However, the specific effects were dependent upon the particular adjuvant used, the species of infecting Eimeria, and the parameter of infection measured. Profilin/ISA 70 and profilin/ISA 71 increased post-infection weight gains, but only following E. acervulina infection, whereas profilin/ISA 71, but not profilin/ ISA 70, decreased parasite shedding following infection with E. acervulina or E. tenella. In addition, we have demonstrated the utility of comparative microarray hybridization for identifying global transcriptional responses to various vaccination strategies that correlate with protection against experimental Eimeria infection [11,12,13]. Therefore, the current study was undertaken to compare the dynamics of lymphocyte transcriptome responses in chickens immunized with profilin/ISA 70 vs. profilin alone, or with profilin/ISA 71 vs. profilin alone, and to use this information to identify the better adjuvant with the potential for stimulating protective immunity against experimental avian coccidiosis.
Materials and Methods
Recombinant Profilin Protein and Adjuvants
Recombinant profilin, originally derived from E. acervulina, was expressed in Escherichia coli with a maltose binding protein epitope tag and purified by amylase affinity chromatography as described [8]. Purified profilin was emulsified with ISA 70 or ISA 71 at a 30:70 ratio (w:w, profilin:adjuvant) as recommended by the manufacturer (Seppic, Puteaux, France).
Chickens and Profilin Immunization
One-day-old Ross broiler chickens (Longenecker’s Hatchery, Elizabethtown, PA) were housed in Petersime starter brooder units and provided with feed and water ad libitum. At 7 days post-hatch, chickens were subcutaneously immunized with 50 mg of profilin emulsified in ISA 70 or ISA 71. Control chickens were immunized with PBS plus adjuvant, or with profilin in the absence of adjuvant. At 7 days post-primary immunization, the chickens were transferred to hanging cages (2 birds per cage) and given secondary subcutaneous booster injections identical with the primary immunization. All experiments were approved by the Beltsville Agricultural Research Center Small Animal Care and Use Committee.
Total RNA Preparation and Labeling
At 7 days post-secondary immunization, birds were sacrificed, single spleen cell suspensions were prepared, and lymphocytes were isolated by Percoll density gradient centrifugation as described [14]. Total RNA was isolated using Trizol (Invitrogen, Carlsbad, CA) and pooled into 2 equal aliquots (3 birds/sample). RNAs were amplified using the Two-Color Quick Amp Labeling Kit (Agilent Technologies, Santa Clara, CA) with cyanine 3 (Cy3)- or Cy5-labeled CTP. Labeled RNAs were purified using the RNeasy Mini Kit (Qiagen, Valencia, CA) and quantified with a Nanodrop ND-1000 UV-VIS spectrophotometer (Thermo Fisher Scientific, Waltham, MA).
Microarray Experimental Design
A standard reference design with hybridization of Cy3- and Cy5-labeled RNAs [15] was used to compare mRNA transcript levels in chickens immunized with profilin/ISA 70 vs. profilin alone and profilin/ISA 71 vs. profilin alone. Two biological replicates were conducted in each comparison with substitution of Cy3- and Cy5-labeled RNAs to prevent data distortion from sample labeling as previously described [12]. Labeled RNAs were hybridized to a Chicken Gene Expression Microarray (Agilent Technologies, Santa Clara, CA) containing 43,803 elements using the Gene Expression Hybridization Kit (Agilent Technologies) with constant mixing at 10 rpm for 17 hr at 65uC. After washing, microarray images were scanned, and data extraction and analysis were performed using Feature Extraction software version 10.7.3.1 (Agilent Technologies).
Microarray Data Analysis
GeneSpring GX10 software (Silicon Genetics, Redwood, CA) was used to qualify and normalize hybridization images, and to perform fold-change analyses as described [11,12]. Median signal intensities were qualified by subtracting the median local background and normalized by locally-weighted scatterplot smoothing (LOWESS). Flag information was applied to strain the spots with 100% valid values from each group and one-way analysis of variance (ANOVA) was performed to analyze the significance of differences between treatment groups. To generate signal ratios, signal channel values from the profilin/ISA 70 and profilin/ISA 71 groups were divided by the channel values from the profilin-only group. Modulated mRNA transcripts, defined as those with a cutoff of P,0.0005, were applied by asymptotic T-test analysis. The significantly differentially expressed transcripts were filtered using the Volcano Plot method [16] built by comparison among the various immunization groups. All microarray information and data was deposited online into the Gene Expression Omnibus (GEO) server (accession number GES 40743).
Bioinformatic Analysis
Differentially expressed transcripts were analyzed by Ingenuity Pathway Analysis (IPA) software (Ingenuity Systems, Redwood City, CA). Each identifier was mapped to its corresponding gene object in the Ingenuity Knowledge Base. Both up-regulated and down-regulated identifiers were defined as value parameters for the analysis. These focus genes were overlaid onto a global molecular network developed from information contained in the Ingenuity Knowledge Base. Biological functional analysis was performed and the canonical pathways from the datasets were mapped with the Ingenuity Pathways Knowledge Base. The Fischer’s exact test was used to calculate the probability that each biological function and associated pathways assigned to that dataset was statistically significant.
Experimental Eimeria Infection
At 7 days post-secondary immunization, chickens were uninfected or were orally infected with 1.0 x 104 sporulated oocysts of E. acervulina, E. tenella or E. maxima as described [17]. The coccidia parasites were originally isolated and maintained at the Animal and Natural Resources Institute, U.S. Department of Agriculture (Beltsville, MD). Prior to infection, sporulated oocysts were cleaned by floatation on 2.5% sodium hypochlorite, washed three times with PBS, and enumerated using a hemocytometer.
Body Weight Gains and Fecal Oocyst Shedding
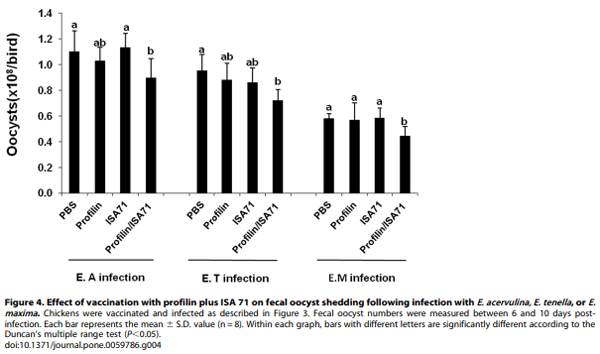
Body weights of uninfected and Eimeria-infected chickens (8/ group) were measured at 0 and 10 days post-infection. For determination of fecal oocysts numbers, birds (8/group) were placed on wire oocyst collection cages, fecal samples were collected between 6 and 10 days post-infection, and total oocysts were enumerated using a McMaster counting chamber as described [18].
Anti-profilin Serum Antibody Levels
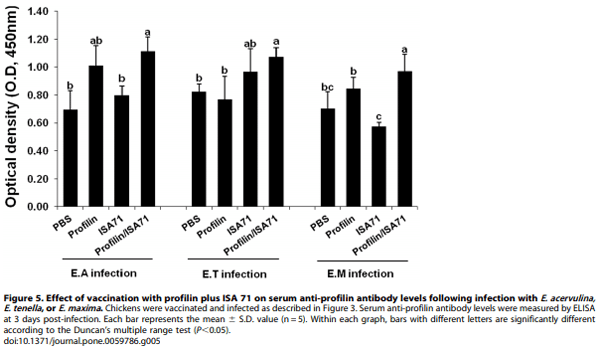
At 3 days post-infection, chickens (5/group) were euthanized by cervical dislocation, blood was collected by cardiac puncture, and sera were prepared by low speed centrifugation. Serum antibodies against profilin were measured by enzyme-linked immunosorbent assay (ELISA) as described [10,18]. Ninety-six-well microtiter plates were coated overnight with 10 mg/well of purified recombinant profilin, washed with PBS containing 0.05% Tween 20, and blocked with PBS containing 1% BSA. Serum samples were added and incubated for 1 hr with continuous shaking, the plates were washed, and bound antibody was detected with peroxidase-conjugated rabbit anti-chicken IgG secondary antibody and peroxidase-substrate (Sigma, St. Louis, MO). Optical density (OD) values at 450 nm were measured with an automated microplate reader (Bio-Rad, Richmond, CA).
Intestinal Cytokine mRNA Levels
At 3 day post-infection, chickens (3/group) were euthanized by cervical dislocation and the intestinal duodenum (E. acervulina-infected), jejunum (E. tenella-infected), and cecum (E. maxima-infected) were removed. Tissues were incised longitudinally, gently washed with ice-cold Hank’s Balanced Salt Solution (Sigma), and intraepithelial lymphocytes (IELs) were isolated by density gradient centrifugation as described [19,20]. Total RNA was isolated, 5.0 mg were incubate with 1.0 U of DNase I and 1.0 ml of 10 x DNase I reaction buffer (Sigma) for 15 min at room temperature, 1.0 ml of stop solution was added, and the mixture was heated at 70uC for 10 min. RNA was reverse-transcribed using the StrataScript first-strand synthesis system (Stratagene, La Jolla, CA) according to the manufacturer’s recommendations. PCR amplification and detection were carried out using equivalent amounts of total RNA and oligonucleotide primers for IFN-c, IL-2, IL-10, IL-17A, and the glyceraldehydes-3-phosphate dehydrogenase (GAPDH) internal control (Table 1) with the Mx3000P system and Brilliant SYBR Green QPCR master mix (Stratagene). Standard curves were generated using log10 diluted standard RNAs and the levels of individual transcripts were normalized to those of GAPDH by the Q-gene program [21]. To normalize RNA levels between samples within an experiment, the mean threshold cycle (Ct) values for the amplification products were calculated by pooling values from all samples in that experiment. Each analysis was performed in triplicate.
Statistical Analysis
Body weight gains, oocyst shedding, anti-profilin antibody titers, and cytokine levels were expressed as means ± SD. Mean values of different treatment groups were compared using ANOVA or the Duncan’s multiple range test with SPSS 15.0 for Windows (SPSS Inc., Chicago, IL). Differences between means were considered statistically significant at P,0.05.
Results
Spleen Lymphocyte Transcript Levels Following Immunization with Profilin Plus ISA 70 or Profilin Plus ISA 71
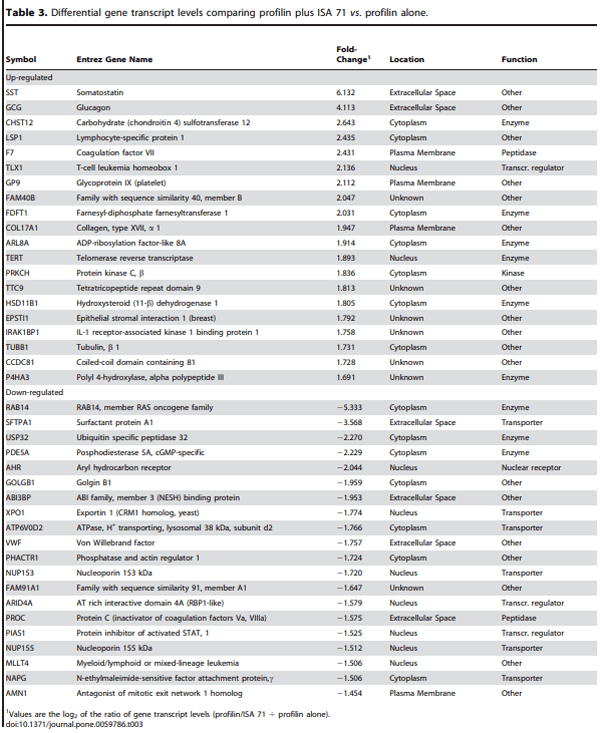
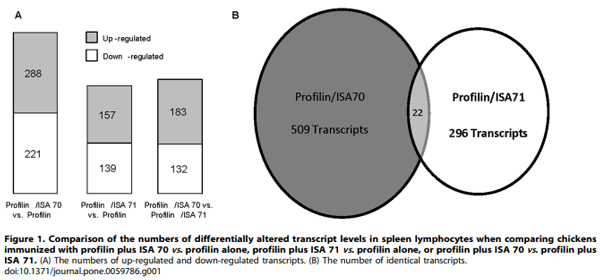
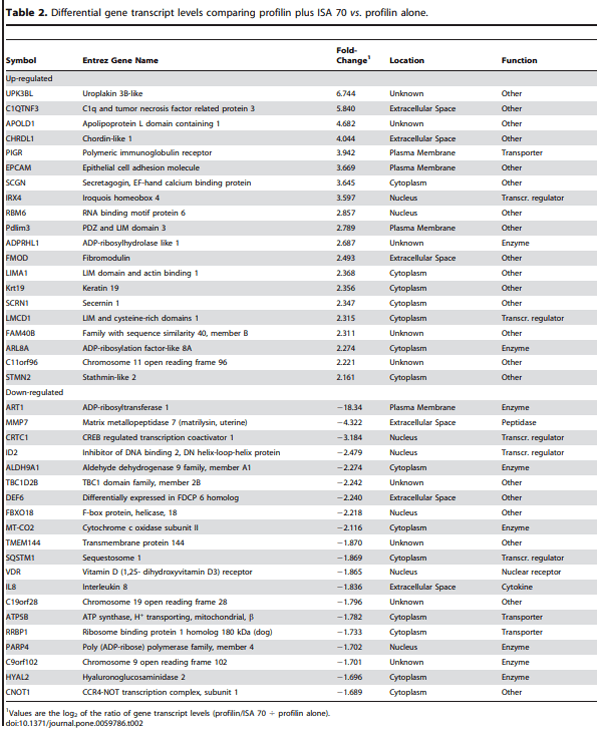
Microarray hybridizations were performed using the Agilent Technology Chicken Gene Expression Microarray with total RNAs isolated from spleen lymphocytes of chickens immunized with profilin alone, profilin plus ISA 70, or profilin plus ISA 71 to identify global transcriptome changes in the respective treatment groups. A critical P value of 0.0005 was employed to compare transcript levels in the profilin/ISA 70 vs. profilin alone, the profilin/ISA 71 vs. profilin alone, and the profilin/ISA 70 vs. profilin/ISA 71 groups. From this dataset, the levels of 509 (288 up-regulated, 221 down-regulated), 296 (157 up-regulated, 139 down-regulated), and 315 (183 up-regulated, 132 down-regulated) mRNAs were identified as differentially altered in the denoted comparisons (Figure 1A). Twenty-two altered transcripts were identical in the profilin/ISA 70 vs. profilin alone (11 up-regulated, 11 down-regulated) and the profilin/ISA 71 vs. profilin alone (10 up-regulated, 12 down-regulated) groups (Figure 1B). Tables 2 and 3 list the 20 most up-regulated and 20 most down-regulated transcripts for these latter two comparisons in decreasing order of fold-change.
Biological Function and Pathway Analysis of Differentially Regulated Splenocyte Transcripts
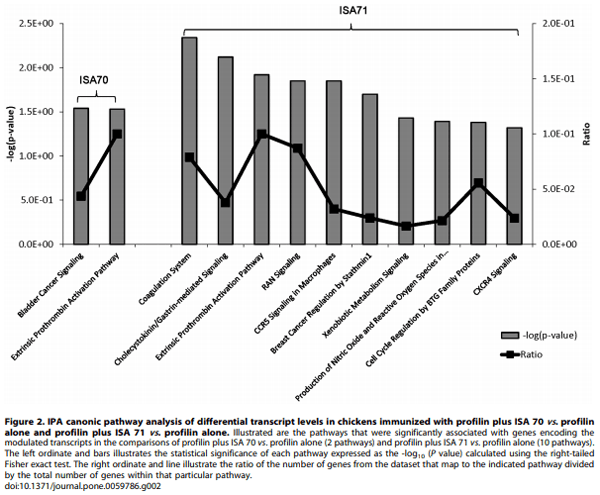
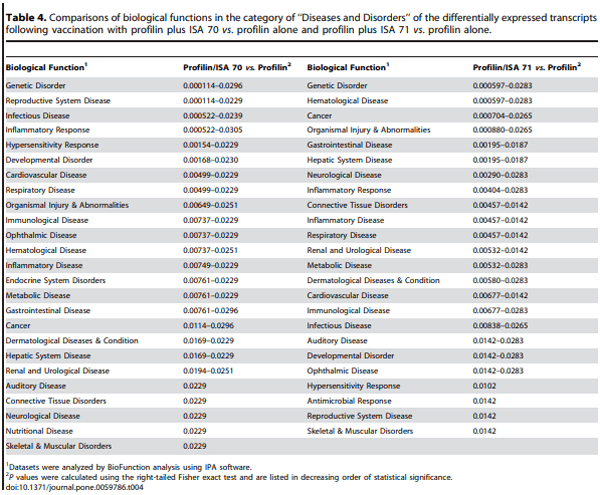
The differently modified datasets were mapped to the corresponding genes of the human, mouse, and rat genomes using Ingenuity Knowledge Base software. From this analysis, 192 chicken genes were identified and annotated in the profilin/ISA 70 vs. profilin alone comparison, and 112 genes in the profilin/ISA 71 vs. profilin alone comparison. Biological function analysis using the IPA database identified the category ‘‘Disease and Disorder’’ as the most significantly associated with the genes identified in both comparisons, with 25 and 24 significantly associated biological functions respectively (Table 4). Of these, two biological functions were uniquely associated with the profilin/ISA 70 vs. profilin alone comparison, ‘‘Endocrine System Disorders’’ and ‘‘Nutritional Disease’’, and one biofunction was exclusively associated with the profilin/ISA 71 vs. profilin alone comparison, ‘‘Antimicrobial Response’’. Finally, the IPA database was used to identify the canonical pathways associated with the respective biofunctions of the two comparison groups. Two pathways were identified in the profilin/ISA 70 vs. profilin alone comparison, while 10 pathways were recognized in the profilin/ISA 71 vs. profilin alone comparison (Figure 2).
Effect of Vaccination with Profilin Plus ISA 71 on Body Weight Gain and Oocyst Shedding
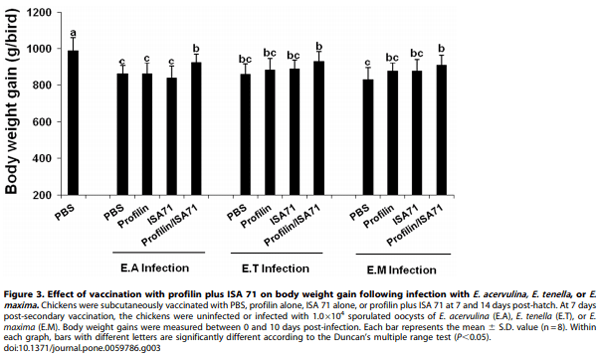
Because immunization with profilin plus ISA 71 vs. profilin alone was associated with more unique biological functions, as well as a larger number of total genes and pathways associated with these functions, compared with profilin/ISA 70 vs. profilin alone, we focused on ISA 71 as a possible adjuvant for in vivo protection studies against experimental Eimeria infection following profilin vaccination. Chickens vaccinated with profilin plus ISA 71 and infected with E. acervulina had increased body weight gains between 0 and 10 days post-infection compared with infected chickens vaccinated with profilin alone (Figure 3). Profilin/ISA 71- vaccinated and E. maxima-infected animals had decreased oocyst fecal shedding between days 6 and 10 post-infection compared with infected chickens vaccinated with profilin alone (Figure 4). Finally, profilin/ISA 71-vaccinated birds that were infected with E. tenella- or E. maxima had greater anti-profilin serum antibody levels at 3 days post-infection compared with infected birds vaccinated with profilin alone (Figure 5).
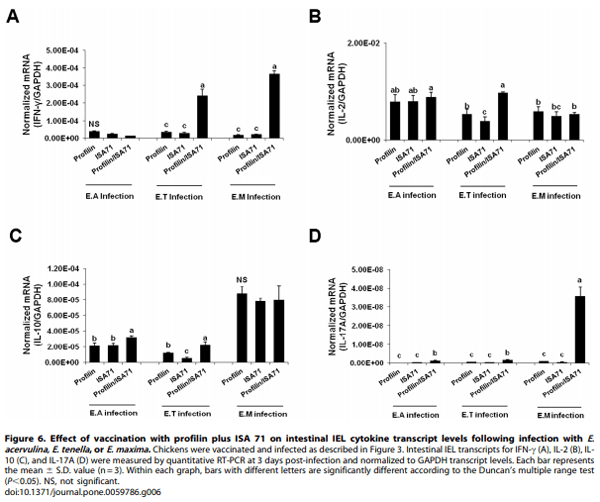
Effects of Vaccination with Profilin Plus ISA 71 Adjuvant on Cytokine Transcript Levels in Intestinal IELs
Chickens vaccinated with profilin plus ISA 71 and infected with all 3 Eimeria parasites had greater levels of intestinal IEL gene transcripts encoding IFN-c, IL-2, IL-10, and/or IL-17A at 3 days post-infection compared with infected birds vaccinated with profilin alone (Figure 6). The greatest cytokine transcriptional response was seen in the E. tenella-infected group. More specifically, E. tenella-infected animals had increased levels of all 4 transcripts, while E. acervulina-infected birds had greater IL-10 and IL-17A mRNA levels and E. maxima-infected birds had increased levels of IFN-γ and IL-17A mRNAs.
Discussion
This study demonstrated that chickens immunized with profilin plus ISA 70 or profilin plus ISA 71 exhibited spleen lymphocyte transcriptome changes, compared with immunization with profilin alone, that were consistent with alteration of levels of mRNAs encoded by genes belonging to the IPA database category ‘‘Disease and Disorder’’. Compared with profilin/ISA 70 vs. profilin alone, the profilin/ISA 71 vs. profilin alone comparison was associated with more unique biological functions, and a larger number of genes and pathways associated with these functions, suggesting that vaccination with profilin/ISA 71 may induce a greater protective host response to experimental Eimeria infection. Compared with profilin alone, vaccination with profilin/ISA 71 was correlated with (a) increased body weight gains following E. acervulina infection, (b) reduced parasite fecal shedding following E. maxima infection, (c) augmented anti-profilin serum antibody titers in E. maxima- and E. tenella-infected chickens, and (d) higher IFN-c, IL-2, IL-10, and IL-17A transcript levels in gut IELs of E. acervulina-, E. maxima-, and/or E. tenella-infected chickens. Weight gains and parasite shedding were equal in the ISA 71 alone vs. PBS negative control groups, and IEL transcript levels were generally equivalent in the ISA 71 only vs. profilin alone groups, indicating that the adjuvant itself was not responsible for these effects. Collectively, these data suggest that vaccination of chickens with profilin plus ISA 71 may increase resistance against experimental avian coccidiosis by selective Eimeria species.
In general, subunit vaccines against many infectious diseases of livestock and poultry in the absence of adjuvants are weakly immunogenic, and repeated vaccinations are often needed to generate sufficient protective immunity to control infection [22]. While immunologic adjuvants are known to stimulate the host’s immune system response to a target antigen, without themselves conferring immunity, the limited availability of safe and efficacious adjuvants for veterinary use hampers disease control strategies against some of the more common infectious pathogens of food animals. The Montanide ISA series of adjuvants are ready-to-use W/O, O/W or W/O/W emulsions, incorporating high-grade, injectable mineral or non-mineral oils. Proven benefits in veterinary medicine include the production of stable vaccine emulsions with low viscosity, ease of injection, reduced toxicity, and induction of a strong, long-lasting immune response [6,7,23,24,25,26,27]. Montanide ISA adjuvants appear to be ideally suited for use with vaccines of limited immunogenicity, such as profilin, an intracellular component that contributes to actin-dependent gliding motility and host cell invasion of apicomplexan parasites, including Toxoplasma gondii and Eimeria spp. [28]. T. gondii profilin binds to Toll-like receptor-11, inducing a potent IL-12 response in dendritic cells, and resulting in enhanced humoral and Th1 cellular immune responses against the parasite [28,29]. The E. acervulina protein, 3-1E, was identified earlier in merozoites of the microorganism as an immunogenic component of the parasite which induced high levels of antigen-specific proliferation and IFN-c production by chicken splenic lymphocytes [30]. Polyclonal antibodies raised against E. acervulina 3-1E cross-reacted with the homologous proteins of E. tenella and E. maxima. Subsequently, 3-1E was shown to be the Eimeria homologue of T. gondii profilin [28].
We previously reported that compared with unimmunized controls, chickens immunized with profilin in the absence of adjuvant had altered levels of 127 gene transcripts (71 upregulated, 56 down-regulated). The total number of transcripts affected by profilin/ISA 70 vs. profilin alone (509) or profilin/ISA 71 vs. profilin alone (296) observed in the current investigation is comparable to the results of our previous studies using other adjuvants and immunomodulators. More specifically, compared with chickens immunized with profilin alone, chickens given profilin plus the Quil A/cholesterol/dimethyl dioctadecyl ammonium bromide/Carbopol (QCDC) adjuvant mixture had 164 altered mRNAs (60 up-regulated, 104 down-regulated), and birds immunized with profilin plus QCDC incorporating the Bay R1005 immunostimulant (QCDCR) had 233 modulated transcripts (103 up-regulated, 130 down-regulated) [31]. In a subsequent study in the absence of profilin vaccination, dietary supplementation of chickens with propyl thiosulfinate, a secondary metabolite of garlic with immunoenhancing properties, identified 1,227 transcripts whose levels were altered in intestinal IELs compared with untreated controls [13]. As in the current report, biological pathway analysis identified the propyl thiosulfinate-altered transcripts to be encoded by genes associated with the IPA category ‘‘Disease and Disorder’’.
Interestingly, whereas ISA 70 and ISA 71 are composed of similar adjuvant formulations, only 22 common transcripts were shared between the profilin/ISA 70 vs. profilin alone and the profilin/ISA 71 vs. profilin alone groups, representing 4.3% and 7.4% of the total number of altered mRNAs, respectively. On the other hand, comparison of profilin/ISA 70-immunized chickens with the profilin/ISA 71 group identified 315 altered transcripts, indicating that the number of dissimilar mRNAs was substantially greater than the number of shared transcripts. Correspondingly, a similar comparison using the QCDC and QCDCR adjuvants revealed 397 altered transcripts in the profilin/QCDC vs. profilin/ QCDCR groups [31]. In another study, comparative microarray analysis between uninfected vs. E. acervulina, E. tenella, or E. maxima infections was used to identify commonly altered transcripts in these 3 denoted groups [12]. Following E. acervulina infection, 2,431 mRNAs were altered, while infection with E. tenella and E. maxima modulated the levels of 2,522 and 1,717 mRNAs respectively. From these, 766 transcripts were common to E. acervulina and E. tenella, 319 were shared between E. acervulina and E. maxima, 289 were common to E. tenella and E. maxima, and 361 mRNAs were shared between all 3 infections. Taken together, these results indicate that infection with intact, viable coccidia parasites stimulates a greater host transcriptional response compared with profilin vaccination in the presence or absence of adjuvant.
Body weight gain and fecal oocyst shedding are reliable clinical signs for the evaluation of protective immunity in avian coccidiosis [32]. Both parameters are directly correlated with the levels of intestinal proinflammatory cytokines in Eimeria-infected chickens [33,34,35]. IFN-c plays a critical role in the Eimeria-stimulated host immune response, and is one of the earliest cytokines detected in infected intestinal mucosa [36]. Indeed, IFN-c is the dominant cytokine elicited in the gut of Eimeria-infected chickens that typifies the Th1 cell-mediated immune response seen during experimental avian coccidiosis [37]. Endogenous IFN-c production in gut epithelia was positively associated with improved body weight gain and decreased oocyst shedding in birds following Eimeria infection [33]. Chickens treated exogenously with purified recombinant IFNc protein showed greater weight gain and reduced fecal oocyst numbers following E. acervulina infection [38]. IL-2 and IL-17A are other members of this Th1 cytokine response that serve to recruit, activate, and amplify immune effector leukocytes with cytotoxic activity against coccidia parasites [39,40]. On the other hand, IL10 driven inhibition of IFN-c production suggests that this counter-regulatory mediator may favor a shift toward a Th2 response later in the course of infection, and prevent tissue damage as a consequence of uncontrolled intestinal inflammation [41]. Therefore, augmented production of IFN-c, IL-2, IL-17A, and IL10 by profilin plus ISA 71 appears to preserve the natural balance of pro- and anti-inflammatory pathways in the gut that are necessary for an effective cellular immune response against the invading parasite while maintaining tissue homeostasis.
In conclusion, this study identified transcriptome dynamics in chickens following immunization with the Eimeria recombinant profilin protein in combination with either the ISA 70 or ISA 71 W/O adjuvants by comparison with immunization with profilin alone. Based on the greater transcriptional response elicited by profilin/ISA 71, this antigen/adjuvant mixture was used to subsequently demonstrate increased protection against experimental avian coccidiosis, as assessed by augmented body weight gains, decreased parasite fecal shedding, greater anti-profilin serum antibody titers, and increased levels of cytokine gene transcripts compared with vaccination with profilin alone. These results suggest that profilin in conjunction with ISA 71 provides an effective means of eliciting humoral and cellular immune responses with the potential to generate protective immunity against Eimeria infection.
Acknowledgments
The authors thank Margie Nichols, Ashley Cox, Bo Hui Hong, and Stacy Torreyson for their significant contribution to this research.
Author Contributions
Conceived and designed the experiments: SIJ DKK HSL SHL KWL SD. Performed the experiments: SIJ DKK SHL KWL. Analyzed the data: SIJ DKK HSL SHL EPL. Contributed reagents/materials/analysis tools: FB LD SD JBA. Wrote the paper: SIJ DKK EPL.
This article was originally published in PLoS ONE 8(4): e59786. doi:10.1371/journal.pone.0059786. This is an Open Access article free of all copyright, and may be freely reproduced, distributed, transmitted, modified, built upon, or otherwise used by anyone for any lawful purpose. The work is made available under the Creative Commons CC0 public domain dedication.










.jpg&w=3840&q=75)