The potential of supplemental enzymes in dairy and feedlot diets: impact of a protected fungal amylase preparation on ruminal fermentation and animal production
Introduction
The use of exogenous enzyme supplements for dairy and beef cattle has received much attention during the last several years. Research efforts have mainly focused on the application of fibrolytic enzymes as feed pre-treatment agents or as direct-fed supplements. Direct feeding of exogenous enzyme preparations is more practical and is supported by studies showing high stability of fungal carbohydrases in rumen fluid (Hristov et al., 1998; Morgavi et al., 2000) and positive production responses in ruminant animals (Howes et al., 1998; Lewis et al., 1999; Zinn and Salinas, 1999). In general, the animal production response to exogenous enzyme supplementation is not linear. Some researchers report a quadratic response to increasing concentrations of exogenous supplemental enzymes (Beauchemin et al., 1995; Sanchez et al., 1996; Lewis et al., 1999). These observations indicate that optimal enzyme concentrations are required to improve animal performance and it is possible to both under- and over-feed exogenous enzyme supplements.
Fiber digestion is usually considered a limiting factor in the overall digestion process in the rumen. For this reason, the majority of the studies performed on exogenous enzyme supplementation of ruminant diets concentrate on the use of preparations containing combinations of various fibrolytic activities. The mechanism by which exogenous fibrolytic enzymes enhance performance is related to their ability to enhance the digestibility of fiber in the feed. For example, direct-fed supplements containing fibrolytic activities can improve ruminal digestion of the neutral detergent fiber (NDF) fraction of the diet (Johnston and Shivas, 1999; Johnston, 2000). In most instances, the increase in NDF digestion results in greater dry matter (DM) intake that ultimately leads to improved animal performance (Zinn and Salinas, 1999; Lewis et al., 1999). However, in vitro studies suggest that some exogenous enzyme preparations also have the ability to alter microbial activities in the rumen (Dawson and Tricarico, 1999). This observation indicates that some enzyme supplements may have an impact on animal production by improving the efficiency of nutrient utilization without effectively increasing digestibility or DM intake.
Unlike fiber digestion, starch digestion is usually not considered to be limiting in the rumen. Moreover, rapid digestion of excessive amounts of starch in the rumen can cause ruminal acidosis and reduce intake and production (Owens et al., 1998). Some researchers have also suggested that starch digested post-ruminally may be used more efficiently by the animal than starch digested in the rumen (Owens et al., 1986; Nocek and Tamminga, 1991). However, no animal performance studies have clearly demonstrated that this is the case. In addition, the rate and extent of starch digestion in the rumen appear to have an impact on total tract starch digestibility and animal performance (Lykos et al., 1997).
Recent studies evaluating the effects of exogenous enzyme supplements for ruminants have rarely focused on starch digesting activities. Amylases were included in many of the early studies on exogenous enzyme supplements for ruminants (Burroughs et al., 1960; Ralston et al., 1962; Rust et al., 1963; van Welleghem et al., 1964; Perry et al., 1966). The inconsistent results and lack of detailed information on the specific activities and concentrations used in these studies do not provide conclusive evidence of a contribution of exogenous amylases to the overall effects of supplemental enzyme preparations on animal production. Ration formulation for high producing dairy cattle or rapidly growing beef cattle has long been fraught with the problems inherent with variations in fiber or starch digestibility. Research has shown that exogenous fibrolytic enzymes can be used to alleviate many problems associated with fiber digestibility. This success has raised hopes that a similar approach could be used to address problems with starch digestibility. For these reasons, we examined the effects of four doses of a proprietary fungal enzyme preparation containing amylase activity on milk production, milk composition and ruminal digestibility and fermentation in lactating Holstein cows.
Methods
A total of 24 multiparous lactating Holstein cows consuming a corn silage-ground corn based TMR (Table 1) were used in a replicated 4x4 Latin square design to examine the effects of four doses of amylase supplementation on milk production and composition. Ruminal parameters and ruminal digestibility of dry corn and corn silage were examined in four cows fitted with ruminal cannulas. The trial was conducted on a dairy farm in the Ottawa valley in Ontario. Cows were housed in tie stalls and milked twice daily. The diet consisted of a total mixed ration (Table 1) with a forage:concentrate ratio of approximately 40:60 (42:58 based on 22.37 kg DM intake/day) The average DIM at the start of the experiment was 117 days (SD=75). Experimental periods were 21 days. Milk production, milk composition, fermentation and digestibility were evaluated after a 14 day adaptation period to the experimental diets.
TREATMENTS
Treatments included supplementation of a proprietary fungal amylase preparation (Alltech Inc., Nicholasville, KY) at 0, 12, 24 and 36 g per cow per day. The four treatments were assigned to 20 intact and four ruminally cannulated cows based on days in milk and presence or absence of ruminal cannulas in a 4x4 Latin square design replicated six times.
Table 1. Chemical composition of the total mixed diet.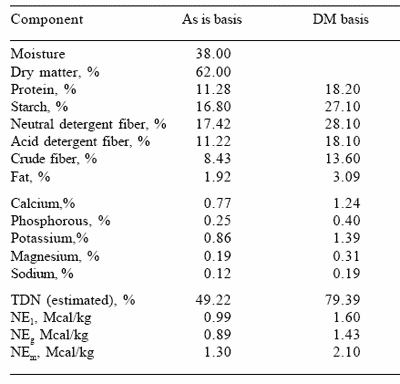
RESPONSE VARIABLES
Feed refusals and DM intake were recorded daily for each treatment group during the last week of each period. Milk production was recorded at each milking on two consecutive days during the last week of each period. Milk samples were collected at each milking and were sent to the DHIA Center in Guelph, ON for fat, protein and milk urea nitrogen (MUN) analyses. Blood samples were collected once during the last week of each treatment period and frozen for subsequent analysis of non-esterified fatty acids (NEFA), 3-D-hydroxybutyrate (BHBA) and glucose. Rumen fluid samples were collected at 9:00 a.m. and 12:00, 3:00 and 6:00 p.m. on two consecutive days during the last week of each treatment period for determination of ruminal volatile fatty acid (VFA) and ammonia concentrations. Ruminal NDF and starch digestibility of corn silage were estimated in each period using in situ bags on one cannulated animal in each treatment group. NDF and starch digestibility data collected from each animal on two consecutive days were considered as analytical replicates. Twelve in situ bags were placed in the rumen of each cannulated animal. Four bags were removed from the rumen after incubation for 3, 8 and 30 hrs. Bags were washed immediately after removal from the rumen and taken to the laboratory to be dried at 60°C. The dried residues in each bag were analyzed for NDF and starch content. Ruminal starch digestibility of 2000 and 2001 dry ground corn was estimated in situ during the last period of enzyme supplementation.
ANALYSIS
All data were analyzed as a one-way treatment classification using the general linear models procedure of SAS. Milk production, milk composition, and plasma metabolite data were analyzed as a replicated Latin square design. Squares were considered to be fixed effects. Periods and treatments were assumed to be independent and cows were nested within square. Ruminal fermentation data were analyzed as a Latin square-repeated measures design with enzyme treatment as the whole plot and sampling time as the subplot. The mean square for enzyme treatment within cow and period was used as the error term for evaluating the main effects of enzyme treatment. In situ digestibility, DMI and efficiency of milk production data were analyzed as a Latin square design. Orthogonal polynomials were used to partition linear, quadratic and cubic effects of enzyme supplementation.
Results
MILK YIELD AND COMPOSITION
Enzyme supplementation had a quadratic effect on milk production (Figure 1) and production of 3.5% fat-corrected milk (FCM) (Figure 2) without a significant increase in DM intake. Milk and 3.5% FCM yields were 1.5 and 1.8 l greater (P<0.05) in animals receiving 12 g enzyme per day than in animals receiving no enzyme supplement. The effects of enzyme supplementation on milk and 3.5% FCM yields decreased as supplemental enzyme concentrations increased above 12 g/day. Enzyme supplementation did not affect the percentage of fat and protein in milk. Therefore, milk fat and protein yields were greater (P<0.05) in animals receiving 12 g enzyme per day than in animals receiving no enzyme supplement (Figures 3 and 4, respectively).
In addition, enzyme supplementation had a significant cubic effect on MUN (Figure 5). Supplementation with 12 g enzyme per cow per day resulted in lower MUN concentrations than in any other treatment.
Enzyme supplementation did not affect the in situ starch or NDF digestibility of corn silage (data not shown). However, starch digestibility of dry ground corn was greater (P<0.05) after in situ incubation for 24 hrs in the rumen of animals receiving 12 g enzyme per day than in the rumen of animals receiving no enzyme or higher enzyme concentrations (Table 2). In addition, a significant year effect was observed on starch digestibility of dry ground corn.
Enzyme supplementation did not affect the concentration of total VFAs in the rumen (Table 3). However, enzyme supplementation had linear, quadratic and cubic effects on the proportions of acetate, propionate and butyrate produced. The addition of 12 g enzyme per day reduced (P<0.01) the proportion of propionate by 9.1 % and increased (P<0.01) the proportions of acetate and butyrate by 1.5 and 9.3 %, respectively. The acetate to propionate ratio (A:P) was greatest in animals fed 24 g enzyme per day, intermediate in animals fed 12 or 36 g enzyme per day and lowest in animals receiving no supplemental enzyme. The addition of supplemental enzyme did not affect the concentration of ammonia in the rumen.
Figure 1. Effects of enzyme supplementation on milk production.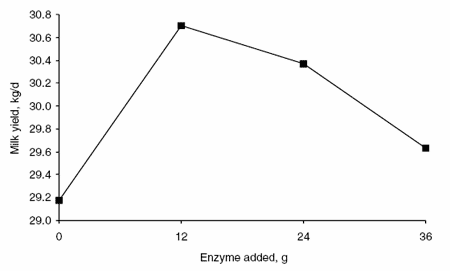
Figure 2. Effects of enzyme supplementation on production of 3.5% FCM.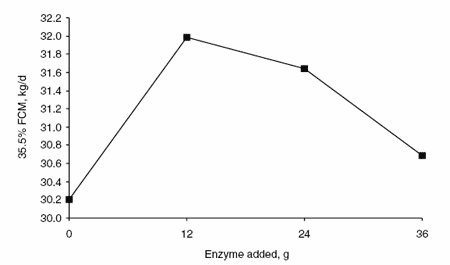
Figure 3. Effects of enzyme supplementation on milk fat production.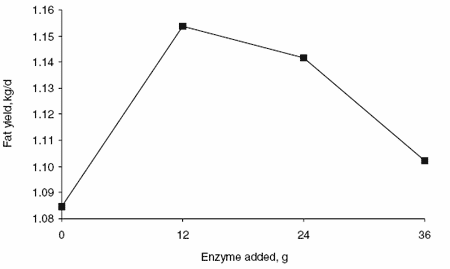
Figure 4. Effects of enzyme supplementation on milk protein production.
Figure 5. Effects of enzyme supplementation on MUN concentration.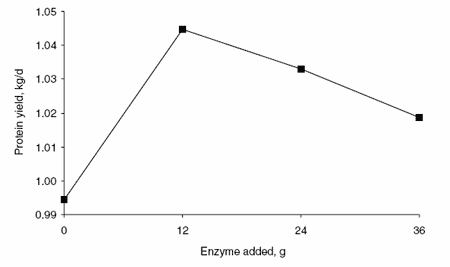
Enzyme supplementation had a quadratic effect on the concentration of BHBA (Figure 6). The concentration of BHB was greatest in animals fed 12 or 24 g enzyme per day, intermediate in animals fed 36 g enzyme per day and lowest in animals fed no supplemental enzyme. The addition of supplemental enzyme had a linear affect on plasma concentrations of NEFA (Figure 7) and glucose (Figure 8). The plasma NEFA concentrations were greater (P<0.05) in animals fed 24 g supplemental enzyme than in animals receiving no enzyme. Glucose concentrations in blood were greater (P<0.05) in animals fed 0 or 12 g enzyme per day than in animals receiving 24 or 36 g.
Table 2. Effects of increasing concentrations of supplemental enzyme and crop year on starch digestibility of dry ground corn.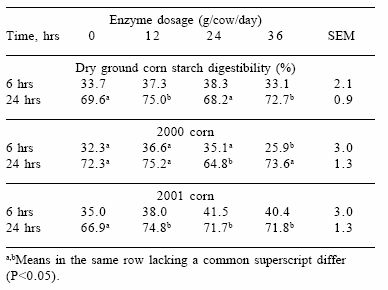
Conclusions and implications
We have shown in this study that addition of supplemental amylase to a corn silage/ground cornbased diet can dramatically improve milk production in dairy cows. The enzyme supplement did not alter the rate or extent of starch digestion of the corn silage, but increased the extent of starch digestion of the dry ground corn. The enzyme effect on ground corn digestibility was year dependent, and was most pronounced on the 2001 crop, which had a higher bushel weight than the 2000 crop.
Table 3. Effects of increasing supplemental enzyme concentrations on ruminal VFA and ammonia concentrations in lactating Holstein cows.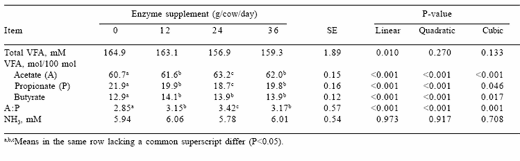
Figure 6. Effects of enzyme supplementation on blood BHBA concentration.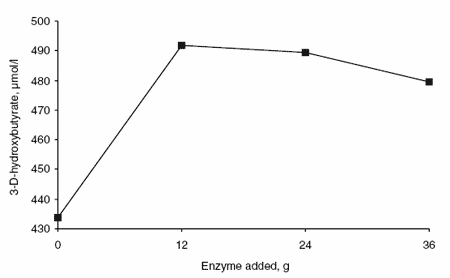
Figure 7. Effects of enzyme supplementation on blood NEFA concentration.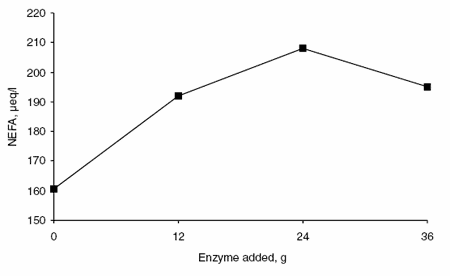
Figure 8. Effects of enzyme supplementation on blood glucose concentration.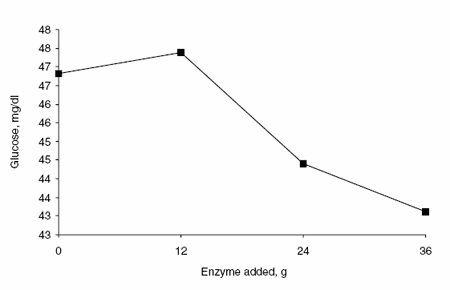
The magnitude of the increase in starch digestibility of the grain portion of the diet is by itself too small to produce the large increase in milk production observed in this study. The majority of the increase in milk production can probably be attributed to the effects of the enzyme supplement on proportions of ruminal VFAs and the effects on nutrient utilization that resulted from those changes in ruminal fermentation. The supplemental enzyme increased the concentration of butyrate at the expense of propionate, regardless of the concentration of enzyme added. These observations suggest that the mechanism by which supplemental amylase enhances butyrate production in the rumen is probably by modifying the ruminal microbial population. Our hypothesis is that addition of low concentrations of supplemental amylase will continually provide low concentrations of one or more soluble sugars that will give a competitive advantage to butyrate producing organisms (e.g. Butyrivibrio fibrisolvens) over lactic acidproducing starch digesters (e.g. Streptococcus bovis). This competitive advantage is probably directly related to the corresponding affinities that the various ruminal microorganisms have for the different soluble sugars (Russell and Baldwin, 1978). Therefore, this competitive advantage will only be attained at low exogenous enzyme concentrations and will be disrupted when high concentrations of supplemental enzymes are fed.
The increases in ruminal production of butyrate and the reductions in propionate production observed in this study appeared to have a large impact on nutrient utilization by the lactating cows.
Approximately 74 to 90% of the butyrate produced in the rumen is metabolized by the rumen epithelial tissue (Remond et al., 1995) with a large portion converted to ketone bodies, mainly BHBA. It is likely that the higher concentrations of BHBA in blood obtained with enzyme addition result from an increase in the amount of butyrate metabolized by the rumen epithelium in the presence of supplemental enzyme. Ruminants can use BHBA as an energy source for skeletal and cardiac muscle tissue and as a precursor for fatty acid synthesis in adipose tissue and the mammary gland (Church, 1988). Therefore, the increased concentration of BHBA in blood represents an additional source of energy for milk synthesis in enzyme-supplemented animals. It is of particular interest to note that the best response in milk production was observed with the lowest concentration of supplemental amylase. This is probably due to the fact that blood glucose concentration was lower in animals receiving 24 or 36 g per day but not in animals receiving 12 g per day. Therefore, cows receiving the lowest concentration of supplemental enzyme had more available energy, as indicated by the sum of blood BHBA and glucose, than cows receiving 0, 24 or 36 g.
J.M. TRICARICO1, J.D. JOHNSTON2 AND K.A. DAWSON1
1 Alltech North American Biosciences Center., Nicholasville, KY, USA
2 Ritchie Feed & Seed, Ottawa, ON, Canada.
References
Beauchemin, K.A., L.M.Rode and V.J.H. Sewalt. 1995. Fibrolytic enzymes increase fiber digestibility and growth rate of steers fed dry forages. Canadian J. of Anim. Sci. 75:641-644.
Burroughs, W., W. Woods, S.A. Ewing, J. Greig and B. Theurer. 1960. Enzyme additions to fattening cattle rations. J. of Anim. Sci. 19:458-464.
Church D. C. 1988. The ruminant animal. Digestive physiology and nutrition. (D.C. Church, ed.). Englewood Cliffs, NJ.
Dawson, K.A. and J.M. Tricarico. 1999. The use of exogenous fibrolytic enzymes to enhance microbial activities in the rumen and the performance of ruminant animals. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 15th Annual Symposium (T.P. Lyons and K.A. Jacques, eds), Nottingham University Press, Nottingham, UK, pp. 303-312.
Howes, D., J.M. Tricarico, K.A. Dawson and P. Karnezos. 1998. Fibrozyme, the first protected enzyme for ruminants: improving fiber digestion and animal performance. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 14th Annual Symposium (T.P. Lyons and K.A. Jacques, eds), Nottingham University Press, Nottingham, UK, pp. 393-403.
Hristov, A.N., T.A. McAllister and K-J. Cheng. 1998. Stability of exogenous polysaccharidedegrading enzymes in the rumen. Animal Feed Sci. and Tech. 76:161-168.
Johnston, J. D. 2000. Fibrozyme and in vitro NDF response: moving from theory to practical commercial reality. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 16th Annual Symposium (T.P. Lyons and K.A. Jacques, eds), Nottingham University Press, Nottingham, UK, pp. 487-500.
Johnston, J.D. and I.C. Shivas. 1999. Formulating the fourth diet. In: Biotechnology in the Feed Industry. Proceedings of Alltech’s 15th Annual Symposium (T.P. Lyons and K.A. Jacques, eds), Nottingham University Press, Nottingham, UK, pp. 321-341.
Lewis, G.E.,W.K. Sanchez, C.W. Hunt, M.A. Guy, G.T. Pritchard, B.I. Swanson and R.J. Treacher. 1999. Effect of direct-fed fibrolytic enzymes on the lactational performance of dairy cows. J. of Dairy Sci. 82:611-617.
Lykos, T., G.A. Varga and D. Casper. 1997. Varying degradation rates of total nonstructural carbohydrates: Effects on ruminal fermentation, blood metabolites, and milk production and composition in high producing Holstein cows. J. of Dairy Sci. 80:3341-3355.
Morgavi, D. P., C.J. Newbold, D.E. Beever and R.J. Wallace. 2000. Stability and stabilization of potential feed additive enzymes in rumen fluid. Enzyme and Microbial Tech. 26:171-177.
Nocek, J.E. and S. Tamminga 1991. Site of digestion of starch in the gastrointestinal tract of dairy cows and its effect on milk yield and composition. J. of Dairy Sci. 74:3598-629.
Owens, F.N., R.A. Zinn and Y.K. Kim. 1986. Limits to starch digestion in the ruminant small intestine. J. Anim. Sci. 63:1634-48.
Owens, F.N., D.S. Secrist, W.J. Hill and D.R. Gill. 1998. Acidosis in cattle: a review. J. of Anim. Sci. 76:275-86.
Perry, T.W., E.D. Purkhiser and W.M. Beeson. 1966. Effects of supplemental enzymes on nitrogen balance, digestibility of energy and nutrients and on growth and feed efficiency of cattle. J. Anim. Sci. 25:760-764.
Ralston, A.T., D.C. Church and J.E. Oldfield. 1962. Effect of enzymes on digestibility of low quality roughage. J. Anim. Sci. 21:306-308.
Remond D., Ortigues I. and J. P. Jouany. 1995. Energy substrates for the rumen epithelium. Proceedings of the Nutrition Society. 54:95-105.
Russell, J.B. and R.L. Baldwin. 1978. Substrate preferences in rumen bacteria: Evidence of catabolite regulatory mechanisms. Applied and Environmental Microbiology 36:319-329.
Rust, J.W., N.L. Jacobson and A.D. McGilliard. 1963. Supplementation of dairy calf diets with enzymes. I. Effect on rate of growth. J. Anim. Sci. 22:1104-1108.
Sanchez, W.K., C.W. Hunt, M.A. Guy, G.T. Pritchard, B. Swanson, T. Warner and R.J. Treacher. 1996. Effect of fibrolytic enzymes on lactational performance of dairy cows. Proceedings of the American Dairy Science Association. Corvallis, OR.
van Walleghem, P.A., C.B. Ammerma, C.F. Chicco, J.E. Moore and L.R. Arrington. 1964. Enzyme supplements and digestibility of protein and energy in rations high in dried citrus pulp. J. of Anim. Sci. 23:960-962.
Zinn, R.A. and J. Salinas. 1999. Influence of Fibrozyme on digestive function and growth performance of feedlot steers fed a 78% concentrate growing diet. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 15th Annual Symposium (T.P. Lyons and K.A. Jacques, eds), Nottingham University Press, Nottingham, UK, pp. 313-319.

Very important point. Especially it could be used together with "Cutinase" effect of fibrolitic and amylolytic enzymes from Asp. spp. biomass. We have check them both in Fibrase (Italy).





.jpg&w=3840&q=75)