Salmonella Harborage Sites in Infected Poultry That May Contribute to Contamination of Ground Meat
Author details:
1 Department of Population Health, Poultry Diagnostic and Research Center, University of Georgia, Athens, GA, United States; 2 Center for Food Safety, College of Agricultural and Environmental Sciences, University of Georgia, Griffin, GA, United States; 3 Department of Veterinary Medicine, Universidade Federal do Parana, Curitiba, Brazil; 4 Department of Population Health, University of Georgia, Athens, GA, United States; 5 Department of Biomedical Sciences and Pathobiology, Virginia-Maryland College of Veterinary Medicine, Virginia Tech, Blacksburg, VA, United States.
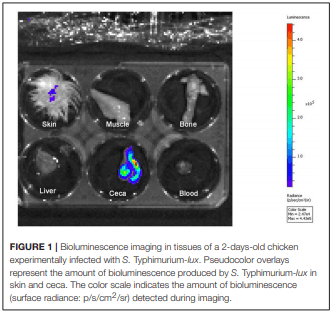
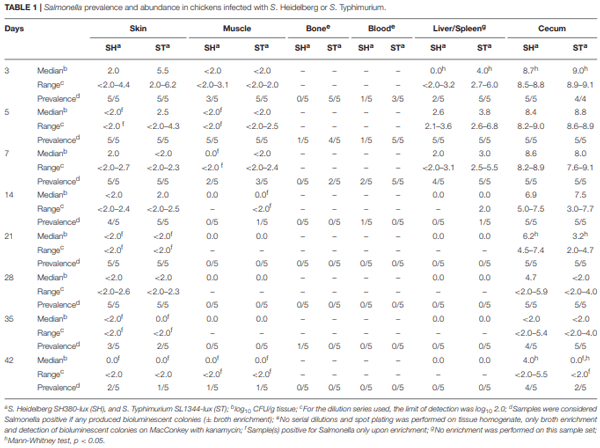
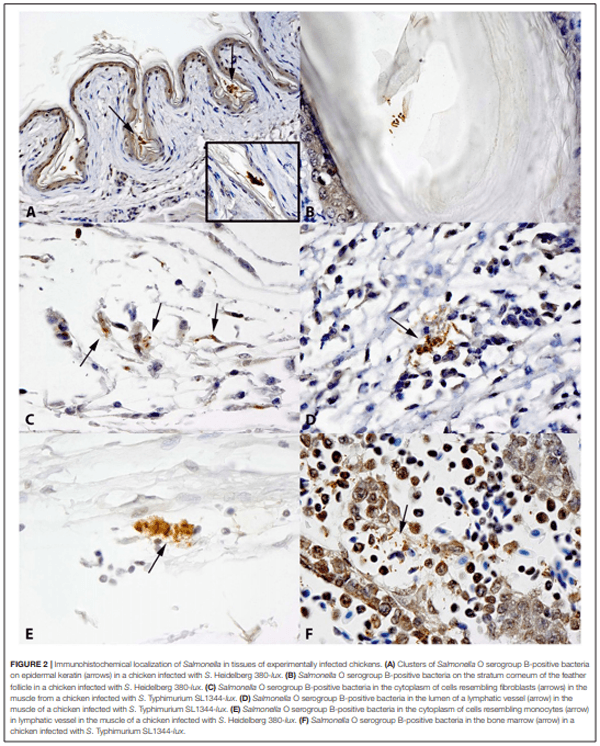
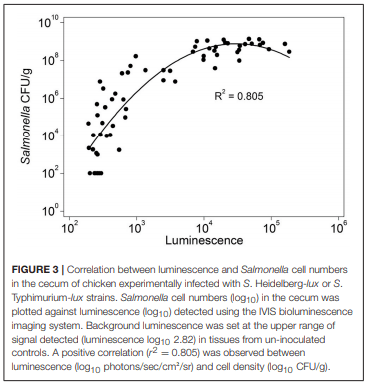
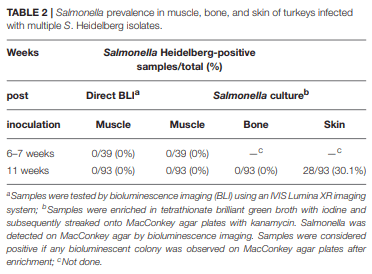
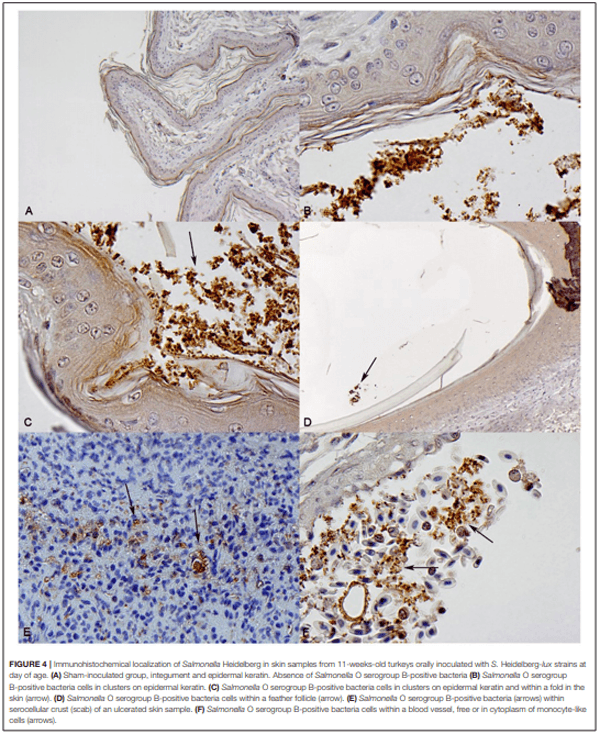
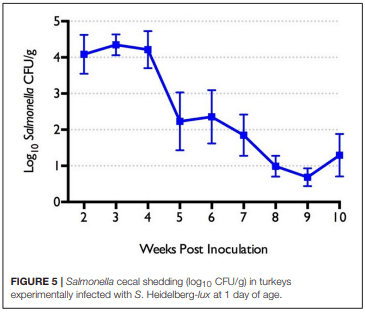
The role of invasive Salmonella in contamination of ground poultry is poorly defined. Salmonella harborage sites were determined in experimentally infected chickens and turkeys. Bioluminescent-tagged Salmonella were used to follow their spread in bone, meat, and skin following infection. Immunohistochemistry and culture were used to localize Salmonella. Chicken neck skin was positive for S. Heidelberg and S. Typhimurium throughout the experimental period, and the bacteria were localized on the stratum corneum of the epidermis and feather follicles. S. Heidelberg and S. Typhimurium were intermittently detected in drumstick muscle of chickens, with Salmonella primarily localized in connective tissues and lymphatics. Twenty percent of the drumstick muscles were culture positive for Salmonella in chickens at 42 days of age. Blood and tibiotarsus bone were culture positive for S. Heidelberg and S. Typhimurium during the first 2 weeks of infection. Salmonella levels in neck skin and muscle were <102 CFU/g in chickens at 42 days of age. Multiple S. Heidelberg isolates associated with foodborne outbreaks were used to infect chickens and turkeys to determine whether some strains attained high abundance in the muscle of infected birds. No chicken drumsticks or thighs were Salmonella positive by bioluminescence in chickens at 42 days of age (n = 210). In turkeys, all drumstick muscles (n = 132) and tibiotarsus bone (n = 93) were negative for S. Heidelberg. Thirty percent of the breast skins (n = 93) were culture positive for S. Heidelberg when turkey hens were 11 weeks old. S. Heidelberg were primarily localized on the stratum corneum of the epidermis in turkeys. Exclusion of skin from ground poultry products may be the best option for reducing Salmonella contamination in ground chicken and ground turkey.
Keywords: Salmonella, Heidelberg, Typhimurium, chicken, turkey, bioluminescence imaging, immunohistochemistry.
Barrow, P. A., Huggins, M. B., Lovell, M. A., and Simpson, J. M. (1987). Observations on the pathogenesis of experimental Salmonella typhimurium infection in chickens. Res. Vet. Sci. 42, 194–199. doi: 10.1016/S0034-5288(18)30685-4
Burns-Guydish, S. M., Olomu, I. N., Zhao, H., Wong, R. J., Stevenson, D. K., and Contag, C. H. (2005). Monitoring age-related susceptibility of young mice to oral Salmonella enterica serovar Typhimurium infection using an in vivo murine model. Pediatr. Res. 58, 153–158. doi: 10.1203/01.PDR.0000157725.44213.C4
CDC. (2011). Multistate Outbreak of Human Salmonella Heidelberg Infections Linked to Ground Turkey. Atlanta, GA. Available online at: http://www.cdc.gov/ salmonella/2011/ground-turkey-11-10-2011.html (accessed June 21, 2016).
CDC. (2014a). Multistate Outbreak of Multidrug-Resistant Salmonella Heidelberg Infections Linked to Foster Farms Brand Chicken. Atlanta, GA. Available online at: http://www.cdc.gov/salmonella/heidelberg-10-13/index.html (accessed June 21, 2014).
CDC. (2014b). Salmonella. Atlanta, GA. Available online at: http://www.cdc.gov/ salmonella/ (accessed June 21, 2016).
Chappell, L., Kaiser, P., Barrow, P., Jones, M. A., Johnston, C., and Wigley, P. (2009). The immunobiology of avian systemic salmonellosis. Vet. Immunol. Immunopathol. 128, 53–59. doi: 10.1016/j.vetimm.2008. 10.295
Cheminay, C., Möhlenbrink, A., and Hensel, M. (2005). Intracellular Salmonella inhibit antigen presentation by dendritic cells. J. Immunol. 174, 2892–2899. doi: 10.4049/jimmunol.174.5.2892
Cheng, Y., Pedroso, A. A., Porwollik, S., Mcclelland, M., Lee, M. D., Kwan, T., et al. (2015). rpoS-Regulated core genes involved in the competitive fitness of Salmonella enterica Serovar Kentucky in the intestines of chickens. Appl. Environ. Microbiol. 81, 502–514. doi: 10.1128/AEM.03219-14
Collinson, S. K., Doig, P. C., Doran, J. L., Clouthier, S., Trust, T. J., and Kay, W. W. (1993). Thin, aggregative fimbriae mediate binding of Salmonella enteritidis to fibronectin. J. Bacteriol. 175, 12–18. doi: 10.1128/jb.175.1.12-18.1993
Cui, Y., Guran, H. S., Harrison, M. A., Hofacre, C. L., and Alali, W. Q. (2015). Salmonella levels in turkey neck skins, drumstick bones, and spleens in relation to ground turkey. J. Food Prot. 78, 1945–1953. doi: 10.4315/0362-028X.JFP-15-240
Engebrecht, J., and Silverman, M. (1984). Identification of genes and gene products necessary for bacterial bioluminescence. Proc. Natl. Acad. Sci. U.S.A. 81, 4154–4158. doi: 10.1073/pnas.81.13.4154
Erol, I., Goncuoglu, M., Ayaz, N. D., Ellerbroek, L., Ormanci, F. S., and Kangal, O. I. (2013). Serotype distribution of Salmonella isolates from turkey ground meat and meat parts. Biomed Res. Int. 2013, 281591. doi: 10.1155/2013/2 81591
Evans, N. P., Collins, D. A., Pierson, F. W., Mahsoub, H. M., Sriranganathan, N., Persia, M. E., et al. (2017). Investigation of medium chain fatty acid feed supplementation for reducing Salmonella typhimurium colonization in Turkey poults. Foodborne Pathog. Dis. 14, 531–536. doi: 10.1089/fpd.2016.2273
Evans, N. P., Evans, R. D., Regalado, J., Sullivan, J. F., Dutta, V., Elvinger, F., et al. (2015). Preharvest Salmonella detection for evaluation of fresh ground poultry product contamination. J. Food Prot. 78, 1266–1271. doi: 10.4315/0362-028X.JFP-14-509
FSIS (2013). Ground Poultry and Food Safety, on United States Department of Agriculture. Available online at: http://www.fsis.usda.gov/wps/portal/fsis/ topics/food-safety-education/get-answers/food-safety-fact-sheets/poultrypreparation/ground-poultry-and-food-safety/CT_Index (accessed June 8, 2016).
FSIS (2015). Meat and Poultry Labeling Terms, on United States Department of Agriculture. Available online at: http://www.fsis.usda.gov/wps/portal/ fsis/topics/food-safety-education/get-answers/food-safety-fact-sheets/foodlabeling/meat-and-poultry-labeling-terms (accessed June 8, 2016).
Gast, R. K. (2013). “Paratyphoid infections,” in Diseases of Poultry, 13th Edn., eds D. E. Swayne, J. R. Glisson, L. R. Mcdougald, L. K. Nolan, D. L. Suarez, and V. Nair (Ames, IA: Wiley-Blackwell), 693–706.
Gast, R. K., and Holt, P. S. (1998). Persistence of Salmonella enteritidis from one day of age until maturity in experimentally infected layer chickens. Poult. Sci. 77, 1759–1762. doi: 10.1093/ps/77.12.1759
Gerstel, U., and Römling, U. (2001). Oxygen tension and nutrient starvation are major signals that regulate agfD promoter activity and expression of the multicellular morphotype in Salmonella typhimurium. Environ. Microbiol. 3, 638–648. doi: 10.1046/j.1462-2920.2001.00235.x
Howe, K., Karsi, A., Germon, P., Wills, R. W., Lawrence, M. L., and Bailey, R. H. (2010). Development of stable reporter system cloning luxCDABE genes into chromosome of Salmonella enterica serotypes using Tn7 transposon. BMC Microbiol. 10:197. doi: 10.1186/1471-2180-10-197
Kassem, I. I., Sanad, Y., Gangaiah, D., Lilburn, M., Lejeune, J., and Rajashekara, G. (2010). Use of bioluminescence imaging to monitor Campylobacter survival in chicken litter. J. Appl. Microbiol. 109, 1988–1997. doi: 10.1111/j.1365-2672.2010.04828.x
Kim, K. Y., Frank, J. F., and Craven, S. E. (1996). Three-dimensional visualization of Salmonella attachment to poultry skin using confocal scanning laser microscopy. Lett. Appl. Microbiol. 22, 280–282. doi: 10.1111/j.1472-765X.1996.tb01161.x
Lee, N. Y., Park, S. Y., Kang, I. S., and Ha, S. D. (2014). The evaluation of combined chemical and physical treatments on the reduction of resident microorganisms and Salmonella typhimurium attached to chicken skin. Poult. Sci. 93, 208–215. doi: 10.3382/ps.2013-03536
Lillard, H. S. (1989). Factors affecting the persistence of Salmonella during the processing of poultry. J. Food Prot. 52, 829–832. doi: 10.4315/0362-028X-52.11.829
Lillard, H. S. (1993). Bactericidal effect of chlorine on attached Salmonellae with and without sonification. J. Food Prot. 56, 716–717. doi: 10.4315/0362-028X-56.8.716
Mastroeni, P., and Grant, A. J. (2011). Spread of Salmonella enterica in the body during systemic infection: unravelling host and pathogen determinants. Expert Rev. Mol. Med. 13, e12. doi: 10.1017/S1462399411001840
Maurer, J. J., Brown, T. P., Steffens, W. L., and Thayer, S. G. (1998). The occurrence of ambient temperature-regulated adhesins, curli, and the temperaturesensitive hemagglutinin tsh among avian Escherichia coli. Avian Dis. 42, 106–118. doi: 10.2307/1592582
Meighen, E. A., and Dunlap, P. V. (1993). Physiological, biochemical and genetic control of bacterial bioluminescence. Adv. Microb. Physiol. 34, 1–67. doi: 10.1016/S0065-2911(08)60027-2
Nde, C. W., Mcevoy, J. M., Sherwood, J. S., and Logue, C. M. (2007). Cross contamination of turkey carcasses by Salmonella species during defeathering. Poult. Sci. 86, 162–167. doi: 10.1093/ps/86.1.162
Ochman, H., Soncini, F. C., Solomon, F., and Groisman, E. A. (1996). Identification of a pathogenicity island required for Salmonella survival in host cells. Proc. Natl. Acad. Sci. U.S.A. 93, 7800–7804. doi: 10.1073/pnas.93.15.7800
Olsén, A., Arnqvist, A., Hammar, M., and Normark, S. (1993). Environmental regulation of curli production in Escherichia coli. Infect. Agents Dis. 2, 272–274.
Özkaya, H., Akcan, A. B., Aydemir, G., Aydinöz, S., Razia, Y., Gammon, S. T., et al. (2012). Salmonella typhimurium infections in BALB/c mice: a comparison of tissue bioluminescence, tissue cultures and mice clinical scores. New Microbiol. 35, 53–59.
Phillips, R. A., and Opitz, H. M. (1995). Pathogenicity and persistence of Salmonella enteritidis and egg contamination in normal and infectious bursal disease virus-infected leghorn chicks. Avian Dis. 39, 778–787. doi: 10.2307/1592414
Rigney, C. P., Salamone, B. P., Anandaraman, N., Rose, B. E., Umholtz, R. L., Ferris, K. E., et al. (2004). Salmonella serotypes in selected classes of food animal carcasses and raw ground products, January 1998 through December 2000. J. Am. Vet. Med. Assoc. 224, 524–530. doi: 10.2460/javma.2004.224.524
Ritchie, J. M., Campbell, G. R., Shepherd, J., Beaton, Y., Jones, D., Killham, K., et al. (2003). A stable bioluminescent construct of Escherichia coli O157:H7 for hazard assessments of long-term survival in the environment. Appl. Environ. Microbiol. 69, 3359–3367. doi: 10.1128/AEM.69.6.3359-3367.2003
Salehi, S., Howe, K., Lawrence, M. L., Brooks, J. P., Bailey, R. H., and Karsi, A. (2017). Salmonella enterica Serovar Kentucky flagella are required for broiler skin adhesion and Caco-2 cell invasion. Appl. Environ. Microbiol. 83:e02115-16. doi: 10.1128/AEM.02115-16
Swart, A. L., and Hensel, M. (2012). Interactions of Salmonella enterica with dendritic cells. Virulence 3, 660–667. doi: 10.4161/viru.22761
USDA-FSIS (2014). Progress Report on Salmonella and Campylobacter Testing of Raw Meat and Poultry Products, CY 1998–2013. Available online at: https://www.fsis.usda.gov/wps/wcm/connect/885647f4-2568-48bf-ae5c4a0d8279f435/Progress-Report-Salmonella-Campylobacter-CY2013.pdf? MOD=AJPERES (Accessed January 22, 2019).
USDA-FSIS (2015). Progress Report on Salmonella and Campylobacter Testing of Raw Meat and Poultry Products, CY, 1998–2014. Available online at: https:// www.fsis.usda.gov/wps/portal/fsis/topics/regulatory-compliance/!ut/p/a0/04_ Sj9CPykssy0xPLMnMz0vMAfGjzOINAg3MDC2dDbwMDIHQ08842MTDy8_ YwMhYvyDbUREAhrzf7A!!/?1dmy¤t=true&urile=wcm%3Apath%3A %2Ffsis-content%2Finternet%2Fmain%2Ftopics%2Fdata-collection-andreports%2Fmicrobiology%2Fannual-progress-reports (Accessed January 22, 2019).
Wu, D., Alali, W. Q., Harrison, M. A., and Hofacre, C. L. (2014). Prevalence of salmonella in neck skin and bone of chickens. J. Food Prot. 77, 1193–1197. doi: 10.4315/0362-028X.JFP-14-006
Yang, H., Li, Y., and Johnson, M. G. (2001). Survival and death of Salmonella typhimurium and Campylobacter jejuni in processing water and on chicken skin during poultry scalding and chilling. J. Food Prot. 64, 770–776. doi: 10.4315/0362-028X-64.6.770










.jpg&w=3840&q=75)