1. Introduction
Newcastle disease (ND), caused by an avian paramyxovirus ser otype 1 (now designated as orthoavulavirus 1) [1], and infectious bronchitis (IB) caused by an avian coronavirus [2], are known to give rise to severe diseases in chickens, with disastrous economic losses and welfare concerns [3,4]. ND and IB have been important and common chicken diseases worldwide since 1926 and 1936 respectively. In the last three decades, publications from both our and other laboratories have highlighted the importance of aMPV [1], which causes a drop in egg production and quality, and swollen head syndrome in chickens [5,6]. Infectious bronchitis virus (IBV), avian metapneumovirus (aMPV) and Newcastle Disease virus (NDV) are respiratory RNA viruses that primarily infect the tra cheal epithelium of chickens [7].
A numberof countries have introduced the use of live and inac tivated NDV, IBV and aMPV vaccines [8]. Simultaneous vaccination with multiple live viral vaccines is carried out in poultry for a num ber of reasons. These include the avoidance of inducing stress in chicks through excessive handling, and to reduce vaccination costs [9]. Traditionally, NDV and single IBV Massachusetts live vaccines are given to day-old chicks at the hatchery or in the farms, mostly by spray. However, as the disease and losses caused by variant IBVs increase [10–12], growing numbers of producers have been includ ing a 793Bvaccine in the day-old vaccination programme. All three diseases (IBV, aMPV and NDV) are now routinely controlled in the poultry industry through the use of live attenuated vaccines [13,14], with long lasting antibody titres recorded for inactivated NDV vaccinations [15].
Reports since the 19600s have described how IBV vaccines can impair the efficacy of NDV vaccines [16,17], potentially leading to production losses [18]. Gelb et al. (2007) demonstrated how NDV immunity decreased when combined with an IBV Arkansas vaccine in broiler chickens [7]. With the introduction of aMPV in 19900s, the possibility of heterologous co-vaccination of NDV + aMPV and IBV+aMPV at day-old was investigated [19–23]. These publications showed that simultaneous administration of aMPVvaccine alongside NDV or IBV vaccine were compatible, with protection against virulent IBV, NDV or aMPV remaining uncom promised. However, in these studies, only a single vaccine strain of IBV (Massachusetts) was used. With the emergence of variant IBVs, inclusion of a variant IBV alongside Massachusetts for day old vaccination has become a common practice [19,24,25].
Recent work has shown that IBV vaccines can undergo genetic mutations following inoculation into the chicken host [26,27].As persistent mutations can alter strain characteristics [28], it is preferable to monitor potential changes. Furthermore, it will be beneficial to establish if simultaneous vaccine administration has any role in nucleotide mutations occurring in the IBV S1 gene and subsequent amino acid changes. In contrast to IBV vaccine strains, prolonged persistence or genetic mutations in NDV or aMPV vaccine strains have not been reported in chickens.
To date, the impact of co-administration of live NDV or aMPV alongside classical and variant live IBV vaccines in IBV-aMPV NDV maternal antibody positive birds has not been investigated. With the increased use of co-vaccination strategies in commercial broiler chicks, it is important to understand how such vaccine combinations may impact protection against IBV, aMPV or NDV challenges. This study reports the results of two experiments where live IBV vaccines of Massachusetts and 793B were co administered alongside a live aMPV subtype B or live Newcastle disease vaccine in commercial broiler chicks.
2. Materials and methods
Chicks: Day-old commercial broiler chicks were obtained from a commercial hatchery, and raised in the University of Liverpool isolation facilities. Chicks were reared on deep litter with water and feed provided ad libitum. No in-feed or in-water antibiotics wereusedthroughout the study. All experimental procedures were conducted according to UK legislation on the use of animals for experiments, and were granted ethical approval by the University of Liverpool ethics committee.
Vaccine strains: Commercially available live IBV Mass type strain H120 (Bioral, Boehringer Ingelheim, Lyon, France) and IBV variant 793B type strain CR88 (Gallivac IB88, Boehringer Ingel heim, Lyon, France), NDV VG/GA-Avinew strain (Avinew NeO, Boehringer Ingelheim, Lyon, France) and aMPV subtype B (Nemo vac , Boehringer Ingelheim, Lyon, France) vaccines were adminis tered during this study. All vaccines were prepared prior to administration using sterile distilled water, to allow for 100 mlof vaccine per chick to be administered according to manufacturer’s instructions [5,19].
Challenge strains: Virulent virus strains of IBV (M41, 105.0 Cil iostatic Dose (CD)50/ml; QX, 106.0 CD50/ml) and aMPV (subtype B; 104.51 CD50/ml) were used for challenge. All challenge strains have been propagated in our laboratory and used in several previous studies [19,23,29]. PCR and bacterial culturing confirmed an absence of NDV, aMPV, avian influenza virus (AIV), infectious laryngotracheitis virus (ILTV), infectious bursal disease virus (IBDV), fowl adenovirus (FAdV) and mycoplasmas in all inocula [30–35].
2.1. Experimental design
Experiment 1: Chicks were randomly divided into four groups (35 chicks per group) and kept in separate isolation units. At day-old, each chick was inoculated via oculo (50ul) and nasal (50ul) routes with the following; VG/GA-Avinew (Group A), Mass + 793B (Group B), VG/GA Avinew + Mass + 793B (Group C) and sterile H₂O (Group D). Chicks were observed daily for clinical signs [36]; Mild = Coughing, head shaking and depression of short duration, Severe = gasping, coughing and depression, accompanied by ruffled feathers. Mortality rates and lesions following necropsy were recorded.
At 21dpv, chicks from each group were separated into two fur ther groups (A1, A2, B1, B2, C1, C2, D1 and D2), with 10 chicks per sub-group. Chicks from A1, B1, C1 and D1 were challenged with 100 ml of virulent M41, and chicks from A2, B2, C2 and D2 were challenged with 100 ml of virulent QX via oculo-nasal route. Chicks were observed daily for clinical signs as previously described, with mortality rates and post mortem lesions recorded.
At 0, 3, 7, 10, 14 and 21dpv, oropharyngeal (OP) swabs were collected from five chicks in each group. At 7, 14, 21 dpv, five tra chea and kidney samples were collected from each group. At 5 dpc, f ive trachea and kidney samples were collected and processed as previously described [26]. Blood was collected from ten chicks at 0 dpv and eight chicks at 21dpv for the detection of IBV and NDV antibodies.
Experiment 2: Day-old broiler chicks were randomly divided into four groups and kept in separate isolation units, with 30 chicks per group. At day-old, each chick was inoculated via oculo (50 ml) and nasal (50ml) routes with the following; aMPV subtype B (Group A), Mass + 793B (Group B), aMPV subtype B+Mass+793B (Group C) and sterile H2O (Group D). Chicks were observed daily for clinical signs as described above.
At 21days post vaccination (dpv), chicks from each group (A-D) were further split into twelve groups (A1-A3, B1-B3, C-C3, D1-D3), with five chicks per group. Chicks from A1, B1, C1 and D1 were challenged with 100 ml of virulent M41, and chicks in the A2, B2, C2 and D2 groups were challenged with 100ml of virulent aMPV B via oculo-nasal route. Chicks from A3, B3, C3 and D3 were sham-inoculated with sterile distilled water. All birds were observed daily for clinical signs, with IBV challenge groups scored as above, and aMPV challenge groups scored as follows: 0 =No nasal exudate, 1 = Mild exudate, 2 = Unilateral excessive exudate, 3 =Bilateral excessive exudate [5].
At 0, 3, 7, 10, 14 and 21dpv, oropharyngeal (OP) swabs were collected from five chicks in each group. At 7, 14, 21 dpv five tra chea and kidney samples were collected. At 5dpc, five trachea and kidney samples were collected and processed as previously described [26]. Blood was collected from ten chicks at 0 dpv and eight chicks at 21dpv for the detection of IBV and aMPV antibodies.
Evaluation of tracheal protection following M41 challenge: At 5days post challenge (dpc), five chicks from each group were humanely killed by wing vein injection of sodium pentobarbital. Tracheas were removed from each chick and processed for ciliary and percentage protection assessment as previously described [14,19]. Briefly, 10 rings from each bird were examined, with a maximum possible ciliary score of 40 indicating no protection (no cilia beating). The mean ciliary score for each bird was calcu lated and percentage protection for each group was calculated.
Measurement of antibody levels: Commercial ELISA kits [IBV (IDEXX, Maine, United States) and aMPV (BioChek, ER Reeuwijk, Netherlands)] were used according to manufacturer’s instructions, with a titre result higher than 396 (IBV) or 1655 (aMPV) indicating a positive protection titre. Haemagluttination inhibition (HI) was utilised to determine IBV M41 and NDV protection status as previ ously described [37].
IBV, NDV and aMPV detection by RT-PCR: Swabs were pooled and dipped as a single sample per group, per day in a sterile bijou containing 1.5 ml of Eagles serum-free minimum essential med ium with glutamine, streptomycin [50mg/ml] and penicillin [50 IU/ml], which was stored at 70 C until required.
RNA was extracted from the swab and tissue samples using the phenol chloroform method [38] and subjected to RT-PCR for IBV, NDV and aMPV [32,39,40]. Positive amplicons were purified and sent for bi-directional commercial Sanger sequencing (Source BioS cience, Nottingham, UK).
IBV and aMPV viral load in trachea and kidney: Total RNA was extracted from each group’s five trachea and kidney samples using the RNeasy Plus Mini Kit (Qiagen, Manchester, UK). Quantifi cation of viral RNA was determined by quantitative RT-PCR (qRT PCR) for both IBV and aMPV [41,42]. The IBV qRT-PCR was per formed using the Qiagen OneStep RT-PCR kit and 40ng of total RNAperreaction. The aMPV assay was performed using the Qiagen Quantitect 2xRT-PCR no ROX master mix and 40ng of total RNA per reaction. Ct values were converted to log2 of Relative Equiva lent Units (REU) of viral RNA, using previously determined stan dard curves for IBV and aMPV [25].
Sequencing analysis: IBV partial-S1 gene sequences were trimmed to 320bp, analysed in ChromasPRO v1.7.3 (http://tech nelysium.com.au/), and BLAST searches were conducted to confirm the isolate identity. Alignments were carried out in MEGA6 [43], using Clustal W [44]. Single nucleotide polymorphism (SNP) and insertion and deletion (indel) detection was carried out in MEGA6, following alignment to sequenced vaccine strains. SNPs were char acterised as non-synonymous (ns) if it led to an amino acid change, and synonymous (s) if it led to no amino acid changes. Insertions and deletions (indels) were defined as the insertion or deletion of a nucleotide that altered the sequence length. The ds/dns ratio, to identify positive or purifying selection pressure for each group was calculated per day. This was done using the Nei-Gojobori method [45] where < 1.0 indicated the recovered strains to be under positive selection pressure, and > 1.0 indicated strains to be under purifying selection [26,46]. Positive or purifying selection was considered significant at P < 0.05. Translated amino acid vari ations were also identified and variations that resulted in a change in hydrophobicity were noted according to the Kyte and Doolittle scale [47].
Statistical analysis: Statistical analysis was conducted using one-way analysis of variance (ANOVA), followed by Turkey’s test to examine the differences between pairs of means. Differences were considered to be significant when p < 0.05. Analysis was car ried out using SPSS Statistics 22.
3. Results
3.1. Experiment 1. Simultaneous administration of NDV alongside IBV Mass + 793B vaccines
3.1.1. Clinical signs
Following day-old vaccination, in both IBV-vaccinated groups, mild respiratory signs were recorded from 2 to 14 dpv. After M41 or QX challenge, groups that received no IBV vaccination had mild signs throughout the experiment. The IBV-vaccinated groups, which were M41-challenged or QX-challenged showed mild signs for a total of two days, at 3–4 dpc.
3.1.2. IBV protection by cilia-stopping test
Ciliostasis results showed that all IBV-vaccinated groups (B1, B2, C1, C2) have high protection against both IBV challenge strains, ranging from 94 to 99% (C1– 94%; C2– 96.76%; B1– 99%; B2 98.56%).
3.1.3. NDV protective haemagluttination inhibition antibody titres
The NDV HI titres in groups that had not received an NDV vac cine were below 2 log2 (Table 1), whereas the groups that received NDVvaccine singly or in combination with IBV were 6.63 and 5.63 respectively. A significant increase (p < 0.05) in titre was seen for Group A (NDV vaccinated) when compared to Group C (combined vaccination) at 21 dpv.
3.1.4. IBV haemagluttination inhibition and ELISA antibody titres
When the same sera were assayed against IBV HI antibodies, IBV-vaccinated groups showed an increase in HI log2 titre at 21 dpv when compared to 0dpv (Table 1). Furthermore, there was a significant increase (p < 0.05) in titre for both Groups A and B whencompared to Group C for the IBV (4/91) antigen. There was astronger response towards the 4/91 compared to the M41 HI antigen for all IBV vaccinated groups (Group B– 6.75 and 4.00; Group C– 5.88 and 5.63). The mean IBV ELISA antibody titre at day-old was 838.38 (±87), indicating the presence of maternally derived antibodies (MDA). By 21 dpv, antibody levels declined to below the detectable titre (3 9 6) in all groups (Group A– 23.3, Group B– 89, Group C– 71.8 and Group D– 27.7). Despite being below detectable levels, the two groups receiving IBV vaccines (B and C) had higher antibody titres compared to the IBV unvaccinated groups (A and D).
3.1.5. Molecular detection of IBV and NDV in OP swabs
IBV RNA was detected at all sampling time points (3, 7, 10, 14 and 21dpv) in Groups B and C, and was absent from the IBV unvaccinated groups (A and D). The Mass-type vaccine was detected up until 10 dpv in Group B and 14dpv in Group C. After this time, only the 793B-type vaccine was identified in all subse quent samples in both IBV-vaccinated groups. All Mass-type iden tified strains had partial-S1 nucleotide similarity to the original vaccine, ranging from 92.97 to 99.74%, whereas the 793B-type had 93.72–99.47% relatedness to the CR88 vaccine strain. NDV was not detected at any time.
Total SNP counts remained low, with the exception of samples from Groups B and C at 14dpv, which contained greater total SNP counts (n=21) (Table 2). This translated to 14 amino acid (AA) changes in Group B and 16 in Group C. All changes in 793B like samples were from hydrophobic to hydrophilic properties, and the majority (75%) of changes in the Mass-like samples were hydrophilic to hydrophobic. The average ds/dns ratio was 1.96 for 793B strains and 2.81 for Mass strains, indicating that the Mass genotype was significantly under purifying selection pressure (p < 0.05). No indels were present for any of the sequences analysed.
3.1.6. Molecular detection of IBV and NDV from trachea and kidney tissues
Up to 21dpv: IBV vaccinal strains were recovered from both tra chea and kidney tissue in Group B at 7 and 14dpv, whereas all sampling days were positive in Group C (Table 3). No trachea sam ples were IBV-positive at 21 dpv and no Mass-type strains were recovered in the kidney past 7 dpv. We did not detect NDV in any samples. Sequence similarity to the IBV vaccine strains were between 98 and 100%, with a single 793B exception of 96% at 21 dpv. The majority of strains recovered from the kidney and tra chea (84.62%) had a ds/dns ratio of over 1.00, demonstrating that the majority of partial S1 nucleotide variations had no effect on the translated amino acid composition. The exceptions were the two 793B-like strains from Group C kidney samples at 21 dpv. This group had an average of five non-synonymous changes per strain, which translated to three hydrophobicity changes within all 13 recovered strains.
IBV M41 challenge at 5 dpc: IBV was detected in the trachea of Group D1 (Non-vaccinated), and in the trachea and kidney of Group A1 (NDV vaccinated) (Table 4). IBV vaccine strains were detected in Groups B1 (Mass vaccinated 99% similarity) and C1 (793B vaccinated 96% similarity). The majority of variations led
Table 1
Experiment 1: Anti-NDV and IBV (4/91 and M41) HI geometric mean titres at 21 dpv, prior to challenge against virulent IBVs. Standard error margins are shown in brackets. Significant differences between groups (p < 0.05) are indicated with different letters.
Table 2
Nucleotide and amino acid variations in IBV strains recovered from pooled OP swabs. No IBV positive samples were detected in NDV vaccinated Group A and control Group D.
Table 3
Nucleotide and amino acid variations in IBV strains recovered from trachea and kidney tissues post vaccination. No IBV positive samples were detected in NDV vaccinated Group A and control Group D.
Table 4
Experiment 1: Nucleotide and amino acid variations in IBV strains recovered from trachea and kidney tissue at 5 days post M41 and QX challenge
to changes in the translated amino acids (n = 14; average dS/dN ratio = 0.85), however only one change in hydrophobic properties (hydrophilic to hydrophobic) was identified from a sample identified as highly similar to the virulent strain (Group A1; trachea).
IBV QX challenge at 5 dpc: Virulent QX strains were detected in the trachea and kidney of Group D2 (Non-vaccinated), with 98–99% nucleotide similarity to the virulent strain. However, only tracheal samples were IBV-positive for Group A2 (NDV-vaccinated) (Table 4). Vaccinal strains were detected in Groups B2 (793B; 98–99% similarity) and C2 (793B and Mass; 99% similarity). The minority of nucleotide variations caused an amino acid change (n = 29; average dS/dN ratio = 0.97), which resulted in seven hydrophobicity changes (hydrophilic to hydrophobic = 4; hydrophobic to hydrophilic = 3). Vaccine strains were recovered from kidney samples in Groups B and C, with the majority of changes in 793B-like samples being non-synonymous. From a total of 18 amino acid changes, only six caused a change in hydrophobicity.
3.1.7. IBV viral load in tracheal and kidney tissue
In the IBV vaccinated tracheal samples, viral load reduced from 7 dpv to 14 dpv (Group B = 1.49 to 0 log REU; Group C = 0.97 to 0.29 log REU), however IBV presence were significantly higher (p < 0.05) at 21 dpv for both groups (Group B = 2.92; Group C = 1.55) when compared to 7 dpv (Fig. 1). At both 7 and 21 dpv, there was a significantly higher tracheal viral load in the IBV vaccination group when compared to the NDV + IBV vaccine group. Post challenge, high viral loads were present in the trachea (Fig. 2) for Groups A1 (2.21 log REU; NDV vaccinated, M41 challenge), A2 (2.46 log REU; NDV vaccinated, QX challenge), D1 (2.83 log REU; non-vaccinated, M41 challenge) and D2 (2.49 log REU; non-vaccinated, QX challenge).
For the kidney samples at 14 and 21 dpv, Groups B (IBV vaccinated) and C (NDV + IBV vaccinated) increased from 14 to 21 dpv (0.81 to 3.13 log REU and 1.15 to 1.42 log REU respectively) (Fig. 1). Post challenge (Fig. 2), significantly lower viral loads were present in non-IBV vaccinated groups (A1 – 0.44 log REU; A2 – 0.25 log REU; C1 – 0.1 log REU; D1 – 0.18 log REU) compared to IBV vaccinated groups (B1 – 2.15 log REU; B2 – 1.22 log REU; C2 – 2.62 log REU). Following M41 challenge, viral load did not change in Group B (IBV vaccinated). The virulent QX strain was detected at a greater level in the control group (Group D) when compared to the M41 challenge control group. However, the viral load in both NDV vaccinated groups (A1 and A2) remained low (< 0.45 log REU) following challenge.
3.2. Experiment 2. Simultaneous administration of aMPV alongside IBV Mass + 793B vaccines
3.2.1. Clinical signs
Up to 21 dpv: All IBV vaccinated groups demonstrated mild clinical signs (prominent snicking and sneezing), starting at 6 and 7 dpv (Group B - Mass + 793B; Group C - aMPV + Mass + 793B respectively), and continuing up to 15 dpv. Group A (aMPV vaccinated) and Group D (Control) showed no clinical signs. No groups presented with moderate clinical signs at any point.
Up to 5 dpc: Following IBV M41 challenge, Groups A1 (aMPV vaccinated) and D1 (Control) showed signs from 1 dpc to 5 and 6 dpc respectively. Mild signs in Groups B1 and C1 subsided after 2 dpc. For aMPV challenge, greater clinical signs were observed in non-vaccinated birds (Group D2), or birds only receiving the IBV vaccines (Group B2), compared to the group receiving the combined vaccination (Group C2). All signs were cleared from aMPV challenge groups by 7 dpc. Groups receiving no challenge virus (A3, B3, C3 and D3) were absent of clinical signs.
3.2.2. Evaluation of IBV protection by cilia-stopping test
All IBV vaccinated groups showed high protection against the M41 challenge strains (91.12–97.84% protection score). The combined groups (C1 and C3) demonstrated a similar level of protection percentage (91.12 and 97.84 respectively) when compared to the single vaccination groups (B1 and B3, both 97.84%).
3.2.3. Anti-aMPV antibody titres by ELISA
The mean serum aMPV antibody titre at 0 dpv was 7217.54 (±572.83), indicating the presence of maternally derived antibodies (MDA) against aMPV (Fig. 3). By 21 dpv, antibody levels declined to below the detectable titre for the IBV vaccinated group (Group B). However, the aMPV vaccinated and combined groups remained at positive titres (3074 and 1668.78 respectively). By 10 dpc, only Groups B1, B3, D1 and D3 showed antibody levels below the detectable limit. Following aMPV challenge, Groups A2 (aMPV vaccinated) and B2 (IBV vaccinated) presented with higher titres (6367.4 and 4543.57 respectively) when compared with Group C2, which received the combined aMPV + IBV vaccination (3509.67). The aMPV vaccinated-aMPV challenged group (A2), and the combined vaccinated-aMPV challenged group (C2) had significantly higher (p < 0.05) titres compared to the non-challenged subgroups A3 and C3.
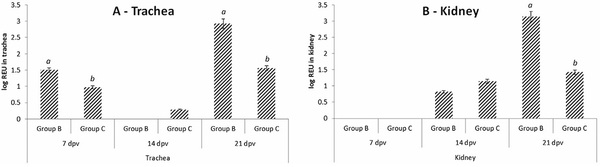
Fig. 1. Experiment 1: Quantification of IBV viral load in the trachea and kidney at 21 dpv following vaccination with either (Group B) H120 + CR88 or (Group C) VG/GA Avinew + H120 + CR88. Data is presented ± standard error margins (SEM). Significance testing carried out within sampling point of the same tissue type, with differences between groups (p < 0.05) labelled with different letters.
Fig.2. Experiment 1: Quantification of IBV viral load in the trachea and kidney at 5 dpc following challenge by either (A) virulent IBV M41 or (B) virulent IBV QX. Data is presented ± standard error margins (SEM). Significance testing carried out between groups of the same tissue type, with differences between groups (p < 0.05) labelled with different letters.
Fig. 3. Experiment 2: Quantification of aMPV antibodies at 0 and 21 dpv, and 10 dpc. Groups were regarded as positive at a titre of at least 1655 (indicated by a dashed line). Groups are indicated as follows: A = aMPV vaccinated, B = IBV vaccinated, C = Combined vaccinated, D = Unvaccinated; 1 = IBV Challenged, 2 = aMPV Challenged, 3 = Unchallenged. Data is presented ± standard error margins (SEM). Significant differences between groups of the same tissue type (p < 0.05) labelled with different letters.
3.2.4. Molecular detection of IBV and aMPV in OP swabs
We were able to detect both IBV and aMPV as early as 3 dpv. However, while IBV was identified until 21 dpv, we did not detect aMPV beyond 7 dpv in either single or combined vaccine groups. Partial S1 sequencing of IBV positive samples demonstrated a high similarity to the original inoculum, with identities ranging from 97.22 to 99.42% in Group B to 98.21–99.54% in Group C. No Mass-types were identified after 10 dpv in either group. On aver
age, samples contained 7.3 SNPs per sampling time, with only two OP samples having a higher than average SNP count (Group B [7 dpv] = 24 and Group C [14 dpv] = 26).
The dS/dN ratio was an average of 3.38 for the Mass-like strains and 1.46 for the 793B samples (Table 2). The two 793B-positive samples with greater than average SNP counts also showed an increase in amino acid changes, and were the only samples with hydrophobicity changes, with the majority (n = 4/6) being a change from hydrophilic to hydrophobic.
3.2.5. Molecular detection of IBV and aMPV from trachea and kidney tissue
Up to 21 dpv: IBV vaccinal strains were recovered from Groups B and C at all sampling points, with the exception of the kidney at 21 dpv (Table 3). All detected IBVs had a high partial-S1 nucleotide similarity with the initial vaccine strain (over 99.1%), with only a single non-synonymous change at all time points for Groups B and C. The tracheal sample from Group C at 21 dpv had 20 SNPs, which translated into nine amino acid variations (containing a single hydrophobic to hydrophilic change). The aMPV vaccine was detected up until 21 dpv in tracheal samples, with the kidney remaining negative throughout.
IBV M41 challenge at 5 dpc: IBV was detected in the kidney of Group B1 (IBV-vaccinated) and Group C1 (aMPV + IBV vaccinated), with samples having a high similarity with the vaccine strain (over 99%) (Table 5). All samples from Groups A1 (aMPV vaccinated) and D1 (Non-vaccinated) were positive for IBV, with 100% identity to the challenge strain. Groups not receiving an M41 challenge were IBV negative and no aMPV was detected in birds challenged with M41.
aMPV subtype B challenge at 5 dpc: No aMPV was detected in Group A2 (aMPV vaccinated) or Group C2 (aMPV + IBV vaccinated). The virus was detected in the trachea of birds in Group B2 (IBV vaccinated) and both trachea and kidneys of Group D2 (non-vaccinated).
3.2.6. Determination of IBV viral load
Post vaccination in the IBV vaccinated tracheal samples, viral load increased from 7 dpv to 14 dpv (Group B = 2.48 to 2.85 log REU; Group C = 1.53 to 1.99 log REU) (Fig. 4). However, IBV presence was significantly lower (p < 0.05) at 21 dpv for Group B when compared to both 7 and 14 dpv. The IBV vaccination group (Group B) had a significantly higher tracheal viral load when compared to the combined (Group C) vaccine group at both 7 and 21 dpv. While both groups were negative at 7 dpv, viral load in the kidney was significantly higher at 14 and 21 dpv (p < 0.05) in Group B compared to the combined Group C.
Post challenge with IBV M41, groups vaccinated with IBV were showed to be negative by qRT-PCR, whereas Group A (aMPV vaccinated) and Group D (non-vaccinated) showed readings of 2.55 and 2.42 log REU respectively (Fig. 5).
Post vaccination, kidney samples were negative for IBV from all groups at 7 dpv. However, viral load increased from 14 to 21 dpv in all IBV vaccinated groups (Group B – 2.24 to 2.99 log REU; Group C – 1.50 to 2.08 log REU). Post challenge with IBV M41, groups not receiving an IBV vaccine showed virus presence (Group A1 – 2.10 log REU; Group D1 – 2.36 log REU), with vaccinated groups having low detection levels (Group B1 – 0.25 log REU; Group C1 – 0.06 log REU).
4. Discussion
Co-vaccination of day-old broiler chicks with NDV or aMPV alongside two different live IBV (Mass and 793B) vaccines does not impair protection conferred against either IBV or aMPV challenge. In both experiments, following IBV challenge, mild respiratory signs were observed for the first two days and none thereafter, demonstrating clinical protection conferred by IBV Mass H120 + 793B CR88, with or without NDV or aMPV. Ciliostasis analysis was also used to confirm IBV protection [14], where a higher protection percentage suggests better protection against a virulent IBV challenge. Ciliostasis results in the current study highlighted excellent IBV protection against M41 and QX challenges in groups receiving H120 + 793B, or the same IBV vaccines given along with aMPV or NDV vaccines. Previous work has shown that the combination of Mass + 793B conferred excellent ciliary protection against Asian (QX and Q1) and Middle East (IS/1494/06 and IS/885/00) IBV strains [25,48–51]. When IBV H120 and CR88
Table 5 Experiment 2: IBV or aMPV detections from trachea and kidney samples at 5 days post M41 or virulent aMPV challenge. Groups are indicated as follows: A = aMPV vaccinated, B = IBV vaccinated, C = Combined vaccinated, D = Unvaccinated; 1 = IBV Challenged, 2 = aMPV Challenged, 3 = Unchallenged.
Fig. 4. Experiment 2: Quantification of IBV viral load in the trachea and kidney at 21 dpv following vaccination with either (Group B) aMPV B or (Group C) aMPV B + H120 + CR88. Data is presented ± standard error margins (SEM). Significant differences between groups of the same tissue type (p < 0.05) labelled with different letters.
Fig. 5. Experiment 2: Quantification of IBV viral load in the trachea and kidney at 5 dpc following challenge by virulent IBV M41. Data is presented ± standard error margins (SEM). Significant differences between groups of the same tissue type (p < 0.05) labelled with different letters.
vaccine strains are combined with the NDV or aMPV vaccine, protection levels remained at or above 94%. This reconfirms that such combined vaccination did not compromise the protection conferred against virulent IBVs.
In addition to the cilia-stopping test, detection of virus and viral load in the trachea and kidney of challenged groups was attempted to demonstrate protection against IBV M41 and QX. No vaccine viruses were detected in the trachea at 5 dpc. However, both virulent challenge viruses were present, suggesting host clearance of IBV vaccines by at least 25 dpv [52,53]. The co-administration of either NDV or aMPV alongside the IBV vaccine caused a lower IBV vaccine virus load in both the trachea and kidney at 21 dpv. However, post-challenge with virulent IBV, the combined IBV with NDV or aMPV vaccination had no effect on viral load when compared with IBV vaccination alone. For both experiments, despite relatively higher tracheal viral load, a low REU value was shown in kidney tissue from non-IBV vaccinated groups following M41 and QX challenge. These results, along with the cilia-stopping test, demonstrated that comparable levels of protection are found when IBV H120 and CR88 vaccine strains are given either alone or alongside an NDV or aMPV vaccine.
Control of Newcastle disease is of paramount importance in endemic countries. Control is normally achieved by the use of live and inactivated vaccines [54]. Haemagglutination inhibition titres were traditionally used to confirm protection against NDV challenge [55,56], and the mean NDV HI titre in both single and combined vaccinated groups described here were > 5.63 log2, which is above the protective titre against virulent NDVs [19,23,57,58]. Current work showed that simultaneous vaccination of NDV + H120 + CR88 in commercial broiler chicks does not interfere with the induction of protective immunity against NDV. In this study, following challenge with virulent aMPV, no clinical signs were found in the single (aMPV) or combined (aMPV + H120 + CR88) vaccinated groups, confirming clinical protection against the challenge virus [19,23,59]. Furthermore, following challenge with the virulent strain, no antigen was detected by RT-PCR from tissue samples in either of the vaccinated groups. The aMPV ELISA antibody titres in the triple-vaccinated group was lower than the single vaccinated group, echoing findings from previous work [23]. However, it has been reported that levels of humoral antibodies have no association with clinical protection against aMPV [60,61]. It is evident that experimental vaccination of commercial broilers (with aMPV and IBV MDAs) with aMPV subtype B and two IBV strains (H120 and CR88) had no adverse effects on clinical protection conferred against a virulent challenge.
In both experiments, the persistence of vaccinal strains following individual IBV, NDV, aMPV and combined (H120 + CR88 + NDV; H120 + CR88 + aMPV) vaccination were monitored at intervals. For NDV and aMPV, classical RT-PCRs [32,40] were applied to RNA extracted from oropharyngeal swabs, with no detection of the NDV vaccine at any sampling points. Previous work has shown detection of NDV vaccine virus in SPF chickens for up to four days post vaccination [19,62]. In this experiment, first sampling was carried out at 7 dpv, and the failure to detect NDV may have been due to rapid clearance of the vaccine [19]. For aMPV, it was possible to detect the virus up to 7 dpv using dry oropharyngeal swabs and until 21 dpv from tracheal tissue. While Awad et al. (2015) identified the aMPV vaccine at 21 dpv in a combined IBV + aMPV vaccinated group [19], there was no difference between single and combined vaccinated groups in the current study, suggesting that combined vaccination does not impair a host’s ability to rapidly clear the aMPV vaccine strain. IBV vaccinal strains have previously been shown to persist in chickens for a longer period [53,63]. To further investigate this, the growth kinetics and genetic characteristics of live IBV vaccines used in this study were cross-compared with previous reports [26,64]. Vaccinal strains were detected in OP swabs throughout the current experiment, with the variant 793B strain dominating at the later ages of the birds. With both experiments, Sanger sequencing showed that the genotype of strains recovered using OP swabs altered from Mass to 793B after 14 dpv. As reported previously [26,27], while the Mass genotype made up the majority of detections during early sampling days (3–14 dpv), the 793B strain became the dominant strain by 21 dpv. The reason for the shifting of genotypes from Mass to 793B is not known, however, previous work has highlighted that different IBV strains can differentially induce host expression pathways, in particular TLR7 [65] and IFN-b [66]. Such differences may have played a role in a host’s ability to clear the Mass vaccine virus at an earlier time point when compared to the 793B vaccine. Further work is required to understand potential underlying mechanisms.
At no point did the nucleotide similarity for recovered Mass vaccine strains drop below 98% when compared to the original vaccine sequence. This suggests that under experimental conditions, the vaccine strain underwent minimal alteration following inoculation [26]. Similarly, the 793B vaccine strain remained high for all but one sampling point (99–100% with the exception of 96% at 21 dpv in Experiment 1). Limited genetic changes were noted in the tissue samples, with 3–4 and 4–7.5 SNPs per sequence in the Mass and 793B vaccine strains respectively. A higher number of polymorphisms were seen in the 793B sequences, compared to Massachusetts, indicating that the persistence of this particular genotype may contribute to greater genetic variations within local virus populations [64,67,68]. However, the dS/dN ratios suggest purifying selection for both genotypes [69], and therefore evolutionary constraint [70]. Previous work implied 793B amino acid variations do not follow a consistent pattern over time [71], which was also witnessed in the current study. No persistent alteration was witnessed from any tissues in any of the vaccination programs.
In conclusion, this study indicates that the combined vaccination consisting of NDV (VG/GA-Avinew) or aMPV (subtype B) alongside two strains IBV (Massachusetts - H120 and 793B CR88) does not impair protective immunity against the globally important IBV, NDV or aMPV strains that were included in this study.
This article was originally published in Vaccine 37 (2019) 7566–7575. https://doi.org/10.1016/j.vaccine.2019.09.081. This is an Open Access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/). 






























