Variations in Pathogenicity and Molecular Characterization of Infectious Bursal Disease Virus (IBDV) in Egypt
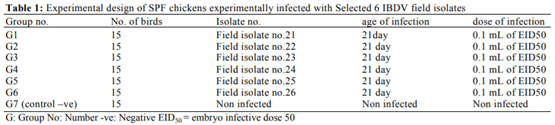
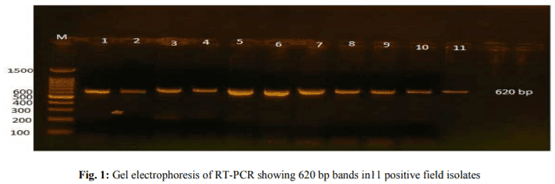
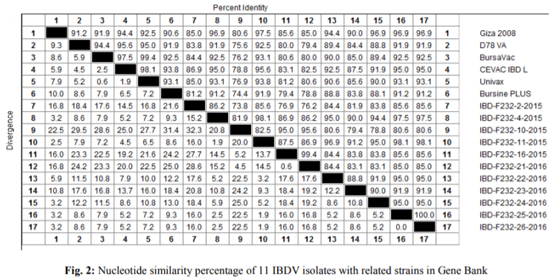
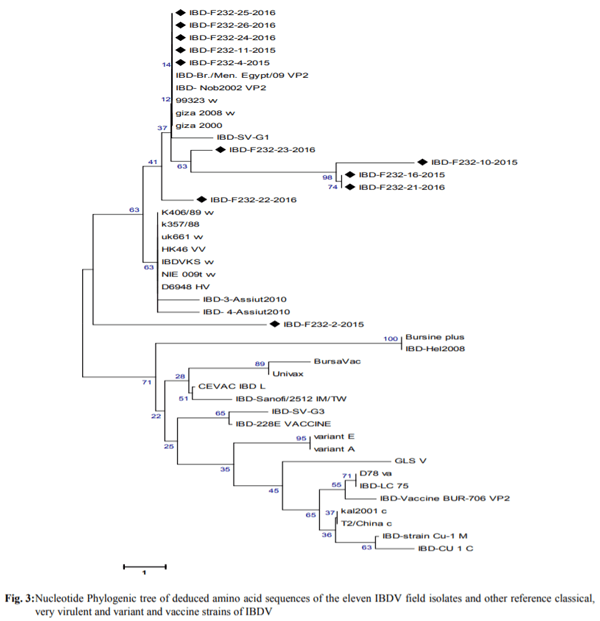
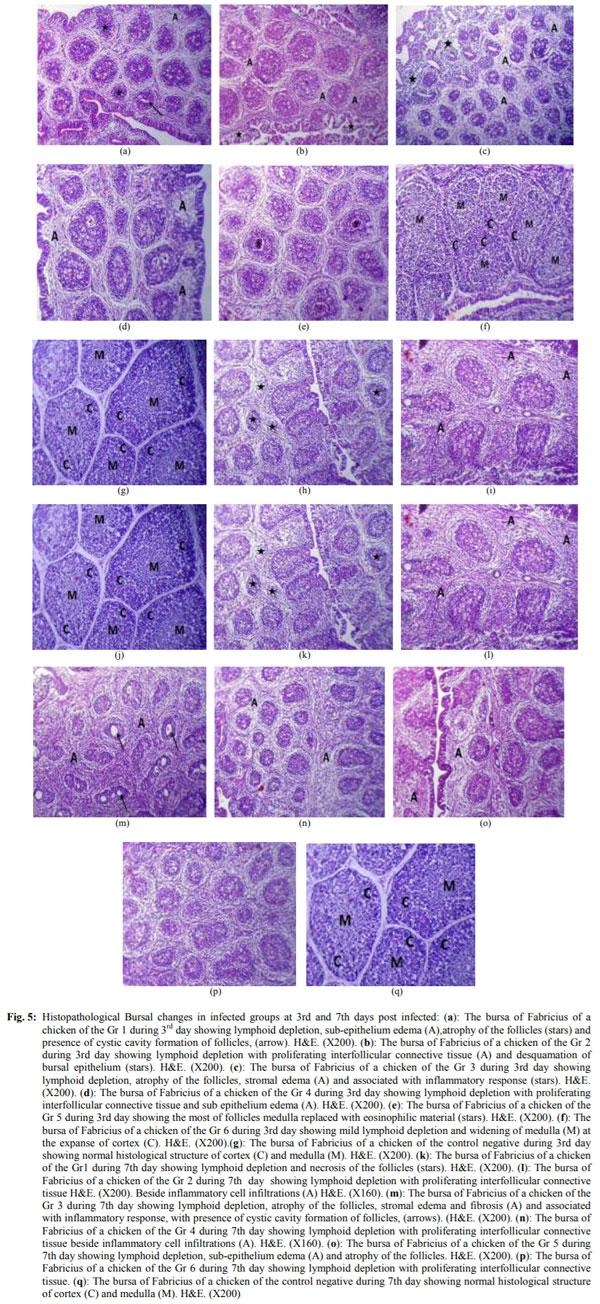
Infectious bursal disease consider a vital threat to poultry industry worldwide and It has special potentiality in Egypt as the country become endemic with several clades from avian influenza from both types highly pathogenic (H5N1 and H5N8) and low pathogenic one (H9N2). The Egyptian government depends on the inactivated vaccine strategy to control avian influenza and any infection with IBD in broiler flocks alter the inactivated vaccine response in addition to other economic loss from the IBD infections. The present study was conducted to investigate the variations in pathogenicity and molecular characterization of IBDV in Egypt. Twenty-six bursal samples were collected from different chicken farms during the period of June 2015 to April 2016; the virus was isolated by inoculation of bursal suspension into 10-11 days ECE via CAM. Reverse Transcriptase Polymerase Chain Reaction (RT-PCR) was applied for IBD targeting Hypervariable Region (HVR) of VP2 gene. Out of 26 tested isolates, 11 isolates were positive by RT-PCR. Sequence analysis of PCR products of 11 positive isolates were carried out and revealed that all +ve isolates were characterized as very virulent (vvIBDV) strains and they showed 80.6-97.5% identity with Giza 2008 IBD strain. Six selected positive vvIBDV isolates were used to study the pathogenicity of IBDV in 105 one-day-old SPF chickens. At day 211st the SPF chickens were divided into 7 groups 15 bird/each, the groups 1-6 were experimentally infected with 0.1 mL of EID50 of 6 selected positive IBDV field isolates via naso- ocular route. Group 7 kept as non-infected control. Experimental infection caused clinical signs and gross lesions typical for IBDV and mortalities ranged from 26.7 to 80% as well as bursal/body weight index lower than 0.7 indicating bursal atrophy. The Microscopical examination of bursal samples collected at days 3 and 7 post-infection revealed different degrees of histopathological bursal changes as lymphoid depletion with proliferating interfollicular connective tissue, atrophy of the follicles, stromal edema and associated with inflammatory response. The circulating strains were highly related to vvIBDV strains belonging to serotype-1 isolates and indicating that they are highly pathogenic to 21-day-old SPF chickens and that vvIBDV strains succeeded in surviving in the Egyptian farms despite of the application of intensive vaccination programs.
Keywords: IBDV, VP2 Gene, Bursa, Variant Strain, vvIBDV


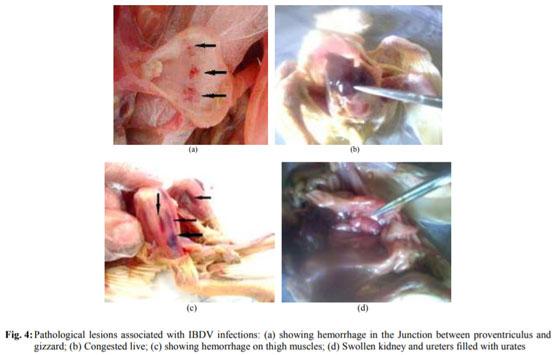
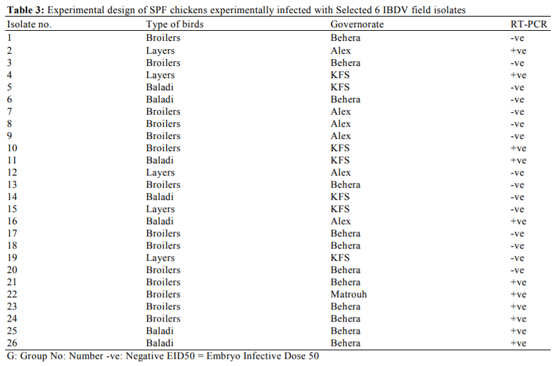
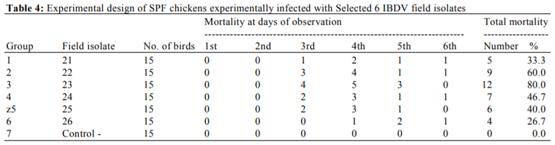
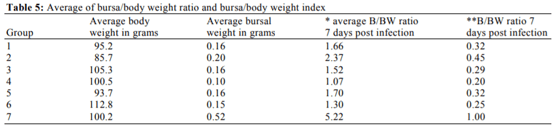
Abouelmaatti, R.R., A.M. Algammal, X. Li, J. Ma and E.A. Abdelnaby et al., 2013. Experimental immunology Cloning and analysis of Nile tilapia Toll-like receptors type-3 mRNA. Central Eur. J. Immunol., 38: 277-282. DOI: 10.5114/ceji.2013.37740
Brandt, M., K. Yao, M. Liu, R.A. Heckert and V.N. Vakharia, 2001. Molecular determinants of virulence, cell tropism and pathogenic phenotype of infectious bursal disease virus. J. Virol., 75: 11974-11982. DOI: 10.1128/JVI.75.24.11974-11982.2001
Cosgrove, A., 1962. An apparently new disease of chickens: Avian nephrosis. Avian Dis., 6: 385-389. DOI: 10.2307/1587909
Culling, C.F.A., 1974. Handbook of Histopathological and Histochemical Techniques: (Including Museum Techniques). 1st Edn., Butterworths, London, ISBN-10: 0407729011, pp: 712.
Eid, H.I., A.M. Algammal, S.A. Nasef, W.K. Elfeil and G.H. Mansour, 2016. Genetic variation among avian pathogenic E. coli strains isolated from broiler chickens. Asian J. Anim. Vet. Adv., 11: 350-356. DOI: 10.3923/ajava.2016.350.356
El-Batrawi, A., 1990. Studies on sever outbreaks of infectious bursal disease 3-deternination of the critiealage of sysceptigilty in maternally immune chicks. Proceedings of the 2nd Science Conference Egypt Vet Poult, (EVP’ 90), pp: 264-269.
El-Sergany, M., A. Moursi, M. Saber and M. Mohamed, 1974. A preliminary investigation on the occurrence of Gumboro disease in Egypt [chickens]. Egypt. J. Vet. Sci. (Egypt).
Elfeil, W.K., R.R. Abouelmaatti, C. Sun, W. Han and X. Li et al., 2012. Identification, cloning, expression of a novel functional Anas platyrhynchos mRNA TLR4. J. Anim. Vet. Adv., 11: 1727-1733. DOI: 10.3923/javaa.2012.1727.1733
Elfeil, W.M.K., A.M. Algammal, R.R. Abouelmaatti, A.R. Gardouh and M.M. Abdel-Daim, 2016. Molecular characterization and analysis of TLR-1 in rabbit tissues. Central Eur. J. Immunol., 41: 236-242. DOI: 10.5114/ceji.2016.63121
Eterradossi, N. and Y.M. Saif, 2013. Infectious Bursal Disease. In: Diseases of Poultry, Suarez, D.L., J.R. Glisson, L.R. McDougald, L.K. Nolan and V. Nair (Eds.), Wiley- Blackwell, pp: 219-246.
Heine, H.G., M. Haritou, P. Failla, K. Fahey and A. Azad, 1991. Sequence analysis and expression of the host-protective immunogen VP2 of a variant strain of infectious bursal disease virus which can circumvent vaccination with standard type I strains. J. General Virol., 72: 1835-1843. DOI: 10.1099/0022-1317-72-8-1835
Hirai, K. and S. Shimakura, 1974. Structure of infectious bursal disease virus. J. Virol., 14: 957-964.
Hussein, E.G.S., 2006. Further studies on infectious bursal disease virus in growing chickens. Alexandria University, Alexandria University.
Islam, M., S. Rahman, M. Noor, E. Chowdhury and H. Müller, 2012. Differentiation of Infectious Bursal Disease Virus (IBDV) genome segment B of very virulent and classical lineage by RT-PCR amplification and restriction enzyme analysis. Arch Virol., 157: 333-336. DOI: 10.1007/s00705-011-1159-9
Jackwood, D., B. Sreedevi, L. LeFever and S. Sommer- Wagner, 2008. Studies on naturally occurring infectious bursal disease viruses suggest that a single amino acid substitution at position 253 in VP2 increases pathogenicity. Virology, 377: 110-116. DOI: 10.1016/j.virol.2008.04.018
Jackwood, D.J., S.E. Sommer-Wagner, B.M. Crossley, S.T. Stoute and P.R. Woolcock et al., 2011. Identification and pathogenicity of a natural reassortant between a very virulent serotype 1 Infectious Bursal Disease Virus (IBDV) and a serotype 2 IBDV. Virology, 420: 98-105. DOI: 10.1016/j.virol.2011.08.023
Leong, J., D. Brown, P. Dobos, F. Kibenge and J. Ludert et al., 2000. Family birnaviridae. Virus taxonomy: Classification and nomenclature of viruses. Seventh Report of the International Committee on Taxonomy of Viruses.
Lucio, B. and S.B. Hitchner, 1979. Infectious bursal disease emulsified vaccine: Effect upon neutralizing-antibody levels in the dam and subsequent protection of the progeny. Avian Dis., 466-478. DOI: 10.2307/1589577
Mawgod, S.A., A.S. Arafa and H.A. Hussein, 2014. Molecular genotyping of the Infectious Bursal Disease Virus (IBDV) isolated from Broiler Flocks in Egypt. Int. J. Vet. Sci. Med., 2: 46-52. DOI: 10.1016/j.ijvsm.2014.02.004
McFerran, J., M. McNulty, E. McKillop, T. Connor and R. McCracken et al., 1980. Isolation and serological studies with infectious bursal disease viruses from fowl, turkeys and ducks: Demonstration of a second serotype. Avian Pathol., 9: 395-404. DOI: 10.1080/03079458008418423
Metwally, A.M., A.A. Yousif, I.B. Shaheed, W.A. Mohammed and A.M. Samy et al., 2009. Re- emergence of very virulent IBDV in Egypt. Int. J. Virol., 5: 1-17. DOI: 10.3923/ijv.2009.1.17
Müller, H., M.R. Islam and R. Raue, 2003. Research on infectious bursal disease-the past, the present and the future. Vet. Microbiol., 97: 153-165. DOI: 10.1016/j.vetmic.2003.08.005
Parkhurst, R.T., 1964. Pattern of Mortality in Avian Nephrosis. Poult. Sci., 43: 788-790. DOI: 10.3382/ps.0430788
Reed, L.J. and H. Muench, 1938. A simple method of estimating fifty percent endpoints. Am J Hygiene.
Rosenberger, J.K., Y.M. Saif and D.J. Jackwood, 1998. Infectious Bursal Disease. In: A Laboratory Manual for the Isolation and Identification of Avian Pathogens, Swayne, D.E., J.R. Glisson, M.W. Jackwood and W.M. Reed (Eds.), American Association of Avian Pathologists, USA, pp: 215-218.
Sultan, H., H.A. Hussein, A.G.A. El-Razik, S. El-Balall and S.M. Talaat and A.A. Shehata, 2012. Efficacy of HVT-IBDV vector vaccine against recent Egyptian vvIBDV in commercial broiler chickens. Int. J. Poultry Sci., 11: 710-710. DOI: 10.3923/ijps.2012.710.717
Van den Berg, T., D. Morales, N. Eterradossi, G. Rivallan and D. Toquin et al., 2004. Assessment of genetic, antigenic and pathotypic criteria for the characterization of IBDV strains. Avian Pathol., 33: 470-476. DOI: 10.1080/03079450400003650
Wlliams, S.M., 2016. A laboratory manual for the isolation, identification and characterization of avian pathogens. American Association of Avian Pathologists, Jacksonville, Fl.









.jpg&w=3840&q=75)