Probiotics, Prebiotics, and Phytogenic Substances for Optimizing Gut Health in Poultry
Author details:
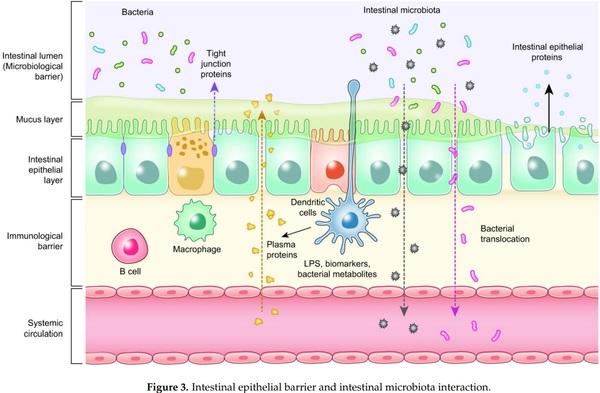
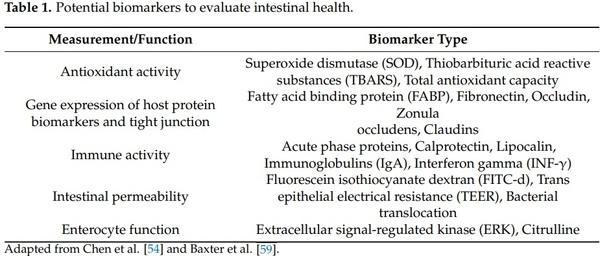

1. BisBischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.-D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal
Permeability—A New Target for Disease Prevention and Therapy. BMC Gastroenterol. 2014, 14, 189. [CrossRef]
2. Zoetendal, E.G.; Rajilic-Stojanovic, M.; de Vos, W.M. High-Throughput Diversity and Functionality Analysis of the Gastrointestinal
Tract Microbiota. Gut 2008, 57, 1605–1615. [CrossRef]
3. Ackermann, W.; Coenen, M.; Schrödl, W.; Shehata, A.A.; Krüger, M. The Influence of Glyphosate on the Microbiota and Production of Botulinum Neurotoxin During Ruminal Fermentation. Curr. Microbiol. 2015, 70, 374–382. [CrossRef]
4. Schrödl, W.; Krüger, S.; Konstantinova-Müller, T.; Shehata, A.A.; Rulff, R.; Krüger, M. Possible Effects of Glyphosate on Mucorales
Abundance in the Rumen of Dairy Cows in Germany. Curr. Microbiol. 2014, 69, 817–823. [CrossRef] [PubMed]
5. Krüger, M.; Neuhaus, J.; Herrenthey, A.G.; Gökce, M.M.; Schrödl, W.; Shehata, A.A. Chronic Botulism in a Saxony Dairy Farm:
Sources, Predisposing Factors, Development of the Disease and Treatment Possibilities. Anaerobe 2014, 28, 220–225. [CrossRef]
6. Abuajamieh, M.; Kvidera, S.K.; Fernandez, M.V.S.; Nayeri, A.; Upah, N.C.; Nolan, E.A.; Lei, S.M.; DeFrain, J.M.; Green, H.B.;
Schoenberg, K.M.; et al. Inflammatory Biomarkers Are Associated with Ketosis in Periparturient Holstein Cows. Res. Vet. Sci.
2016, 109, 81–85. [CrossRef]
7. Hafez, H.M.; Shehata, A.A. Turkey Production and Health: Current Challenges. Ger. J. Vet. Res. 2021, 1, 3–14. [CrossRef]
8. Gernat, A.A.; Santos, F.B.O.; Grimes, J.L. Alternative Approaches to Antimicrobial Use in the Turkey Industry: Challenges and
Perspectives. Ger. J. Vet. Res. 2021, 1, 37–47. [CrossRef]
9. López-García, P.; Eme, L.; Moreira, D. Symbiosis in Eukaryotic Evolution. J. Theor. Biol. 2017, 434, 20–33. [CrossRef]
10. Wren, B.W. Microbial Genome Analysis: Insights into Virulence, Host Adaptation and Evolution. Nat. Rev. Genet. 2000, 1, 30–39.
[CrossRef] [PubMed]
11. McFall-Ngai, M.J. Identifying “Prime Suspects”: Symbioses and the Evolution of Multicellularity. Comp. Biochem. Physiol. B
Biochem. Mol. Biol. 2001, 129, 711–723. [CrossRef]
12. Xu, J.; Mahowald, M.A.; Ley, R.E.; Lozupone, C.A.; Hamady, M.; Martens, E.C.; Henrissat, B.; Coutinho, P.M.; Minx, P.; Latreille,
P.; et al. Evolution of Symbiotic Bacteria in the Distal Human Intestine. PLoS Biol. 2007, 5, e156. [CrossRef] [PubMed]
13. Lee, S.; La, T.-M.; Lee, H.-J.; Choi, I.-S.; Song, C.-S.; Park, S.-Y.; Lee, J.-B.; Lee, S.-W. Characterization of Microbial Communities in the Chicken Oviduct and the Origin of Chicken Embryo Gut Microbiota. Sci. Rep. 2019, 9, 6838. [CrossRef] [PubMed]
14. Kikuchi, Y.; Hosokawa, T.; Nikoh, N.; Meng, X.-Y.; Kamagata, Y.; Fukatsu, T. Host-Symbiont Co-Speciation and Reductive
Genome Evolution in Gut Symbiotic Bacteria of Acanthosomatid Stinkbugs. BMC Biol. 2009, 7, 2. [CrossRef]
15. Tellez, G. Prokaryotes Versus Eukaryotes: Who Is Hosting Whom? Front. Vet. Sci. 2014, 1, 3. [CrossRef] [PubMed]
16. Blaser, M.J. Who Are We? Indigenous Microbes and the Ecology of Human Diseases. EMBO Rep. 2006, 7, 956–960. [CrossRef]
[PubMed]
17. Kau, A.L.; Ahern, P.P.; Griffin, N.W.; Goodman, A.L.; Gordon, J.I. Human Nutrition, the Gut Microbiome and the Immune System.
Nature 2011, 474, 327–336. [CrossRef]
18. Hadrich, D. Microbiome Research Is Becoming the Key to Better Understanding Health and Nutrition. Front. Genet. 2018, 9, 212.
[CrossRef] [PubMed]
19. O’Hara, A.M.; Shanahan, F. The Gut Flora as a Forgotten Organ. EMBO Rep. 2006, 7, 688–693. [CrossRef] [PubMed]
20. Van Der Wielen, P.W.; Biesterveld, S.; Notermans, S.; Hofstra, H.; Urlings, B.A.; van Knapen, F. Role of Volatile Fatty Acids in Development of the Cecal Microflora in Broiler Chickens during Growth. Appl. Environ. MicroBiol. 2000, 66, 2536–2540.
[CrossRef]
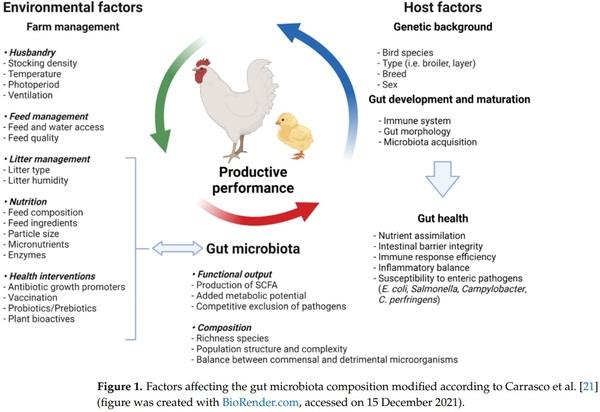
21. Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms 2019, 7, 374. [CrossRef]
22. Adak, A.; Khan, M.R. An Insight into Gut Microbiota and Its Functionalities. Cell Mol. Life Sci. 2019, 76, 473–493. [CrossRef]
23. Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.-P. The Firmicutes/Bacteroidetes
Ratio of the Human Microbiota Changes with Age. BMC MicroBiol. 2009, 9, 123. [CrossRef] [PubMed]
24. Costea, P.I.; Hildebrand, F.; Arumugam, M.; Bäckhed, F.; Blaser, M.J.; Bushman, F.D.; de Vos, W.M.; Ehrlich, S.D.; Fraser, C.M.;
Hattori, M.; et al. Enterotypes in the Landscape of Gut Microbial Community Composition. Nat. MicroBiol. 2018, 3, 8–16.
[CrossRef] [PubMed]
25. Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with
Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [CrossRef] [PubMed]
26. Ley, R.E.; Knight, R.; Gordon, J.I. The Human Microbiome: Eliminating the Biomedical/Environmental Dichotomy in Microbial
Ecology. Environ. MicroBiol. 2007, 9, 3–4. [CrossRef] [PubMed]
27. Zhao, W.; Wang, Y.; Liu, S.; Huang, J.; Zhai, Z.; He, C.; Ding, J.; Wang, J.; Wang, H.; Fan, W.; et al. The Dynamic Distribution of
Porcine Microbiota across Different Ages and Gastrointestinal Tract Segments. PLoS ONE 2015, 10, e0117441. [CrossRef]
28. Subbiah, M.T.R. Nutrigenetics and Nutraceuticals: The next Wave Riding on Personalized Medicine. Transl. Res. 2007, 149, 55–61.
[CrossRef] [PubMed]
29. Shehata, A.M.; Paswan, V.K.; Attia, Y.A.; Abdel-Moneim, A.-M.E.; Abougabal, M.S.; Sharaf, M.; Elmazoudy, R.; Alghafari,
W.T.; Osman, M.A.; Farag, M.R.; et al. Managing Gut Microbiota through in Ovo Nutrition Influences Early-Life Programming in Broiler Chickens. Animals 2021, 11, 3491. [CrossRef]
30. Sugiharto, S. Role of Nutraceuticals in Gut Health and Growth Performance of Poultry. J. Saudi Soc. Agric. Sci. 2016, 15, 99–111.
[CrossRef]
31. Hailu, G.; Boecker, A.; Henson, S.; Cranfield, J. Consumer Valuation of Functional Foods and Nutraceuticals in Canada. A Conjoint
Study Using Probiotics. Appetite 2009, 52, 257–265. [CrossRef] [PubMed]
32. Wareth, G.; Sprague, L.D.; Neubauer, H.; Pletz, M.W. Klebsiella Pneumoniae in Germany: An Overview on Spatiotemporal
Distribution and Resistance Development in Humans. Ger. J. Microbiol. 2021, 1, 16–25. [CrossRef]
33. Kaonga, N.; Hang’ombe, B.M.; Lupindu, A.M.; Hoza, A.S. Detection of CTX-M-Type Extended-Spectrum Beta-Lactamase
Producing Salmonella typhimurium in Commercial Poultry Farms in Copperbelt Province, Zambia. Ger. J. Vet. Res. 2021, 1, 27–34.
[CrossRef]
34. Njeru, J. Emerging Carbapenem Resistance in ESKAPE Organisms in Sub-Saharan Africa and the Way Forward. Ger. J. Microbiol.
2021, 1, 3–6. [CrossRef]
35. Ballou, M.A.; Davis, E.M.; Kasl, B.A. Nutraceuticals. Vet. Clin. North Am. Food Anim. Pract. 2019, 35, 507–534. [CrossRef] [PubMed]
36. Metchnikoff, E. The Prolongation of Life. Optimistic Studies; Chalmers, P.M., Ed.; Heinemann: London, UK, 1907.
37. Dhama, K.; Tiwari, R.; Khan, R.U.; Chakrabort, S.; Gopi, M.; Karthik, K.; Saminathan, M.; Desingu, P.A.; Sunkara, L.T. Growth
Promoters and Novel Feed Additives Improving Poultry Production and Health, Bioactive Principles and Beneficial Applications:
The Trends and Advances-A Review. Int. J. Pharmacol. 2014, 10, 129–159. [CrossRef]
38. Tlaskalová-Hogenová, H.; Stˇepánková, R.; Kozáková, H.; Hudcovic, T.; Vannucci, L.; Tuˇcková, L.; Rossmann, P.; Hrnˇcíˇr, T.;
Kverka, M.; Zákostelská, Z.; et al. The Role of Gut Microbiota (Commensal Bacteria) and the Mucosal Barrier in the Pathogenesis of Inflammatory and Autoimmune Diseases and Cancer: Contribution of Germ-Free and Gnotobiotic Animal Models of Human
Diseases. Cell. Mol. Immunol. 2011, 8, 110–120. [CrossRef] [PubMed]
39. Pourabedin, M.; Zhao, X. Prebiotics and Gut Microbiota in Chickens. FEMS MicroBiol. Lett. 2015, 362, fnv122. [CrossRef]
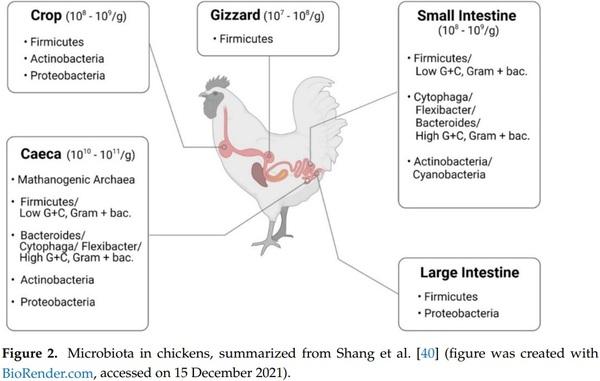
40. Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken Gut Microbiota: Importance and Detection Technology. Front. Vet. Sci. 2018,
5, 254. [CrossRef]
41. Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of Tight Junction Permeability by Intestinal Bacteria and Dietary Components. J. Nutr. 2011, 141, 769–776. [CrossRef] [PubMed]
42. Ballard, S.T.; Hunter, J.H.; Taylor, A.E. Regulation of Tight-Junction Permeability During Nutrient Absorption Across the Intestinal
Epithelium. Annu. Rev. Nutr. 1995, 15, 35–55. [CrossRef]
43. González-Mariscal, L.; Tapia, R.; Chamorro, D. Crosstalk of Tight Junction Components with Signaling Pathways. Biochim.
Biophys. Acta (BBA) Biomembr. 2008, 1778, 729–756. [CrossRef] [PubMed]
44. Harhaj, N.S.; Antonetti, D.A. Regulation of Tight Junctions and Loss of Barrier Function in Pathophysiology. Int. J. Biochem. Cell.
Biol. 2004, 36, 1206–1237. [CrossRef] [PubMed]
45. Faralli, A.; Shekarforoush, E.; Ajalloueian, F.; Mendes, A.C.; Chronakis, I.S. In Vitro Permeability Enhancement of Curcumin across Caco-2 Cells Monolayers Using Electrospun Xanthan-Chitosan Nanofibers. Carbohydr. Polym. 2019, 206, 38–47. [CrossRef]
[PubMed]
46. Wang, J.; Ghosh, S.S.; Ghosh, S. Curcumin Improves Intestinal Barrier Function: Modulation of Intracellular Signaling, and
Organization of Tight Junctions. Am. J. Physiol. Cell. Physiol. 2017, 312, C438–C445. [CrossRef]
47. Baumgard, L.H.; Rhoads, R.P. Effects of Heat Stress on Postabsorptive Metabolism and Energetics. Annu. Rev. Anim. BioSci. 2013,
1, 311–337. [CrossRef]
48. Pearce, S.C.; Mani, V.; Weber, T.E.; Rhoads, R.P.; Patience, J.F.; Baumgard, L.H.; Gabler, N.K. Heat Stress and Reduced Plane of
Nutrition Decreases Intestinal Integrity and Function in Pigs. J. Anim. Sci. 2013, 91, 5183–5193. [CrossRef]
49. Gilani, S.; Chrystal, P.V.; Barekatain, R. Current Experimental Models, Assessment and Dietary Modulations of Intestinal
Permeability in Broiler Chickens. Anim. Nutr. 2021, 7, 801–811. [CrossRef]
50. Kvidera, S.K.; Dickson, M.J.; Abuajamieh, M.; Snider, D.B.; Fernandez, M.V.S.; Johnson, J.S.; Keating, A.F.; Gorden, P.J.; Green,
H.B.; Schoenberg, K.M.; et al. Intentionally Induced Intestinal Barrier Dysfunction Causes Inflammation, Affects Metabolism, and
Reduces Productivity in Lactating Holstein Cows. J. Dairy Sci. 2017, 100, 4113–4127. [CrossRef]
51. Gilani, S.; Howarth, G.S.; Kitessa, S.M.; Tran, C.D.; Forder, R.E.A.; Hughes, R.J. New Biomarkers for Increased Intestinal
Permeability Induced by Dextran Sodium Sulphate and Fasting in Chickens. J. Anim. Physiol. Anim. Nutr. 2017, 101, e237–e245.
[CrossRef]
52. Ducatelle, R.; Goossens, E.; De Meyer, F.; Eeckhaut, V.; Antonissen, G.; Haesebrouck, F.; Van Immerseel, F. Biomarkers for
Monitoring Intestinal Health in Poultry: Present Status and Future Perspectives. Vet. Res. 2018, 49, 43. [CrossRef]
53. Tellez, G.; Latorre, J.D.; Kuttappan, V.A.; Kogut, M.H.; Wolfenden, A.; Hernandez-Velasco, X.; Hargis, B.M.; Bottje, W.G.;
Bielke, L.R.; Faulkner, O.B. Utilization of Rye as Energy Source Affects Bacterial Translocation, Intestinal Viscosity, Microbiota
Composition, and Bone Mineralization in Broiler Chickens. Front. Genet. 2014, 5, 339. [CrossRef]
54. Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of Potential Biomarkers for Gut Barrier Failure in Broiler Chickens.
Front. Vet. Sci. 2015, 2, 14. [CrossRef]
55. Ruff, J.; Tellez, G.; Forga, A.J.; Señas-Cuesta, R.; Vuong, C.N.; Greene, E.S.; Hernandez-Velasco, X.; Uribe, Á.J.; Martínez, B.C.;
Angel-Isaza, J.A.; et al. Evaluation of Three Formulations of Essential Oils in Broiler Chickens under Cyclic Heat Stress. Animals
2021, 11, 1084. [CrossRef]
56. Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier
Model Systems. J. Lab. Autom. 2015, 20, 107–126. [CrossRef] [PubMed]
57. Janssen Duijghuijsen, L.M.; Grefte, S.; de Boer, V.C.J.; Zeper, L.; van Dartel, D.A.M.; van der Stelt, I.; Bekkenkamp-Grovenstein,
M.; van Norren, K.; Wichers, H.J.; Keijer, J. Mitochondrial ATP Depletion Disrupts Caco-2 Monolayer Integrity and Internalizes
Claudin 7. Front. Physiol. 2017, 8, 794. [CrossRef] [PubMed]
58. Xu, J.-X.; Cao, C.-Y.; Sun, Y.-C.; Wang, L.-L.; Li, N.; Xu, S.-W.; Li, J.-L. Effects on Liver Hydrogen Peroxide Metabolism Induced by
Dietary Selenium Deficiency or Excess in Chickens. Biol. Trace Elem. Res. 2014, 159, 174–182. [CrossRef]
59. Baxter, M.F.A.; Latorre, J.D.; Dridi, S.; Merino-Guzman, R.; Hernandez-Velasco, X.; Hargis, B.M.; Tellez-Isaias, G. Identification of
Serum Biomarkers for Intestinal Integrity in a Broiler Chicken Malabsorption Model. Front. Vet. Sci. 2019, 6, 144. [CrossRef]
[PubMed]
60. Windmueller, H.G.; Spaeth, A.E. Source and Fate of Circulating Citrulline. Am. J. Physiol. Endocrinol. Metab. 1981, 241, E473–E480.
[CrossRef] [PubMed]
61. Berkeveld, M.; Langendijk, P.; Verheijden, J.H.M.; Taverne, M.A.M.; van Nes, A.; van Haard, P.; Koets, A.P. Citrulline and
Intestinal Fatty Acid-Binding Protein: Longitudinal Markers of Postweaning Small Intestinal Function in Pigs? J. Anim. Sci. 2008,
86, 3440–3449. [CrossRef]
62. Iizuka, M. Wound Healing of Intestinal Epithelial Cells. WJG 2011, 17, 2161. [CrossRef] [PubMed]
63. Staley, M.; Conners, M.G.; Hall, K.; Miller, L.J. Linking Stress and Immunity: Immunoglobulin A as a Non-Invasive Physiological
Biomarker in Animal Welfare Studies. Horm. Behav. 2018, 102, 55–68. [CrossRef] [PubMed]
64. Utech, M.; Ivanov, A.I.; Samarin, S.N.; Bruewer, M.; Turner, J.R.; Mrsny, R.J.; Parkos, C.A.; Nusrat, A. Mechanism of IFN-γ-Induced
Endocytosis of Tight Junction Proteins: Myosin II-Dependent Vacuolarization of the Apical Plasma Membrane. MBoC 2005, 16,
5040–5052. [CrossRef]
65. Nava, P.; Koch, S.; Laukoetter, M.G.; Lee, W.Y.; Kolegraff, K.; Capaldo, C.T.; Beeman, N.; Addis, C.; Gerner-Smidt, K.; Neumaier,
I.; et al. Interferon-γ Regulates Intestinal Epithelial Homeostasis through Converging β-Catenin Signaling Pathways. Immunity
2010, 32, 392–402. [CrossRef] [PubMed]
66. Awad, W.A.; Ghareeb, K.; Böhm, J. Evaluation of the Chicory Inulin Efficacy on Ameliorating the Intestinal Morphology and
Modulating the Intestinal Electrophysiological Properties in Broiler Chickens: Evaluation of the Chicory Inulin Efficacy in Broiler
Chickens. J. Anim. Physiol. Anim. Nutr. 2011, 95, 65–72. [CrossRef]
67. Wideman, R.F.; Hamal, K.R.; Stark, J.M.; Blankenship, J.; Lester, H.; Mitchell, K.N.; Lorenzoni, G.; Pevzner, I. A Wire-Flooring
Model for Inducing Lameness in Broilers: Evaluation of Probiotics as a Prophylactic Treatment. Poult. Sci. 2012, 91, 870–883.
[CrossRef]
68. Wideman, R.F.; Al-Rubaye, A.; Kwon, Y.M.; Blankenship, J.; Lester, H.; Mitchell, K.N.; Pevzner, I.Y.; Lohrmann, T.; Schleifer, J.
Prophylactic Administration of a Combined Prebiotic and Probiotic, or Therapeutic Administration of Enrofloxacin, to Reduce the Incidence of Bacterial Chondronecrosis with Osteomyelitis in Broilers. Poult. Sci. 2015, 94, 25–36. [CrossRef]
69. Saleh, M.; Elson, C.O. Experimental Inflammatory Bowel Disease: Insights into the Host-Microbiota Dialog. Immunity 2011, 34,
293–302. [CrossRef]
70. Latorre, J.D.; Hernandez-Velasco, X.; Bielke, L.R.; Vicente, J.L.; Wolfenden, R.; Menconi, A.; Hargis, B.M.; Tellez, G. Evaluation of a Bacillus Direct-Fed Microbial Candidate on Digesta Viscosity, Bacterial Translocation, Microbiota Composition and Bone
Mineralisation in Broiler Chickens Fed on a Rye-Based Diet. Br. Poult. Sci. 2015, 56, 723–732. [CrossRef]
71. Baxter, M.F.A.; Merino-Guzman, R.; Latorre, J.D.; Mahaffey, B.D.; Yang, Y.; Teague, K.D.; Graham, L.E.; Wolfenden, A.D.;
Hernandez-Velasco, X.; Bielke, L.R.; et al. Optimizing Fluorescein Isothiocyanate Dextran Measurement as a Biomarker in a 24-h
Feed Restriction Model to Induce Gut Permeability in Broiler Chickens. Front. Vet. Sci. 2017, 4, 56. [CrossRef]
72. Furuhashi, M.; Hotamisligil, G.S. Fatty Acid-Binding Proteins: Role in Metabolic Diseases and Potential as Drug Targets. Nat. Rev.
Drug Discov. 2008, 7, 489–503. [CrossRef] [PubMed]
73. Albala, C.; Santos, J.L.; Cifuentes, M.; Villarroel, A.C.; Lera, L.; Liberman, C.; Angel, B.; Pérez-Bravo, F. Intestinal FABP2 A54T
Polymorphism: Association with Insulin Resistance and Obesity in Women. Obes. Res. 2004, 12, 340–345. [CrossRef]
74. Dal Pont, G.C.; Belote, B.L.; Lee, A.; Bortoluzzi, C.; Eyng, C.; Sevastiyanova, M.; Khadem, A.; Santin, E.; Farnell, Y.Z.; Gougoulias,
C.; et al. Novel Models for Chronic Intestinal Inflammation in Chickens: Intestinal Inflammation Pattern and Biomarkers. Front.
Immunol. 2021, 12, 676628. [CrossRef]
75. Schrezenmeir, J.; de Vrese, M. Probiotics, Prebiotics, and Synbiotics-Approaching a Definition. Am J. Clin. Nutr. 2001, 73,
361S–364S. [CrossRef]
76. Yurong, Y.; Ruiping, S.; Shimin, Z.; Yibao, J. Effect of Probiotics on Intestinal Mucosal Immunity and Ultrastructure of Cecal
Tonsils of Chickens. Arch. Anim. Nutr. 2005, 59, 237–246. [CrossRef] [PubMed]
77. Vanderpool, C.; Yan, F.; Polk, D.B. Mechanisms of Probiotic Action: Implications for Therapeutic Applications in Inflammatory
Bowel Diseases. Inflamm. Bowel Dis. 2008, 14, 1585–1596. [CrossRef]
78. Prado-Rebolledo, O.F.; de Jesus Delgado-Machuca, J.; Macedo-Barragan, R.J.; Garcia-Márquez, L.J.; Morales-Barrera, J.E.; Latorre,
J.D.; Hernandez-Velasco, X.; Tellez, G. Evaluation of a Selected Lactic Acid Bacteria-Based Probiotic on Salmonella enterica Serovar
Enteritidis Colonization and Intestinal Permeability in Broiler Chickens. Avian Pathol. 2017, 46, 90–94. [CrossRef] [PubMed]
79. Molinaro, F.; Paschetta, E.; Cassader, M.; Gambino, R.; Musso, G. Probiotics, Prebiotics, Energy Balance, and Obesity: Mechanistic
Insights and Therapeutic Implications. Gastroenterol. Clin. North Am. 2012, 41, 843–854. [CrossRef]
80. Higgins, J.P.; Higgins, S.E.; Vicente, J.L.; Wolfenden, A.D.; Tellez, G.; Hargis, B.M. Temporal Effects of Lactic Acid Bacteria
Probiotic Culture on Salmonella in Neonatal Broilers. Poult. Sci. 2007, 86, 1662–1666. [CrossRef]
81. Vicente, J.; Wolfenden, A.; Torres-Rodriguez, A.; Higgins, S.; Tellez, G.; Hargis, B. Effect of a Lactobacillus Species-Based Probiotic and Dietary Lactose Prebiotic on Turkey Poult Performance with or Without Salmonella enteritidis Challenge. J. Appl. Poult. Res.
2007, 16, 361–364. [CrossRef]
82. Menconi, A.; Wolfenden, A.D.; Shivaramaiah, S.; Terraes, J.C.; Urbano, T.; Kuttel, J.; Kremer, C.; Hargis, B.M.; Tellez, G. Effect of
Lactic Acid Bacteria Probiotic Culture for the Treatment of Salmonella Enterica Serovar Heidelberg in Neonatal Broiler Chickens and Turkey Poults. Poult. Sci. 2011, 90, 561–565. [CrossRef]
83. Higgins, S.E.; Torres-Rodriguez, A.; Vicente, J.L.; Sartor, C.D.; Pixley, C.M.; Nava, G.M.; Tellez, G.; Barton, J.T.; Hargis, B.M.
Evaluation of Intervention Strategies for Idiopathic Diarrhea in Commercial Turkey Brooding Houses. J. Appl. Poult. Res. 2005, 14,
345–348. [CrossRef]
84. Torres-Rodriguez, A.; Higgins, S.E.; Vicente, J.L.S.; Wolfenden, A.D.; Gaona-Ramirez, G.; Barton, J.T.; Tellez, G.; Donoghue, A.M.;
Hargis, B.M. Effect of Lactose as a Prebiotic on Turkey Body Weight Under Commercial Conditions. J. Appl. Poult. Res. 2007, 16,
635–641. [CrossRef]
85. Higgins, S.E.; Wolfenden, A.D.; Tellez, G.; Hargis, B.M.; Porter, T.E. Transcriptional Profiling of Cecal Gene Expression in Probioticand Salmonella-Challenged Neonatal Chicks. Poult Sci. 2011, 90, 901–913. [CrossRef] [PubMed]
86. Tellez-Isaias, G.; Vuong, C.N.; Graham, B.D.; Selby, C.M.; Graham, L.E.; Señas-Cuesta, R.; Barros, T.L.; Beer, L.C.; Coles, M.E.;
Forga, A.J.; et al. Developing Probiotics, Prebiotics, and Organic Acids to Control Salmonella Spp. in Commercial Turkeys at the University of Arkansas, USA. Ger. J. Vet. Res. 2021, 1, 7–12. [CrossRef]
87. Vreeland, R.H.; Rosenzweig, W.D.; Powers, D.W. Isolation of a 250 Million-Year-Old Halotolerant Bacterium from a Primary Salt
Crystal. Nature 2000, 407, 897–900. [CrossRef]
88. Hong, H.A.; Duc, L.H.; Cutting, S.M. The Use of Bacterial Spore Formers as Probiotics. FEMS MicroBiol. Rev. 2005, 29, 813–835.
[CrossRef]
89. Shivaramaiah, S.; Pumford, N.R.; Morgan, M.J.; Wolfenden, R.E.; Wolfenden, A.D.; Torres-Rodríguez, A.; Hargis, B.M.; Téllez,
G. Evaluation of Bacillus Species as Potential Candidates for Direct-Fed Microbials in Commercial Poultry. Poult. Sci. 2011, 90,
1574–1580. [CrossRef]
90. Wolfenden, R.E.; Pumford, N.R.; Morgan, M.J.; Shivaramaiah, S.; Wolfenden, A.D.; Pixley, C.M.; Green, J.; Tellez, G.; Hargis, B.M.
Evaluation of Selected Direct-Fed Microbial Candidates on Live Performance and Salmonella Reduction in Commercial Turkey
Brooding Houses. Poult. Sci. 2011, 90, 2627–2631. [CrossRef]
91. Wolfenden, R.E.; Pumford, N.R.; Morgan, M.J.; Shivaramai, S.; Wolfenden, A.D.; Tellez, G.; Hargis, B.M. Evaluation of a Screening and Selection Method for Bacillus Isolates for Use as Effective Direct-Fed Microbials in Commercial Poultry. Int. J. Poult. Sci. 2010,
9, 317–323. [CrossRef]
92. Sen, S.; Ingale, S.L.; Kim, Y.W.; Kim, J.S.; Kim, K.H.; Lohakare, J.D.; Kim, E.K.; Kim, H.S.; Ryu, M.H.; Kwon, I.K.; et al. Effect of
Supplementation of Bacillus Subtilis LS 1-2 to Broiler Diets on Growth Performance, Nutrient Retention, Caecal Microbiology and Small Intestinal Morphology. Res. Vet. Sci. 2012, 93, 264–268. [CrossRef] [PubMed]
93. Latorre, J.D.; Hernandez-Velasco, X.; Kallapura, G.; Menconi, A.; Pumford, N.R.; Morgan, M.J.; Layton, S.L.; Bielke, L.R.; Hargis,
B.M.; Téllez, G. Evaluation of Germination, Distribution, and Persistence of Bacillus subtilis Spores through the Gastrointestinal
Tract of Chickens. Poult. Sci. 2014, 93, 1793–1800. [CrossRef]
94. Bedford, M.R.; Schulze, H. Exogenous Enzymes for Pigs and Poultry. Nutr. Res. Rev. 1998, 11, 91–114. [CrossRef]
95. Esteve-Garcia, E.; Brufau, J.; Pérez-Vendrell, A.; Miquel, A.; Duven, K. Bioefficacy of Enzyme Preparations Containing BetaGlucanase and Xylanase Activities in Broiler Diets Based on Barley or Wheat, in Combination with Flavomycin. Poult. Sci. 1997,
76, 1728–1737. [CrossRef] [PubMed]
96. Latorre, J.D.; Hernandez-Velasco, X.; Kogut, M.H.; Vicente, J.L.; Wolfenden, R.; Wolfenden, A.; Hargis, B.M.; Kuttappan, V.A.;
Tellez, G. Role of a Bacillus Subtilis Direct-Fed Microbial on Digesta Viscosity, Bacterial Translocation, and Bone Mineralization in Turkey Poults Fed with a Rye-Based Diet. Front. Vet. Sci. 2014, 1, 26. [CrossRef] [PubMed]
97. Adhikari, B.; Hernandez-Patlan, D.; Solis-Cruz, B.; Kwon, Y.M.; Arreguin, M.A.; Latorre, J.D.; Hernandez-Velasco, X.; Hargis,
B.M.; Tellez-Isaias, G. Evaluation of the Antimicrobial and Anti-Inflammatory Properties of Bacillus-DFM (NorumTM) in Broiler
Chickens Infected with Salmonella Enteritidis. Front. Vet. Sci. 2019, 6, 282. [CrossRef]
98. Solis-Cruz, B.; Hernandez-Patlan, D.; Petrone, V.M.; Pontin, K.P.; Latorre, J.D.; Beyssac, E.; Hernandez-Velasco, X.; Merino-Guzman, R.;
Arreguin, M.A.; Hargis, B.M.; et al. Evaluation of a Bacillus -Based Direct-Fed Microbial on Aflatoxin B1 Toxic Effects, Performance,
Immunologic Status, and Serum Biochemical Parameters in Broiler Chickens. Avian Dis. 2019, 63, 659–669. [CrossRef]
99. Black, S.; Fahrenholz, A.; Grimes, J.L. The effect of a direct-fed microbial and dietary fat inclusion on performance and energy metabolism in broiler chicks and turkey poults. Ger. J. Vet. Res. 2021, 1, 1–10. [CrossRef]
100. Liu, X.; Cao, S.; Zhang, X. Modulation of Gut Microbiota-Brain Axis by Probiotics, Prebiotics, and Diet. J. Agric. Food Chem. 2015,
63, 7885–7895. [CrossRef]
101. Hamilton-Miller, J.M.T. Probiotics and Prebiotics in the Elderly. Postgrad. Med. J. 2004, 80, 447–451. [CrossRef]
102. Hedin, C.; Whelan, K.; Lindsay, J.O. Evidence for the Use of Probiotics and Prebiotics in Inflammatory Bowel Disease: A Review of Clinical Trials. Proc. Nutr. Soc. 2007, 66, 307–315. [CrossRef] [PubMed]
103. Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.G.; Muccioli, G.M.; Neyrinck, A.M.; Possemiers, S.; Van Holle,
A.; François, P.; et al. Responses of Gut Microbiota and Glucose and Lipid Metabolism to Prebiotics in Genetic Obese and
Diet-Induced Leptin-Resistant Mice. Diabetes 2011, 60, 2775–2786. [CrossRef] [PubMed]
104. Ducatelle, R.; Eeckhaut, V.; Haesebrouck, F.; Van Immerseel, F. A Review on Prebiotics and Probiotics for the Control of Dysbiosis:
Present Status and Future Perspectives. Animal 2015, 9, 43–48. [CrossRef] [PubMed]
105. Ajuwon, K.M. Toward a Better Understanding of Mechanisms of Probiotics and Prebiotics Action in Poultry Species. J. Appl.
Poult. Res. 2016, 25, 277–283. [CrossRef]
106. Collins, M.D.; Gibson, G.R. Probiotics, Prebiotics, and Synbiotics: Approaches for Modulating the Microbial Ecology of the Gut.
Am. J. Clin. Nutr. 1999, 69, 1052S–1057S. [CrossRef]
107. Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, Prebiotics and Synbiotics- a Review. J. Food Sci. Technol. 2015, 52, 7577–7587.
[CrossRef]
108. Van den Broek, L.A.M.; Hinz, S.W.A.; Beldman, G.; Vincken, J.-P.; Voragen, A.G.J. Bifidobacterium Carbohydrases-Their Role in Breakdown and Synthesis of (Potential) Prebiotics. Mol. Nutr. Food Res. 2008, 52, 146–163. [CrossRef]
109. Dhama, K.; Mahendran, M.; Tomar, S.; Chauhan, R. Beneficial Effects of Probiotics and Prebiotics in Livestock and Poultry:
The Current Perspectives. Intas. Polivet. 2008, 9, 1–12.
110. Janssens, G.P.J.; Millet, S.; Van Immerseel, F.; De Buck, J.; Hesta, M. The Impact of Prebiotics and Salmonellosis on Apparent
Nutrient Digestibility and Salmonella typhimurium Var. Copenhagen Excretion in Adult Pigeons (Columba Livia Domestica). Poult.
Sci. 2004, 83, 1884–1890. [CrossRef]
111. Parracho, H.; McCartney, A.L.; Gibson, G.R. Probiotics and Prebiotics in Infant Nutrition. Proc. Nutr. Soc. 2007, 66, 405–411.
[CrossRef]
112. Teitelbaum, J.E.; Walker, W.A. Nutritional Impact of Pre- and Probiotics as Protective Gastrointestinal Organisms. Annu. Rev.
Nutr. 2002, 22, 107–138. [CrossRef] [PubMed]
113. Yalcin, S.; Ilyas, O.; Handan, E. Effects of dietary yeast cell wall on performance, egg quality and humoral immune response in laying hens. Ank. Üniv. Vet. Fakültesi Derg. 2014, 61, 289–294. [CrossRef]
114. Harms, R.H.; Miles, R.D. Influence of Fermacto on the Performance of Laying Hens When Fed Diets with Different Levels of
Methionine. Poult. Sci. 1988, 67, 842–844. [CrossRef] [PubMed]
115. Torres-Rodriguez, A.; Sartor, C.; Higgins, S.E.; Wolfenden, A.D.; Bielke, L.R.; Pixley, C.M.; Sutton, L.; Tellez, G.; Hargis, B.M.
Effect of Aspergillus Meal Prebiotic (Fermacto) on Performance of Broiler Chickens in the Starter Phase and Fed Low Protein
Diets. J. Appl. Poult. Res. 2005, 14, 665–669. [CrossRef]
116. Uchima, C.A.; Tokuda, G.; Watanabe, H.; Kitamoto, K.; Arioka, M. Heterologous Expression and Characterization of a GlucoseStimulated β-Glucosidase from the Termite Neotermes Koshunensis in Aspergillus Oryzae. Appl. MicroBiol. Biotechnol. 2011, 89,
1761–1771. [CrossRef]
117. Hernandez-Patlan, D.; Solis-Cruz, B.; Hargis, B.M.; Tellez, G. Chitoneous Materials for Control of Foodborne Pathogens and
Mycotoxins in Poultry. In Chitin-Chitosan—Myriad Functionalities in Science and Technology; Dongre, R.S., Ed.; InTech: London, UK,
2018; ISBN 978-1-78923-406-0.
118. Jonker, D.; Kuper, C.F.; Maquet, V.; Nollevaux, G.; Gautier, S. Subchronic (13-Week) Oral Toxicity Study in Rats with Fungal
Chitin-Glucan from Aspergillus Niger. Food Chem. Toxicol. 2010, 48, 2695–2701. [CrossRef] [PubMed]
119. Hooge, D.M.; Sims, M.D.; Sefton, A.E.; Spring, P.; Connolly, A. Effect of Dietary Mannan Oligosaccharide, With or Without
Bacitracin or Virginiamycin, on Live Performance of Broiler Chickens at Relatively High Stocking Density on New Litter. J. Appl.
Poult. Res. 2003, 12, 461–467. [CrossRef]
120. Kim, W.K.; Donalson, L.M.; Mitchell, A.D.; Kubena, L.F.; Nisbet, D.J.; Ricke, S.C. Effects of Alfalfa and Fructooligosaccharide on
Molting Parameters and Bone Qualities Using Dual Energy X-ray Absorptiometry and Conventional Bone Assays. Poult. Sci.
2006, 85, 15–20. [CrossRef]
121. Tellez, G.; Nava, G.M.; Vicente, J.L.; De Frances, M.; Morales, E.J.; Prado, O.; Terraes, J.C.; Hargis, B.M. Evaluation of Dietary
Aspergillus Meal on Intestinal Morphometry in Turkey Poults. Int. J. Poult. Sci. 2010, 9, 875–878. [CrossRef]
122. Amirdahri, S.; Janmohammadi, H.; Taghizadeh, A.; Rafat, S.A. Effect of Dietary Aspergillus Meal Prebiotic on Growth Performance, Carcass Characteristics, Nutrient Digestibility, and Serum Lipid Profile in Broiler Chick Low-Protein Diets. Turk. J. Vet.
Anim. Sci. 2012, 36. [CrossRef]
123. Reginatto, A.R.; Menconi, A.; Londero, A.; Lovato, M.; Rosa, A.P.; Shivaramai, S.; Wolfenden, A.D.; Huff, W.E.; Huff, G.R.; Rath,
N.C.; et al. Effects of Dietary Aspergillus Meal Prebiotic on Turkey Poults Production Parameters and Bone Qualities. Int. J. Poult.
Sci. 2011, 10, 496–499. [CrossRef]
124. Scholz-Ahrens, K.E.; Ade, P.; Marten, B.; Weber, P.; Timm, W.; Açil, Y.; Glüer, C.-C.; Schrezenmeir, J. Prebiotics, Probiotics, and
Synbiotics Affect Mineral Absorption, Bone Mineral Content, and Bone Structure. J. Nutr. 2007, 137, 838S–846S. [CrossRef]
[PubMed]
125. Van Immerseel, F.; De Buck, J.; De Smet, I.; Mast, J.; Haesebrouck, F.; Ducatelle, R. Dynamics of Immune Cell Infiltration in the Caecal Lamina Propria of Chickens after Neonatal Infection with a Salmonella enteritidis Strain. Dev. Comp. Immunol. 2002,
26, 355–364. [CrossRef]
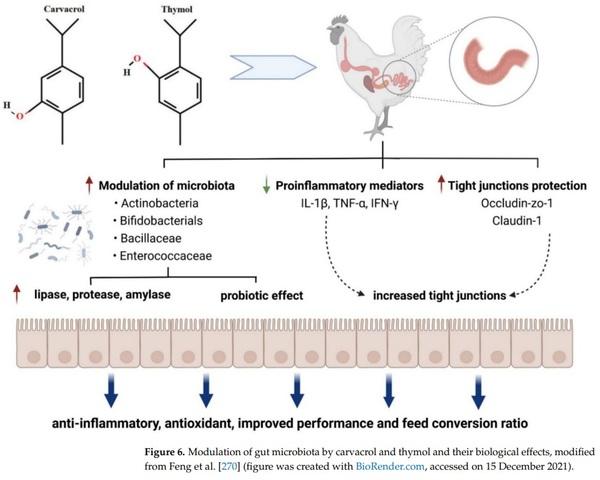
126. Burkholder, K.M.; Thompson, K.L.; Einstein, M.E.; Applegate, T.J.; Patterson, J.A. Influence of Stressors on Normal Intestinal
Microbiota, Intestinal Morphology, and Susceptibility to Salmonella enteritidis Colonization in Broilers. Poult. Sci. 2008, 87,
1734–1741. [CrossRef] [PubMed]
127. Filipkowska, U.; Jó´zwiak, T.; Szymczyk, P. Application of cross-linked chitosan for phosphate removal from aqueous solutions.
Prog. Chem. Appl. Chitin Its Deriv. Volume XIX 2014, 19, 5–14. [CrossRef]
128. Ravi Kumar, M.N.V. A Review of Chitin and Chitosan Applications. React. Funct. Polym. 2000, 46, 1–27. [CrossRef]
129. Londero, A.; Menconi, A.; Reginatto, A.R.; Bacocina, I.; Wolfenden, A.; Shivaramai, S.; Hargis, B.M.; Tellez, G. Effect of an Aspergillus Meal Prebiotic on Salmonella Infection in Turkeys and Broiler Chickens. Int. J. Poult. Sci. 2011, 10, 946–951.
[CrossRef]
130. Yalçin, S.; Yalçin, S.; Eser, H.; ¸Sahin, A.; Yalçin, S.S.; Güçer, ¸S. Effects of Dietary Yeast Cell Wall Supplementation on Performance,
Carcass Characteristics, Antibody Production and Histopathological Changes in Broilers. Kafkas Univ. Vet. Fak. Derg. 2014, 20.
[CrossRef]
131. Awad, W.A.; Ghareeb, K.; Abdel-Raheem, S.; Böhm, J. Effects of Dietary Inclusion of Probiotic and Synbiotic on Growth
Performance, Organ Weights, and Intestinal Histomorphology of Broiler Chickens. Poult. Sci. 2009, 88, 49–56. [CrossRef]
132. Maiorano, G.; Sobolewska, A.; Cianciullo, D.; Walasik, K.; Elminowska-Wenda, G.; Slawinska, A.; Tavaniello, S.; Zylinska, J.;
Bardowski, J.; Bednarczyk, M. Influence of in Ovo Prebiotic and Synbiotic Administration on Meat Quality of Broiler Chickens.
Poult. Sci. 2012, 91, 2963–2969. [CrossRef]
133. Yang, X.J.; Li, W.L.; Feng, Y.; Yao, J.H. Effects of Immune Stress on Growth Performance, Immunity, and Cecal Microflora in Chickens. Poult. Sci. 2011, 90, 2740–2746. [CrossRef] [PubMed]
134. Dimitrov, D.V. The Human Gutome: Nutrigenomics of the Host-Microbiome Interactions. OMICS 2011, 15, 419–430. [CrossRef]
[PubMed]
135. Tellez, G.; Higgins, S.E.; Donoghue, A.M.; Hargis, B.M. Digestive Physiology and the Role of Microorganisms. J. Appl. Poult. Res.
2006, 15, 136–144. [CrossRef]
136. Weiss, A.S.; Burrichter, A.G.; Durai Raj, A.C.; von Strempel, A.; Meng, C.; Kleigrewe, K.; Münch, P.C.; Rössler, L.; Huber, C.;
Eisenreich, W.; et al. In Vitro Interaction Network of a Synthetic Gut Bacterial Community. ISME J. 2021, 1–15. [CrossRef]
137. Plöger, S.; Stumpff, F.; Penner, G.B.; Schulzke, J.-D.; Gäbel, G.; Martens, H.; Shen, Z.; Günzel, D.; Aschenbach, J.R. Microbial
Butyrate and Its Role for Barrier Function in the Gastrointestinal Tract. Ann. N. Y. Acad. Sci. 2012, 1258, 52–59. [CrossRef]
[PubMed]
138. Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Guan, N.L.; et al.
The Gut Microbiota Influences Blood-Brain Barrier Permeability in Mice. Sci. Transl. Med. 2014, 6, 263ra158. [CrossRef]
139. Shen, J.; Obin, M.S.; Zhao, L. The Gut Microbiota, Obesity and Insulin Resistance. Mol. Aspects Med. 2013, 34, 39–58. [CrossRef]
[PubMed]
140. Begley, M.; Gahan, C.G.M.; Hill, C. The Interaction between Bacteria and Bile. FEMS MicroBiol. Rev. 2005, 29, 625–651. [CrossRef]
141. Hofmann, A.F. Bile Acids: The Good, the Bad, and the Ugly. News Physiol. Sci. 1999, 14, 24–29. [CrossRef]
142. Calik, A.; Ceylan, A.; Ekim, B.; Adabi, S.G.; Dilber, F.; Bayraktaroglu, A.G.; Tekinay, T.; Özen, D.; Sacakli, P. The Effect of
Intra-Amniotic and Posthatch Dietary Synbiotic Administration on the Performance, Intestinal Histomorphology, Cecal Microbial
Population, and Short-Chain Fatty Acid Composition of Broiler Chickens. Poult. Sci. 2017, 96, 169–183. [CrossRef]
143. Montagne, L.; Piel, C.; Lallès, J.P. Effect of Diet on Mucin Kinetics and Composition: Nutrition and Health Implications. Nutr. Rev.
2004, 62, 105–114. [CrossRef]
144. Schippa, S.; Conte, M. Dysbiotic Events in Gut Microbiota: Impact on Human Health. Nutrients 2014, 6, 5786–5805. [CrossRef]
[PubMed]
145. EC (European Commission). Regulation 1831 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition. Off. J. Eur. Union L. 2003, 268, 29–43.
146. Giannenas, I.; Florou-Paneri, P.; Papazahariadou, M.; Christaki, E.; Botsoglou, N.A.; Spais, A.B. Effect of Dietary Supplementation with Oregano Essential Oil on Performance of Broilers after Experimental Infection with Eimeria tenella. Arch. Anim. Nutr. 2003,
57, 99–106. [CrossRef] [PubMed]
147. Isabel, B.; Santos, Y. Effects of Dietary Organic Acids and Essential Oils on Growth Performance and Carcass Characteristics of
Broiler Chickens. J. Appl. Poult. Res. 2009, 18, 472–476. [CrossRef]
148. Jamroz, D.; Wiliczkiewicz, A.; Wertelecki, T.; Orda, J.; Skorupi ´nska, J. Use of Active Substances of Plant Origin in Chicken Diets
Based on Maize and Locally Grown Cereals. Br. Poult. Sci. 2005, 46, 485–493. [CrossRef]
149. McReynolds, J.; Waneck, C.; Byrd, J.; Genovese, K.; Duke, S.; Nisbet, D. Efficacy of Multistrain Direct-Fed Microbial and
Phytogenetic Products in Reducing Necrotic Enteritis in Commercial Broilers. Poult. Sci. 2009, 88, 2075–2080. [CrossRef]
150. Murugesan, G.R.; Syed, B.; Haldar, S.; Pender, C. Phytogenic Feed Additives as an Alternative to Antibiotic Growth Promoters in Broiler Chickens. Front. Vet. Sci. 2015, 2, 21. [CrossRef]
151. Yener, Y.; Yalçin, S.; Çolpan, ˙I. Effects of Dietary Supplementation of Red Ginseng Root Powder on Performance, Immune System,
Cecal Microbial Population and Some Blood Parameters in Broilers. Ank. Üniv. Vet. Fakültesi Derg. 2020, 68, 137–145. [CrossRef]
152. Mathe, A. Essential Oils: Basic and Applied Research. Allured Publishing Corporation, Carol Stream. In Essential Oils as Phytogenic
Feed Additives; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; pp. 315–325.
153. Salaheen, S.; Kim, S.-W.; Haley, B.J.; Van Kessel, J.A.S.; Biswas, D. Alternative Growth Promoters Modulate Broiler Gut Microbiome and Enhance Body Weight Gain. Front. Microbiol. 2017, 8, 2088. [CrossRef]
154. Li, Y.; Fu, X.; Ma, X.; Geng, S.; Jiang, X.; Huang, Q.; Hu, C.; Han, X. Intestinal Microbiome-Metabolome Responses to Essential
Oils in Piglets. Front. Microbiol. 2018, 9, 1988. [CrossRef] [PubMed]
155. Díaz Carrasco, J.M.; Redondo, E.A.; Pin Viso, N.D.; Redondo, L.M.; Farber, M.D.; Fernández Miyakawa, M.E. Tannins and
Bacitracin Differentially Modulate Gut Microbiota of Broiler Chickens. BioMed Res. Int. 2018, 2018, 1–11. [CrossRef] [PubMed]
156. Jha, R.; Berrocoso, J.D. Review: Dietary Fiber Utilization and Its Effects on Physiological Functions and Gut Health of Swine.
Animal 2015, 9, 1441–1452. [CrossRef] [PubMed]
157. Schokker, D.; Jansman, A.J.M.; Veninga, G.; de Bruin, N.; Vastenhouw, S.A.; de Bree, F.M.; Bossers, A.; Rebel, J.M.J.; Smits, M.A.
Perturbation of Microbiota in One-Day Old Broiler Chickens with Antibiotic for 24 Hours Negatively Affects Intestinal Immune
Development. BMC Genom. 2017, 18, 241. [CrossRef] [PubMed]
158. Mathlouthi, N.; Mallet, S.; Saulnier, L.; Quemener, B.; Larbier, M. Effects of Xylanase and?—Glucanase Additionon Performance,
Nutrient Digestibility and Physico-Chemical Conditions in the Small Intestine Contents and Caecal Microflora of Broiler Chickens
Feda Wheat and Barley-Based Diet. Anim. Res. 2002, 51, 395–406. [CrossRef]
159. Cherbut, C. Motor Effects of Short-Chain Fatty Acids and Lactate in the Gastrointestinal Tract. Proc. Nutr. Soc. 2003, 62, 95–99.
[CrossRef]
160. Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al.
Enterotypes of the Human Gut Microbiome. Nature 2011, 473, 174–180. [CrossRef]
161. Xu, X.; Yi, H.; Wu, J.; Kuang, T.; Zhang, J.; Li, Q.; Du, H.; Xu, T.; Jiang, G.; Fan, G. Therapeutic Effect of Berberine on Metabolic
Diseases: Both Pharmacological Data and Clinical Evidence. Biomed. Pharmacother. 2021, 133, 110984. [CrossRef] [PubMed]
162. Chen, W.; Miao, Y.-Q.; Fan, D.-J.; Yang, S.-S.; Lin, X.; Meng, L.-K.; Tang, X. Bioavailability Study of Berberine and the Enhancing
Effects of TPGS on Intestinal Absorption in Rats. AAPS PharmSciTech 2011, 12, 705–711. [CrossRef]
163. Wang, Y.; Tong, Q.; Shou, J.-W.; Zhao, Z.-X.; Li, X.-Y.; Zhang, X.-F.; Ma, S.-R.; He, C.-Y.; Lin, Y.; Wen, B.-Y.; et al. Gut MicrobiotaMediated Personalized Treatment of Hyperlipidemia Using Berberine. Theranostics 2017, 7, 2443–2451. [CrossRef]
164. Zhang, H.; Wei, J.; Xue, R.; Wu, J.-D.; Zhao, W.; Wang, Z.-Z.; Wang, S.-K.; Zhou, Z.-X.; Song, D.-Q.; Wang, Y.-M.; et al. Berberine
Lowers Blood Glucose in Type 2 Diabetes Mellitus Patients through Increasing Insulin Receptor Expression. Metabolism 2010, 59,
285–292. [CrossRef] [PubMed]
165. Takahara, M.; Takaki, A.; Hiraoka, S.; Adachi, T.; Shimomura, Y.; Matsushita, H.; Nguyen, T.T.T.; Koike, K.; Ikeda, A.; Takashima,
S.; et al. Berberine Improved Experimental Chronic Colitis by Regulating Interferon-γ- and IL-17A-Producing Lamina Propria
CD4+ T Cells through AMPK Activation. Sci. Rep. 2019, 9, 11934. [CrossRef] [PubMed]
166. Zeng, Q.; Deng, H.; Li, Y.; Fan, T.; Liu, Y.; Tang, S.; Wei, W.; Liu, X.; Guo, X.; Jiang, J.; et al. Berberine Directly Targets the NEK7
Protein to Block the NEK7–NLRP3 Interaction and Exert Anti-Inflammatory Activity. J. Med. Chem. 2021, 64, 768–781. [CrossRef]
167. Ilyas, Z.; Perna, S.; Al-thawadi, S.; Alalwan, T.A.; Riva, A.; Petrangolini, G.; Gasparri, C.; Infantino, V.; Peroni, G.; Rondanelli, M.
The Effect of Berberine on Weight Loss in Order to Prevent Obesity: A Systematic Review. Biomed. Pharmacother. 2020, 127, 110137.
[CrossRef]
168. Ke, X.; Huang, Y.; Li, L.; Xin, F.; Xu, L.; Zhang, Y.; Zeng, Z.; Lin, F.; Song, Y. Berberine Attenuates Arterial Plaque Formation in Atherosclerotic Rats with Damp-Heat Syndrome via Regulating Autophagy. DDDT 2020, 14, 2449–2460. [CrossRef]
169. Tan, W.; Wang, Y.; Wang, K.; Wang, S.; Liu, J.; Qin, X.; Dai, Y.; Wang, X.; Gao, X. Improvement of Endothelial Dysfunction of
Berberine in Atherosclerotic Mice and Mechanism Exploring through TMT-Based Proteomics. Oxid. Med. Cell. Longev. 2020, 2020,
1–22. [CrossRef] [PubMed]
170. Yu, Y.; Zhang, M.; Hu, Y.; Zhao, Y.; Teng, F.; Lv, X.; Li, J.; Zhang, Y.; Hatch, G.M.; Chen, L. Increased Bioavailable Berberine
Protects Against Myocardial Ischemia Reperfusion Injury Through Attenuation of NFκB and JNK Signaling Pathways. Int. Heart
J. 2018, 59, 1378–1388. [CrossRef] [PubMed]
171. Gu, L.; Li, N.; Gong, J.; Li, Q.; Zhu, W.; Li, J. Berberine Ameliorates Intestinal Epithelial Tight-Junction Damage and downRegulates Myosin Light Chain Kinase Pathways in a Mouse Model of Endotoxinemia. J. Infect. Dis. 2011, 203, 1602–1612.
[CrossRef]
172. Gong, J.; Hu, M.; Huang, Z.; Fang, K.; Wang, D.; Chen, Q.; Li, J.; Yang, D.; Zou, X.; Xu, L.; et al. Berberine Attenuates Intestinal
Mucosal Barrier Dysfunction in Type 2 Diabetic Rats. Front. Pharmacol. 2017, 8, 42. [CrossRef]
173. Gu, L.; Li, N.; Li, Q.; Zhang, Q.; Wang, C.; Zhu, W.; Li, J. The Effect of Berberine in Vitro on Tight Junctions in Human Caco-2
Intestinal Epithelial Cells. Fitoterapia 2009, 80, 241–248. [CrossRef]
174. Li, N.; Gu, L.; Qu, L.; Gong, J.; Li, Q.; Zhu, W.; Li, J. Berberine Attenuates Pro-Inflammatory Cytokine-Induced Tight Junction
Disruption in an in Vitro Model of Intestinal Epithelial Cells. Eur. J. Pharm. Sci. 2010, 40, 1–8. [CrossRef] [PubMed]
175. Fernandez, C.P.; Afrin, F.; Flores, R.A.; Kim, W.H.; Jeong, J.; Kim, S.; Chang, H.H.; Lillehoj, H.S.; Min, W. Downregulation of
Inflammatory Cytokines by Berberine Attenuates Riemerella anatipestifer Infection in Ducks. Dev. Comp. Immunol. 2017, 77,
121–127. [CrossRef] [PubMed]
176. Habtemariam, S. Berberine Pharmacology and the Gut Microbiota: A Hidden Therapeutic Link. Pharmacol. Res. 2020, 155, 104722.
[CrossRef] [PubMed]
177. Lyu, Y.; Lin, L.; Xie, Y.; Li, D.; Xiao, M.; Zhang, Y.; Cheung, S.C.K.; Shaw, P.C.; Yang, X.; Chan, P.K.S.; et al. Blood-Glucose-Lowering
Effect of Coptidis Rhizoma Extracts from Different Origins via Gut Microbiota Modulation in Db/Db Mice. Front. Pharmacol.
2021, 12, 684358. [CrossRef]
178. Zhu, C.; Huang, K.; Bai, Y.; Feng, X.; Gong, L.; Wei, C.; Huang, H.; Zhang, H. Dietary Supplementation with Berberine Improves
Growth Performance and Modulates the Composition and Function of Cecal Microbiota in Yellow-Feathered Broilers. Poult. Sci.
2021, 100, 1034–1048. [CrossRef]
179. Xiang Yu, D.; He, Z.; Pouton, C.; Hoerr, F.J.; Xiao, Z.C. Target Animal Safety and Residual Study for Berberine and Other
Phytogenic Compounds in Broiler Chickens. Arch. Clin. MicroBiol. 2017, 8, 69. [CrossRef]
180. Malik, T.A.; Kamili, A.N.; Chishti, M.Z.; Tanveer, S.; Ahad, S.; Johri, R.K. Synergistic Approach for Treatment of Chicken
Coccidiosis Using Berberine—A Plant Natural Product. Microb. Pathog. 2016, 93, 56–62. [CrossRef]
181. Yang, L.; Liu, G.; Liang, X.; Wang, M.; Zhu, X.; Luo, Y.; Shang, Y.; Yang, J.; Zhou, P.; Gu, X. Effects of Berberine on the Growth
Performance, Antioxidative Capacity and Immune Response to Lipopolysaccharide Challenge in Broilers. Anim. Sci. J. 2019, 90,
1229–1238. [CrossRef]
182. Shen, Y.B.; Piao, X.S.; Kim, S.W.; Wang, L.; Liu, P. The Effects of Berberine on the Magnitude of the Acute Inflammatory Response
Induced by Escherichia Coli Lipopolysaccharide in Broiler Chickens. Poult. Sci. 2010, 89, 13–19. [CrossRef]
183. Zhang, Y.; Guo, L.; Huang, J.; Sun, Y.; He, F.; Zloh, M.; Wang, L. Inhibitory Effect of Berberine on Broiler P-Glycoprotein Expression and Function: In Situ and In Vitro Studies. IJMS 2019, 20, 1966. [CrossRef]
184. Frank, M.B.; Yang, Q.; Osban, J.; Azzarello, J.T.; Saban, M.R.; Saban, R.; Ashley, R.A.; Welter, J.C.; Fung, K.-M.; Lin, H.-K.
Frankincense Oil Derived from Boswellia Carteri Induces Tumor Cell Specific Cytotoxicity. BMC Complement. Altern. Med. 2009,
9, 6. [CrossRef]
185. Poeckel, D.; Werz, O. Boswellic Acids: Biological Actions and Molecular Targets. Curr. Med. Chem. 2006, 13, 3359–3369. [CrossRef]
186. Anthoni, C.; Laukoetter, M.G.; Rijcken, E.; Vowinkel, T.; Mennigen, R.; Müller, S.; Senninger, N.; Russell, J.; Jauch, J.; Bergmann,
J.; et al. Mechanisms Underlying the Anti-Inflammatory Actions of Boswellic Acid Derivatives in Experimental Colitis. Am. J.
Physiol. Gastrointest. Liver Physiol. 2006, 290, G1131–G1137. [CrossRef] [PubMed]
187. Ammon, H.P.T. Modulation of the Immune System by Boswellia Serrata Extracts and Boswellic Acids. Phytomedicine 2010, 17,
862–867. [CrossRef] [PubMed]
188. Abdel-Tawab, M.; Werz, O.; Schubert-Zsilavecz, M. Boswellia serrata: An Overall Assessment of in Vitro, Preclinical, Pharmacokinetic and Clinical Data. Clin. Pharmacokinet. 2011, 50, 349–369. [CrossRef] [PubMed]
189. Catanzaro, D.; Rancan, S.; Orso, G.; Dall’Acqua, S.; Brun, P.; Giron, M.C.; Carrara, M.; Castagliuolo, I.; Ragazzi, E.; Caparrotta, L.; et al. Boswellia Serrata Preserves Intestinal Epithelial Barrier from Oxidative and Inflammatory Damage. PLoS ONE 2015, 10, e0125375. [CrossRef]
190. Ismail, I.E.; Abdelnour, S.A.; Shehata, S.A.; Abd El-Hack, M.E.; El-Edel, M.A.; Taha, A.E.; Schiavitto, M.; Tufarelli, V. Effect of
Dietary Boswellia Serrata Resin on Growth Performance, Blood Biochemistry, and Cecal Microbiota of Growing Rabbits. Front.
Vet. Sci. 2019, 6, 471. [CrossRef]
191. Mohamed, S.H.; Attia, A.I.; Reda, F.M.; Abd El-Hack, M.E.; Ismail, I.E. Impacts of Dietary Supplementation of Boswellia Serrata on
Growth, Nutrients Digestibility, Immunity, Antioxidant Status, Carcase Traits and Caecum Microbiota of Broilers. Ital. J. Anim.
Sci. 2021, 20, 205–214. [CrossRef]
192. Al-Yasiry, A.R.M.; Kiczorowska, B.; Samoli ´nska, W.; Kowalczuk-Vasilev, E.; Kowalczyk-Pecka, D. The Effect of Boswellia Serrata
Resin Diet Supplementation on Production, Hematological, Biochemical and Immunological Parameters in Broiler Chickens.
Animal 2017, 11, 1890–1898. [CrossRef]
193. Alvarez-Parrilla, E.; de la Rosa, L.A.; Amarowicz, R.; Shahidi, F. Antioxidant Activity of Fresh and Processed Jalapeño and
Serrano Peppers. J. Agric. Food Chem. 2011, 59, 163–173. [CrossRef]
194. Zhuang, Y.; Chen, L.; Sun, L.; Cao, J. Bioactive Characteristics and Antioxidant Activities of Nine Peppers. J. Funct. Foods 2012, 4,
331–338. [CrossRef]
195. Santos, E.A.D.; Alvarez-Leite, J.I. Capsaicin: A Potential Therapy Adjuvant for Intestinal Bowel Disease. JDDD 2019, 2, 8–16.
[CrossRef]
196. Kang, C.; Zhang, Y.; Zhu, X.; Liu, K.; Wang, X.; Chen, M.; Wang, J.; Chen, H.; Hui, S.; Huang, L.; et al. Healthy Subjects
Differentially Respond to Dietary Capsaicin Correlating with Specific Gut Enterotypes. J. Clin. Endocrinol. Metab. 2016, 101,
4681–4689. [CrossRef] [PubMed]
197. Mason, J.R.; Maruniak, J.A. Behavioral and Physiological Effects of Capsaicin in Red-Winged Blackbirds. Pharmacol. Biochem.
Behav. 1983, 19, 857–862. [CrossRef]
198. Geisthövel, E.; Ludwig, O.; Simon, E. Capsaicin Fails to Produce Disturbances of Autonomic Heat and Cold Defence in an Avian
Species (Anas platyrhynchos). Pflugers. Arch. 1986, 406, 343–350. [CrossRef]
199. Szolcsányi, J.; Jancsó-Gábor, A. Sensory Effects of Capsaicin Congeners I. Relationship between Chemical Structure and PainProducing Potency of Pungent Agents. Arzneimittelforschung 1975, 25, 1877–1881.
200. Muley, B.; Khadabadi, S.; Banarase, N. Phytochemical Constituents and Pharmacological Activities of Calendula Officinalis Linn (Asteraceae): A Review. Trop. J. Pharm. Res. 2009, 8. [CrossRef]
201. Ukiya, M.; Akihisa, T.; Yasukawa, K.; Tokuda, H.; Suzuki, T.; Kimura, Y. Anti-Inflammatory, Anti-Tumor-Promoting, and
Cytotoxic Activities of Constituents of Marigold (Calendula officinalis) Flowers. J. Nat. Prod. 2006, 69, 1692–1696. [CrossRef]
202. Yoshikawa, M.; Murakami, T.; Kishi, A.; Kageura, T.; Matsuda, H. Medicinal Flowers. III. Marigold. (1): Hypoglycemic, Gastric
Emptying Inhibitory, and Gastroprotective Principles and New Oleanane-Type Triterpene Oligoglycosides, Calendasaponins A,
B, C, and D, from Egyptian Calendula Officinalis. Chem. Pharm. Bull. 2001, 49, 863–870. [CrossRef]
203. Zitterl-Eglseer, K.; Sosa, S.; Jurenitsch, J.; Schubert-Zsilavecz, M.; Della Loggia, R.; Tubaro, A.; Bertoldi, M.; Franz, C. AntiOedematous Activities of the Main Triterpendiol Esters of Marigold (Calendula officinalis L.). J. Ethnopharmacol. 1997, 57, 139–144.
[CrossRef]
204. Kalvatchev, Z.; Walder, R.; Garzaro, D. Anti-HIV Activity of Extracts from Calendula officinalis Flowers. Biomed. Pharmacother. 1997,
51, 176–180. [CrossRef]
205. Dall’Acqua, S.; Catanzaro, D.; Cocetta, V.; Igl, N.; Ragazzi, E.; Giron, M.C.; Cecconello, L.; Montopoli, M. Protective Effects of ψ
Taraxasterol 3-O-Myristate and Arnidiol 3-O-Myristate Isolated from Calendula officinalis on Epithelial Intestinal Barrier. Fitoterapia
2016, 109, 230–235. [CrossRef] [PubMed]
206. Foroutankhah, M.; Toghyani, M.; Landy, N. Evaluation of Calendula officinalis L. (Marigold) Flower as a Natural Growth Promoter in Comparison with an Antibiotic Growth Promoter on Growth Performance, Carcass Traits and Humoral Immune Responses of
Broilers. Anim. Nutr. 2019, 5, 314–318. [CrossRef]
207. Rajput, N.; Naeem, M.; Ali, S.; Rui, Y.; Tian, W. Effect of Dietary Supplementation of Marigold Pigment on Immunity, Skin and
Meat Color, and Growth Performance of Broiler Chickens. Braz. J. Poult. Sci. 2012, 4, 233–304. [CrossRef]
208. Page, J.E.; Nagel, J. Chapter Eight Biosynthesis of Terpenophenolic Metabolites in Hop and Cannabis. In Recent Advances in Phytochemistry; Elsevier: Amsterdam, The Netherlands, 2006.
209. Klein, T.W.; Cabral, G.A. Cannabinoid-Induced Immune Suppression and Modulation of Antigen-Presenting Cells. J. Neuroimmune
Pharmacol. 2006, 1, 50–64. [CrossRef]
210. Izzo, A.A.; Sharkey, K.A. Cannabinoids and the Gut: New Developments and Emerging Concepts. Pharmacol. Ther. 2010, 126,
21–38. [CrossRef] [PubMed]
211. Konieczka, P.; Szkopek, D.; Kinsner, M.; Fotschki, B.; Ju´skiewicz, J.; Banach, J. Cannabis-Derived Cannabidiol and Nanoselenium
Improve Gut Barrier Function and Affect Bacterial Enzyme Activity in Chickens Subjected to C. Perfringens Challenge. Vet. Res.
2020, 51, 141. [CrossRef]
212. Alhamoruni, A.; Lee, A.C.; Wright, K.L.; Larvin, M.; O’Sullivan, S.E. Pharmacological Effects of Cannabinoids on the Caco-2 Cell
Culture Model of Intestinal Permeability. J. Pharmacol. Exp. Ther. 2010, 335, 92–102. [CrossRef]
213. Barceloux, D. Medical Toxicology of Natural Substances: Foods, Fungi, Medicinal Herbs, Plants and Venomous Animals; Wiley: Hoboken,
NJ, USA, 2008.
214. Han, X.; Parker, T.L. Anti-Inflammatory Activity of Clove (Eugenia caryophyllata) Essential Oil in Human Dermal Fibroblasts.
Pharm. Biol. 2017, 55, 1619–1622. [CrossRef]
215. Pirgozliev, V.; Rose, S.; Catherine, I.; Blanchard, A. Phytogenic Feed Additives Can Alleviate the Negative Impact of Necrotic
Enteritis in Broilers. In Proceedings of the 6th International Conference on Poultry Intestinal Health, Rome, Italy, 3–5 April 2019.
216. Kumar, A.; Kheravii, S.K.; Ionescu, C.; Blanchard, A.; Barekatain, R.; Bajagai, Y.S.; Wu, S.-B. A Microencapsulated Mixture of
Eugenol and Garlic Tincture Supplementation Mitigates the Effect of Necrotic Enteritis on Intestinal Integrity and Increases
Goblet Cells in Broilers. Microorganisms 2021, 9, 1451. [CrossRef]
217. Ajdžanovi´c, V.Z.; Medigovi´c, I.M.; Panteli´c, J.B.; Miloševi´c, V.Lj. Soy Isoflavones and Cellular Mechanics. J. Bioenerg. Biomembr.
2014, 46, 99–107. [CrossRef]
218. Jiang, Z.Y.; Jiang, S.Q.; Lin, Y.C.; Xi, P.B.; Yu, D.Q.; Wu, T.X. Effects of Soybean Isoflavone on Growth Performance, Meat Quality, and Antioxidation in Male Broilers. Poult. Sci. 2007, 86, 1356–1362. [CrossRef] [PubMed]
219. Zhang, Y.-Z.; Shi, D.-X.; Zhou, B.-C.; Zeng, C.-K.; Pang, S.-J. Study on the Structure of C-Phycocyanin in Spirulina Platensis with
Scanning Tunneling Microscope. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao 1997, 29, 521–525. [PubMed]
220. Messina, M.; Ho, S.; Alekel, D.L. Skeletal Benefits of Soy Isoflavones: A Review of the Clinical Trial and Epidemiologic Data. Curr.
Opin. Clin. Nutr. Metab. Care 2004, 7, 649–658. [CrossRef] [PubMed]
221. Iqbal, Y.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. Gut Microbiota-Polyphenol Interactions in Chicken: A Review. Animals 2020,
10, 1391. [CrossRef]
222. Cassidy, A.; Brown, J.E.; Hawdon, A.; Faughnan, M.S.; King, L.J.; Millward, J.; Zimmer-Nechemias, L.; Wolfe, B.; Setchell,
K.D.R. Factors Affecting the Bioavailability of Soy Isoflavones in Humans after Ingestion of Physiologically Relevant Levels from
Different Soy Foods. J. Nutr. 2006, 136, 45–51. [CrossRef]
223. Azzam, M.M.; Jiang, S.; Chen, J.; Lin, X.; Gou, Z.; Fan, Q.; Wang, Y.; Li, L.; Jiang, Z. Effect of Soybean Isoflavones on Growth
Performance, Immune Function, and Viral Protein 5 MRNA Expression in Broiler Chickens Challenged with Infectious Bursal
Disease Virus. Animals 2019, 9, 247. [CrossRef]
224. Cassidy, A. Potential Risks and Benefits of Phytoestrogen-Rich Diets. Int. J. Vitam. Nutr. Res. 2003, 73, 120–126. [CrossRef]
225. Yousef, M.I.; Esmail, A.M.; Baghdadi, H.H. Effect of Isoflavones on Reproductive Performance, Testosterone Levels, Lipid
Peroxidation, and Seminal Plasma Biochemistry of Male Rabbits. J. Environ. Sci. Health B 2004, 39, 819–833. [CrossRef]
226. Shin, J.-H.; Park, J.-M.; Bak, D.-J.; Jean, W.-M.; Song, J.-C.; Kim, S.-K.; An, B.-K.; Kang, C.-W.; Jung, W.-S.; Kim, J.-M. Effects of
Germinated and Fermented Unmarketable Soybean on Laying Performance and Egg Quality in Laying Hens. Korean J. Food Sci.
Anim. Resour. 2008, 28, 667–674. [CrossRef]
227. Shi, S.R.; Gu, H.; Chang, L.L.; Wang, Z.Y.; Tong, H.B.; Zou, J.M. Safety Evaluation of Daidzein in Laying Hens: Part I. Effects on
Laying Performance, Clinical Blood Parameters, and Organs Development. Food Chem. Toxicol. 2013, 55, 684–688. [CrossRef]
228. Zhao, X.; Shao, T.; Wang, Y.Q.; Lu, X.L.; Luo, J.B.; Zhou, W.D. The Phytoestrogen Daidzein May Affect Reproductive Performance of Zhedong White Geese by Regulating Gene MRNA Levels in the HPG Axis. Br. Poult. Sci. 2013, 54, 252–258. [CrossRef]
[PubMed]
229. Zhao, R.Q.; Zhou, Y.C.; Ni, Y.D.; Lu, L.Z.; Tao, Z.R.; Chen, W.H.; Chen, J. Effect of Daidzein on Egg-Laying Performance in Shaoxing Duck Breeders during Different Stages of the Egg Production Cycle. Br. Poult. Sci. 2005, 46, 175–181. [CrossRef]
[PubMed]
230. Ni, Y.D.; Wu, J.; Tong, H.Y.; Huang, Y.B.; Lu, L.Z.; Grossmann, R.; Zhao, R.Q. Effect of Dietary Daidzein Supplementation on Egg
Laying Rate Was Associated with the Change of Hepatic VTG-II MRNA Expression and Higher Antioxidant Activities during the Post-Peak Egg Laying Period of Broiler Breeders. Anim. Feed. Sci. Technol. 2012, 177, 116–123. [CrossRef]
231. Sahin, N.; Onderci, M.; Balci, T.A.; Cikim, G.; Sahin, K.; Kucuk, O. The Effect of Soy Isoflavones on Egg Quality and Bone
Mineralisation during the Late Laying Period of Quail. Br. Poult. Sci. 2007, 48, 363–369. [CrossRef]
232. Kajiya, H.; Okabe, K.; Okamoto, F.; Tsuzuki, T.; Soeda, H. Protein Tyrosine Kinase Inhibitors Increase Cytosolic Calcium and
Inhibit Actin Organization as Resorbing Activity in Rat Osteoclasts. J. Cell Physiol. 2000, 183, 83–90. [CrossRef]
233. Kosina, P.; Gregorova, J.; Gruz, J.; Vacek, J.; Kolar, M.; Vogel, M.; Roos, W.; Naumann, K.; Simanek, V.; Ulrichova, J. Phytochemical and Antimicrobial Characterization of Macleaya Cordata Herb. Fitoterapia 2010, 81, 1006–1012. [CrossRef]
234. Le, H.H.; Shakeri, M.; Suleria, H.A.R.; Zhao, W.; McQuade, R.M.; Phillips, D.J.; Vidacs, E.; Furness, J.B.; Dunshea, F.R.; ArtusoPonte, V.; et al. Betaine and Isoquinoline Alkaloids Protect against Heat Stress and Colonic Permeability in Growing Pigs.
Antioxidants 2020, 9, 1024. [CrossRef]
235. Vrublova, E.; Vostalova, J.; Ehrmann, J.; Palikova, I.; Vrbkova, J.; Vacek, J.; Cibicek, N.; Vecera, R.; Ulrichova, J.; Simanek, V.
The Phytogenetic Feed Additive Sangrovit Modulates Dextran Sulfate Sodium-Induced Colitis in Rats. Vet. Med. 2010, 55,
610–618. [CrossRef]
236. Robbins, R.C.; Artuso-Ponte, V.C.; Moeser, A.J.; Morrow, W.E.M.; Spears, J.W.; Gebreyes, W.A. Effects of Quaternary
Benzo(c)Phenanthridine Alkaloids on Growth Performance, Shedding of Organisms, and Gastrointestinal Tract Integrity in Pigs
Inoculated with Multidrug-Resistant Salmonella Spp. Am. J. Vet. Res. 2013, 74, 1530–1535. [CrossRef]
237. Liu, W.; Zhai, Y.; Heng, X.; Che, F.Y.; Chen, W.; Sun, D.; Zhai, G. Oral Bioavailability of Curcumin: Problems and Advancements.
J. Drug Target 2016, 24, 694–702. [CrossRef] [PubMed]
238. El, S.N.; Karakaya, S. Olive Tree (Olea europaea) Leaves: Potential Beneficial Effects on Human Health. Nutr. Rev. 2009, 67, 632–638.
[CrossRef]
239. Talhaoui, N.; Vezza, T.; Gómez-Caravaca, A.M.; Fernández-Gutiérrez, A.; Gálvez, J.; Segura-Carretero, A. Phenolic Compounds and in Vitro Immunomodulatory Properties of Three Andalusian Olive Leaf Extracts. J. Funct. Foods 2016, 22, 270–277. [CrossRef]
240. Farràs, M.; Martinez-Gili, L.; Portune, K.; Arranz, S.; Frost, G.; Tondo, M.; Blanco-Vaca, F. Modulation of the Gut Microbiota by Olive Oil Phenolic Compounds: Implications for Lipid Metabolism, Immune System, and Obesity. Nutrients 2020, 12, 2200.
[CrossRef]
241. Deiana, M.; Serra, G.; Corona, G. Modulation of Intestinal Epithelium Homeostasis by Extra Virgin Olive Oil Phenolic Compounds.
Food Funct. 2018, 9, 4085–4099. [CrossRef] [PubMed]
242. Millman, J.; Okamoto, S.; Kimura, A.; Uema, T.; Higa, M.; Yonamine, M.; Namba, T.; Ogata, E.; Yamazaki, S.; Shimabukuro,
M.; et al. Metabolically and Immunologically Beneficial Impact of Extra Virgin Olive and Flaxseed Oils on Composition of Gut
Microbiota in Mice. Eur. J. Nutr. 2020, 59, 2411–2425. [CrossRef]
243. ¸Senay, S.; Dursen, Ü. The Use of Grape Seed-, Olive Leaf- and Pomegranate Peel-Extracts as Alternative Natural Antimicrobial Feed
Additives in Broiler Diets; Verlag Eugen Ulmer: Stuttgart, Germany, 2016.
244. Herrero-Encinas, J.; Blanch, M.; Pastor, J.J.; Mereu, A.; Ipharraguerre, I.R.; Menoyo, D. Effects of a Bioactive Olive Pomace Extract from Olea Europaea on Growth Performance, Gut Function, and Intestinal Microbiota in Broiler Chickens. Poult. Sci. 2020, 99,
2–10. [CrossRef] [PubMed]
245. Liu, Z.; Wang, N.; Ma, Y.; Wen, D. Hydroxytyrosol Improves Obesity and Insulin Resistance by Modulating Gut Microbiota in High-Fat Diet-Induced Obese Mice. Front. Microbiol. 2019, 10, 390. [CrossRef]
246. Vezza, T.; Algieri, F.; Rodríguez-Nogales, A.; Garrido-Mesa, J.; Utrilla, M.P.; Talhaoui, N.; Gómez-Caravaca, A.M.; SeguraCarretero, A.; Rodríguez-Cabezas, M.E.; Monteleone, G.; et al. Immunomodulatory Properties of Olea Europaea Leaf Extract in Intestinal Inflammation. Mol. Nutr. Food Res. 2017, 61, 1601066. [CrossRef]
247. Hertog, M.G.L.; Hollman, P.C.H.; Katan, M.B. Content of Potentially Anticarcinogenic Flavonoids of 28 Vegetables and 9 Fruits
Commonly Consumed in the Netherlands. J. Agric. Food Chem. 1992, 40, 2379–2383. [CrossRef]
248. Erlund, I. Review of the Flavonoids Quercetin, Hesperetin, and Naringenin. Dietary Sources, Bioactivities, Bioavailability, and
Epidemiology. Nutr. Res. 2004, 24, 851–874. [CrossRef]
249. Tasdemir, D.; Lack, G.; Brun, R.; Rüedi, P.; Scapozza, L.; Perozzo, R. Inhibition of Plasmodium f Alciparum Fatty Acid Biosynthesis:
Evaluation of FabG, FabZ, and FabI as Drug Targets for Flavonoids. J. Med. Chem. 2006, 49, 3345–3353. [CrossRef]
250. Wu, D.; Kong, Y.; Han, C.; Chen, J.; Hu, L.; Jiang, H.; Shen, X. D-Alanine:D-Alanine Ligase as a New Target for the Flavonoids
Quercetin and Apigenin. Int. J. Antimicrob. Agents 2008, 32, 421–426. [CrossRef] [PubMed]
251. Abdel-Latif, M.A.; Elbestawy, A.R.; El-Far, A.H.; Noreldin, A.E.; Emam, M.; Baty, R.S.; Albadrani, G.M.; Abdel-Daim, M.M.;
Abd El-Hamid, H.S. Quercetin Dietary Supplementation Advances Growth Performance, Gut Microbiota, and Intestinal MRNA
Expression Genes in Broiler Chickens. Animals 2021, 11, 2302. [CrossRef] [PubMed]
252. Suzuki, T.; Hara, H. Quercetin Enhances Intestinal Barrier Function through the Assembly of Zonula [Corrected] Occludens-2,
Occludin, and Claudin-1 and the Expression of Claudin-4 in Caco-2 Cells. J. Nutr. 2009, 139, 965–974. [CrossRef]
253. Amasheh, M.; Grotjohann, I.; Amasheh, S.; Fromm, A.; Söderholm, J.D.; Zeitz, M.; Fromm, M.; Schulzke, J.-D. Regulation of
Mucosal Structure and Barrier Function in Rat Colon Exposed to Tumor Necrosis Factor Alpha and Interferon Gamma in Vitro:
A Novel Model for Studying the Pathomechanisms of Inflammatory Bowel Disease Cytokines. Scand. J. Gastroenterol. 2009, 44,
1226–1235. [CrossRef]
254. Carrasco-Pozo, C.; Morales, P.; Gotteland, M. Polyphenols Protect the Epithelial Barrier Function of Caco-2 Cells Exposed to
Indomethacin through the Modulation of Occludin and Zonula Occludens-1 Expression. J. Agric. Food Chem. 2013, 61, 5291–5297.
[CrossRef]
255. Sim, G.-S.; Lee, B.-C.; Cho, H.S.; Lee, J.W.; Kim, J.-H.; Lee, D.-H.; Kim, J.-H.; Pyo, H.-B.; Moon, D.C.; Oh, K.-W.; et al. Structure
Activity Relationship of Antioxidative Property of Flavonoids and Inhibitory Effect on Matrix Metalloproteinase Activity in UVA-Irradiated Human Dermal Fibroblast. Arch. Pharm. Res. 2007, 30, 290–298. [CrossRef]
256. Agullo, G.; Gamet-Payrastre, L.; Manenti, S.; Viala, C.; Rémésy, C.; Chap, H.; Payrastre, B. Relationship between Flavonoid
Structure and Inhibition of Phosphatidylinositol 3-Kinase: A Comparison with Tyrosine Kinase and Protein Kinase C Inhibition.
Biochem. Pharmacol. 1997, 53, 1649–1657. [CrossRef]
257. Fachini-Queiroz, F.C.; Kummer, R.; Estevão-Silva, C.F.; Carvalho, M.D.D.B.; Cunha, J.M.; Grespan, R.; Bersani-Amado, C.A.;
Cuman, R.K.N. Effects of Thymol and Carvacrol, Constituents of Thymus vulgaris L. Essential Oil, on the Inflammatory Response.
Evid. Based Complement Alternat. Med. 2012, 2012, 657026. [CrossRef]
258. Hosseinzadeh, S.; Jafarikukhdan, A.; Hosseini, A.; Armand, R. The Application of Medicinal Plants in Traditional and Modern
Medicine: A Review of Thymus vulgaris. IJCM 2015, 06, 635–642. [CrossRef]
259. Sakkas, H.; Papadopoulou, C. Antimicrobial Activity of Basil, Oregano, and Thyme Essential Oils. J. MicroBiol. Biotechnol. 2017,
27, 429–438. [CrossRef] [PubMed]
260. Yalçin, S.; Eser, H.; Onba¸silar, ˙I.; Yalçin, S. Effects of Dried Thyme (Thymus vulgaris L.) Leaves on Performance, Some Egg Quality
Traits and Immunity in Laying Hens. Ank. Üniv. Vet. Fakültesi Derg. 2020, 67, 303–311. [CrossRef]
261. Turner, J.R. Molecular Basis of Epithelial Barrier Regulation: From Basic Mechanisms to Clinical Application. Am. J. Pathol. 2006,
169, 1901–1909. [CrossRef]
262. Placha, I.; Chrastinova, L.; Laukova, A.; Cobanova, K.; Takacova, J.; Strompfova, V.; Chrenkova, M.; Formelova, Z.; Faix, S. Effect of Thyme Oil on Small Intestine Integrity and Antioxidant Status, Phagocytic Activity and Gastrointestinal Microbiota in Rabbits.
Acta Vet. Hung. 2013, 61, 197–208. [CrossRef] [PubMed]
263. Yoshino, K.; Higashi, N.; Koga, K. Antioxidant and Antiinflammatory Activities of Oregano Extract. J. Health Sci. 2006, 52,
169–173. [CrossRef]
264. Han, X.; Parker, T.L. Anti-Inflammatory, Tissue Remodeling, Immunomodulatory, and Anticancer Activities of Oregano (Origanum vulgare) Essential Oil in a Human Skin Disease Model. Biochim. Open 2017, 4, 73–77. [CrossRef] [PubMed]
265. Avola, R.; Granata, G.; Geraci, C.; Napoli, E.; Graziano, A.C.E.; Cardile, V. Oregano (Origanum vulgare L.) Essential Oil Provides
Anti-Inflammatory Activity and Facilitates Wound Healing in a Human Keratinocytes Cell Model. Food Chem. Toxicol. 2020, 144,
111586. [CrossRef]
266. Du, E.; Wang, W.; Gan, L.; Li, Z.; Guo, S.; Guo, Y. Effects of Thymol and Carvacrol Supplementation on Intestinal Integrity and
Immune Responses of Broiler Chickens Challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2016, 7, 19. [CrossRef]
267. Pham, V.H.; Kan, L.; Huang, J.; Geng, Y.; Zhen, W.; Guo, Y.; Abbas, W.; Wang, Z. Dietary Encapsulated Essential Oils and Organic
Acids Mixture Improves Gut Health in Broiler Chickens Challenged with Necrotic Enteritis. J. Anim. Sci. Biotechnol. 2020, 11, 18.
[CrossRef] [PubMed]
268. Hashemipour, H.; Kermanshahi, H.; Golian, A.; Veldkamp, T. Effect of Thymol and Carvacrol Feed Supplementation on
Performance, Antioxidant Enzyme Activities, Fatty Acid Composition, Digestive Enzyme Activities, and Immune Response in Broiler Chickens. Poult. Sci. 2013, 92, 2059–2069. [CrossRef] [PubMed]
269. Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential Oil and Aromatic Plants as Feed Additives in Non-Ruminant Nutrition: A Review.
J. Anim. Sci. Biotechnol. 2015, 6, 7. [CrossRef]
270. Feng, J.; Lu, M.; Wang, J.; Zhang, H.; Qiu, K.; Qi, G.; Wu, S. Dietary Oregano Essential Oil Supplementation Improves Intestinal
Functions and Alters Gut Microbiota in Late-Phase Laying Hens. J. Anim. Sci. Biotechnol. 2021, 12, 72. [CrossRef] [PubMed]
271. Zhang, C.; Zhao, X.H.; Yang, L.; Chen, X.Y.; Jiang, R.S.; Jin, S.H.; Geng, Z.Y. Resveratrol Alleviates Heat Stress-Induced Impairment of Intestinal Morphology, Microflora, and Barrier Integrity in Broilers. Poult. Sci. 2017, 96, 4325–4332. [CrossRef] [PubMed]
272. Madeo, F.; Carmona-Gutierrez, D.; Hofer, S.J.; Kroemer, G. Caloric Restriction Mimetics against Age-Associated Disease: Targets,
Mechanisms, and Therapeutic Potential. Cell Metab. 2019, 29, 592–610. [CrossRef] [PubMed]
273. Manna, S.K.; Mukhopadhyay, A.; Aggarwal, B.B. Resveratrol Suppresses TNF-Induced Activation of Nuclear Transcription
Factors NF-Kappa B, Activator Protein-1, and Apoptosis: Potential Role of Reactive Oxygen Intermediates and Lipid Peroxidation.
J. Immunol. 2000, 164, 6509–6519. [CrossRef]
274. Liu, L.L.; He, J.H.; Xie, H.B.; Yang, Y.S.; Li, J.C.; Zou, Y. Resveratrol Induces Antioxidant and Heat Shock Protein MRNA
Expression in Response to Heat Stress in Black-Boned Chickens. Poult. Sci. 2014, 93, 54–62. [CrossRef]
275. Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol Improves Mitochondrial Function and Protects against Metabolic Disease by Activating SIRT1 and PGC-1alpha.
Cell 2006, 127, 1109–1122. [CrossRef]
276. Mayangsari, Y.; Suzuki, T. Resveratrol Enhances Intestinal Barrier Function by Ameliorating Barrier Disruption in Caco-2 Cell
Monolayers. J. Funct. Foods 2018, 51, 39–46. [CrossRef]
277. Zhao, W.; Huang, X.; Han, X.; Hu, D.; Hu, X.; Li, Y.; Huang, P.; Yao, W. Resveratrol Suppresses Gut-Derived NLRP3 Inflammasome
Partly through Stabilizing Mast Cells in a Rat Model. Mediat. Inflamm. 2018, 2018, 1–10. [CrossRef]
278. Zhang, C.; Yang, L.; Zhao, X.; Chen, X.; Wang, L.; Geng, Z. Effect of Dietary Resveratrol Supplementation on Meat Quality, Muscle
Antioxidative Capacity and Mitochondrial Biogenesis of Broilers. J. Sci. Food Agric. 2018, 98, 1216–1221. [CrossRef] [PubMed]
279. Wang, C.; Zhao, F.; Li, Z.; Jin, X.; Chen, X.; Geng, Z.; Hu, H.; Zhang, C. Effects of Resveratrol on Growth Performance, Intestinal
Development, and Antioxidant Status of Broilers under Heat Stress. Animals 2021, 11, 1427. [CrossRef] [PubMed]
280. Pan, M.-H.; Wu, J.-C.; Ho, C.-T.; Lai, C.-S. Antiobesity Molecular Mechanisms of Action: Resveratrol and Pterostilbene. Biofactors
2018, 44, 50–60. [CrossRef] [PubMed]
281. Cai, T.-T.; Ye, X.-L.; Li, R.-R.; Chen, H.; Wang, Y.-Y.; Yong, H.-J.; Pan, M.-L.; Lu, W.; Tang, Y.; Miao, H.; et al. Resveratrol
Modulates the Gut Microbiota and Inflammation to Protect Against Diabetic Nephropathy in Mice. Front. Pharmacol. 2020, 11,
1249. [CrossRef] [PubMed]
282. Alrafas, H.R.; Busbee, P.B.; Nagarkatti, M.; Nagarkatti, P.S. Resveratrol Modulates the Gut Microbiota to Prevent Murine Colitis
Development through Induction of Tregs and Suppression of Th17 Cells. J. Leukoc. Biol. 2019, 106, 467–480. [CrossRef] [PubMed]
283. Kocaadam, B.; ¸Sanlier, N. Curcumin, an Active Component of Turmeric (Curcuma Longa), and Its Effects on Health. Crit. Rev.
Food Sci. Nutr. 2017, 57, 2889–2895. [CrossRef] [PubMed]
284. Tsuda, T. Curcumin as a Functional Food-Derived Factor: Degradation Products, Metabolites, Bioactivity, and Future Perspectives.
Food Funct. 2018, 9, 705–714. [CrossRef] [PubMed]
285. Asai, A.; Miyazawa, T. Occurrence of Orally Administered Curcuminoid as Glucuronide and Glucuronide/Sulfate Conjugates in Rat Plasma. Life Sci. 2000, 67, 2785–2793. [CrossRef]
286. Ireson, C.R.; Jones, D.J.L.; Orr, S.; Coughtrie, M.W.H.; Boocock, D.J.; Williams, M.L.; Farmer, P.B.; Steward, W.P.; Gescher, A.J.
Metabolism of the Cancer Chemopreventive Agent Curcumin in Human and Rat Intestine. Cancer Epidemiol. Biomark. Prev. 2002,
11, 105–111.
287. Pan, M.H.; Huang, T.M.; Lin, J.K. Biotransformation of Curcumin through Reduction and Glucuronidation in Mice. Drug Metab.
Dispos. 1999, 27, 486–494.
288. Stohs, S.J.; Chen, O.; Ray, S.D.; Ji, J.; Bucci, L.R.; Preuss, H.G. Highly Bioavailable Forms of Curcumin and Promising Avenues for
Curcumin-Based Research and Application: A Review. Molecules 2020, 25, 1397. [CrossRef]
289. Shehzad, A.; Wahid, F.; Lee, Y.S. Curcumin in Cancer Chemoprevention: Molecular Targets, Pharmacokinetics, Bioavailability, and Clinical Trials. Arch. Pharm. 2010, 343, 489–499. [CrossRef] [PubMed]
290. Han, H.-K. The Effects of Black Pepper on the Intestinal Absorption and Hepatic Metabolism of Drugs. Expert Opin. Drug Metab.
Toxicol. 2011, 7, 721–729. [CrossRef] [PubMed]
291. Shaikh, J.; Ankola, D.D.; Beniwal, V.; Singh, D.; Kumar, M.N.V.R. Nanoparticle Encapsulation Improves Oral Bioavailability of
Curcumin by at Least 9-Fold When Compared to Curcumin Administered with Piperine as Absorption Enhancer. Eur. J. Pharm.
Sci. 2009, 37, 223–230. [CrossRef]
292. Itokawa, H.; Shi, Q.; Akiyama, T.; Morris-Natschke, S.L.; Lee, K.-H. Recent Advances in the Investigation of Curcuminoids. Chin.
Med. 2008, 3, 11. [CrossRef] [PubMed]
293. Bao, W.; Li, K.; Rong, S.; Yao, P.; Hao, L.; Ying, C.; Zhang, X.; Nussler, A.; Liu, L. Curcumin Alleviates Ethanol-Induced
Hepatocytes Oxidative Damage Involving Heme Oxygenase-1 Induction. J. Ethnopharmacol. 2010, 128, 549–553. [CrossRef]
[PubMed]
294. Ma, T.Y.; Iwamoto, G.K.; Hoa, N.T.; Akotia, V.; Pedram, A.; Boivin, M.A.; Said, H.M. TNF-Alpha-Induced Increase in Intestinal
Epithelial Tight Junction Permeability Requires NF-Kappa B Activation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286,
G367–G376. [CrossRef]
295. Motterlini, R.; Foresti, R.; Bassi, R.; Green, C.J. Curcumin, an Antioxidant and Anti-Inflammatory Agent, Induces Heme
Oxygenase-1 and Protects Endothelial Cells against Oxidative Stress. Free Radic. Biol. Med. 2000, 28, 1303–1312. [CrossRef]
296. McNally, S.J.; Harrison, E.M.; Ross, J.A.; Garden, O.J.; Wigmore, S.J. Curcumin Induces Heme Oxygenase 1 through Generation of
Reactive Oxygen Species, P38 Activation and Phosphatase Inhibition. Int. J. Mol. Med. 2007, 19, 165–172. [CrossRef]
297. Huang, M.T.; Newmark, H.L.; Frenkel, K. Inhibitory Effects of Curcumin on Tumorigenesis in Mice. J. Cell Biochem. Suppl. 1997,
27, 26–34. [CrossRef]
298. Carmody, R.N.; Turnbaugh, P.J. Host-Microbial Interactions in the Metabolism of Therapeutic and Diet-Derived Xenobiotics. J.
Clin. Invest. 2014, 124, 4173–4181. [CrossRef] [PubMed]
299. Jiang, H. Curcumin-Attenuated Trinitrobenzene Sulphonic Acid Induces Chronic Colitis by Inhibiting Expression of
Cyclooxygenase-2. WJG 2006, 12, 3848. [CrossRef] [PubMed]
300. Scazzocchio, B.; Minghetti, L.; D’Archivio, M. Interaction between Gut Microbiota and Curcumin: A New Key of Understanding for the Health Effects of Curcumin. Nutrients 2020, 12, E2499. [CrossRef] [PubMed]
301. Peterson, C.T.; Vaughn, A.R.; Sharma, V.; Chopra, D.; Mills, P.J.; Peterson, S.N.; Sivamani, R.K. Effects of Turmeric and Curcumin
Dietary Supplementation on Human Gut Microbiota: A Double-Blind, Randomized, Placebo-Controlled Pilot Study. J. Evid. Based
Integr. Med. 2018, 23. [CrossRef]
302. Li, S.; Fu, C.; Zhao, Y.; He, J. Intervention with α -Ketoglutarate Ameliorates Colitis-Related Colorectal Carcinoma via Modulation of the Gut Microbiome. BioMed Res. Int. 2019, 2019, 1–9. [CrossRef]
303. Wang, N.; Wang, G.; Hao, J.; Ma, J.; Wang, Y.; Jiang, X.; Jiang, H. Curcumin Ameliorates Hydrogen Peroxide-Induced Epithelial
Barrier Disruption by Upregulating Heme Oxygenase-1 Expression in Human Intestinal Epithelial Cells. Dig. Dis. Sci. 2012, 57,
1792–1801. [CrossRef]
304. Leyva-Diaz, A.A.; Hernandez-Patlan, D.; Solis-Cruz, B.; Adhikari, B.; Kwon, Y.M.; Latorre, J.D.; Hernandez-Velasco, X.; FuenteMartinez, B.; Hargis, B.M.; Lopez-Arellano, R.; et al. Evaluation of Curcumin and Copper Acetate against Salmonella Typhimurium
Infection, Intestinal Permeability, and Cecal Microbiota Composition in Broiler Chickens. J. Anim. Sci. Biotechnol. 2021, 12, 23.
[CrossRef]


")











