Innovative drugs, chemicals, and enzymes within the animal production chain
The alarming number of recently reported human illnesses with bacterial infections resistant to multiple antibacterial agents has become a serious concern in recent years. This phenomenon is a core challenge for both the medical and animal health communities since the use of antibiotics has formed the cornerstone of modern medicine for treating bacterial infections. The empirical benefits of using antibiotics to address animal health issues in animal agriculture (using therapeutic doses) and increasing the overall productivity of animals (using sub-therapeutic doses) are well established. The use of antibiotics to enhance profitability margins in the animal production industry is still practiced worldwide. Although many technical and economic reasons gave rise to these practices, the continued emergence of antimicrobial resistant bacteria is furthering the need to reduce the use of medically important antibiotics. This will require improving on-farm management and biosecurity practices, and the development of effective antibiotic alternatives that will reduce the dependence on antibiotics within the animal industry in the foreseeable future. A number of approaches are being closely scrutinized and optimized to achieve this goal, including the development of promising antibiotic alternatives to control bacterial virulence through quorum-sensing disruption, the use of synthetic polymers and nanoparticles, the exploitation of recombinant enzymes/proteins (such as glucose oxidases, alkaline phosphatases and proteases), and the use of phytochemicals. This review explores the most recent approaches within this context and provides a summary of practical mitigation strategies for the extensive use of antibiotics within the animal production chain in addition to several future challenges that need to be addressed.
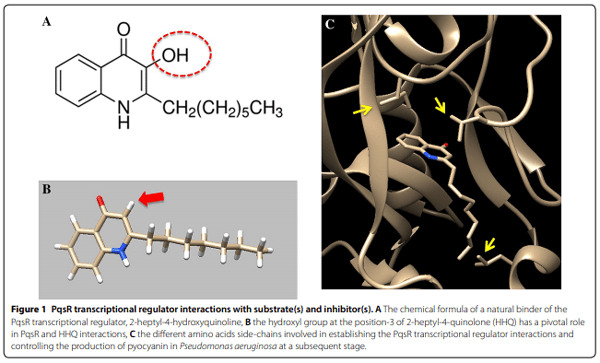
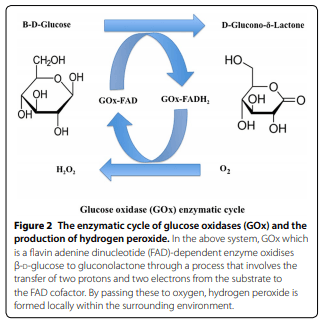
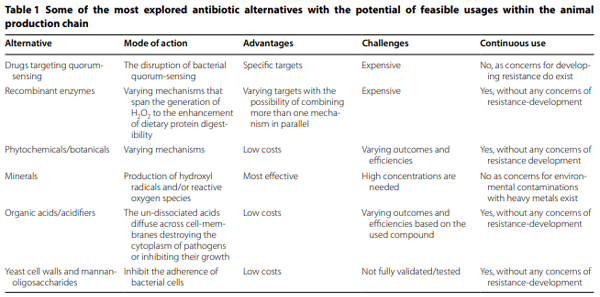
1. Founou LL, Founou RC, Essack SY (2016) Antibiotic resistance in the food chain: a developing country-perspective. Front Microbiol 7:1881
2. Johnstone J, Patel SN (2017) Provincial infectious diseases advisory committee—infection P., control, an emerging multidrug-resistant bacteria. CMAJ 189:E1115
3. Goldberg AM (2016) Farm animal welfare and human health. Curr Environ Health Rep 3:313–321
4. Martens E, Demain AL (2017) The antibiotic resistance crisis, with a focus on the United States. J Antibiot (Tokyo) 70:520–526
5. Nhung NT, Cuong NV, Thwaites G, Carrique-Mas J (2016) Antimicrobial usage and antimicrobial resistance in animal production in southeast asia: a review. Antibiotics (Basel) 5:E37
6. Rhouma M, Beaudry F, Letellier A (2016) Resistance to colistin: what is the fate for this antibiotic in pig production? Int J Antimicrob Agents 48:119–126
7. Premarathne J, Anuar AS, Thung TY, Satharasinghe DA, Jambari NN, Abdul-Mutalib NA, Huat JTY, Basri DF, Rukayadi Y, Nakaguchi Y, Nishibuchi M, Radu S (2017) Prevalence and antibiotic resistance against tetracycline in Campylobacter jejuni and E. coli in cattle and beef meat from Selangor Malaysia. Front Microbiol 8:2254
8. Shrestha A, Bajracharya AM, Subedi H, Turha RS, Kafe S, Sharma S, Neupane S, Chaudhary DK (2017) Multi-drug resistance and extended spectrum beta lactamase producing Gram negative bacteria from chicken meat in Bharatpur Metropolitan, Nepal. BMC Res Notes 10:574
9. Wang Y, Zhang A, Yang Y, Lei C, Jiang W, Liu B, Shi H, Kong L, Cheng G, Zhang X, Yang X, Wang H (2017) Emergence of Salmonella enterica serovar indiana and california isolates with concurrent resistance to cefotaxime, amikacin and ciprofoxacin from chickens in China. Int J Food Microbiol 262:23–30
10. Cameron A, McAllister TA (2016) Antimicrobial usage and resistance in beef production. J Anim Sci Biotechnol 7:68
11. Kim WH, Lillehoj HS, Gay CG (2016) Using genomics to identify novel antimicrobials. Rev Sci Tech 35:95–103
12. Allen RC, Popat R, Diggle SP, Brown SP (2014) Targeting virulence: can we make evolution-proof drugs? Nat Rev Microbiol 12:300–308
13. Majumdar S, Pal S (2017) Bacterial intelligence: imitation games, timesharing, and long-range quantum coherence. J Cell Commun Signal 11:281–284
14. Singh S, Singh SK, Chowdhury I, Singh R (2017) Understanding the mechanism of bacterial bioflms resistance to antimicrobial agents. Open Microbiol J 11:53–62
15. Saurav K, Costantino V, Venturi V, Steindler L (2017) Quorum-sensing inhibitors from the sea discovered using bacterial N-acyl-homoserine lactone-based biosensors. Mar Drugs 15:E53
16. Musthafa KS, Sianglum W, Saising J, Lethongkam S, Voravuthikunchai SP (2017) Evaluation of phytochemicals from medicinal plants of Myrtaceae family on virulence factor production by Pseudomonas aeruginosa. APMIS 125:482–490
17. Kamal AAM, Petrera L, Eberhard J, Hartmann RW (2017) Structure-functionality relationship and pharmacological profles of Pseudomonas aeruginosa alkylquinolone quorum-sensing modulators. Org Biomol Chem 15:4620–4630
18. Defoirdt T, Crab R, Wood TK, Sorgeloos P, Verstraete W, Bossier P (2006) Quorum-sensing-disrupting brominated furanones protect the gnotobiotic brine shrimp Artemia franciscana from pathogenic Vibrio harveyi, Vibrio campbellii, and Vibrio parahaemolyticus isolates. Appl Environ Microbiol 72:6419–6423
19. Pande GSJ, Scheie AA, Benneche T, Wille M, Sorgeloos P, Bossier P, Defoirdt T (2013) Quorum-sensing-disrupting compounds protect larvae of the giant freshwater prawn Macrobrachium rosenbergii from Vibrio harveyi infection. Aquaculture 406:121–124
20. Hao H, Dai M, Wang Y, Huang L, Yuan Z (2012) Key genetic elements and regulation systems in methicillin-resistant Staphylococcus aureus. Future Microbiol 7:1315–1329
21. Jeong DW, Cho H, Jones MB, Shatzkes K, Sun F, Ji Q, Liu Q, Peterson SN, He C, Bae T (2012) The auxiliary protein complex SaePQ activates the phosphatase activity of sensor kinase SaeS in the SaeRS two-component system of Staphylococcus aureus. Mol Microbiol 86:331–348
22. Baldry M, Nielsen A, Bojer MS, Zhao Y, Friberg C, Ifrah D, Glasser Heede N, Larsen TO, Frokiaer H, Frees D, Zhang L, Dai H, Ingmer H (2016) Norlichexanthone reduces virulence gene expression and bioflm formation in Staphylococcus aureus. PLoS One 11:e0168305
23. Yang C, Chowdhury MA, Huo Y, Gong J (2015) Hytogenic compounds as alternatives to in-feed antibiotics: potentials and challenges in application. Pathogens 4:137–156
24. Cheng G, Hao H, Xie S, Wang X, Dai M, Huang L, Yuan Z (2014) Antibiotic alternatives: the substitution of antibiotics in animal husbandry? Front Microbiol 5:217
25. Ricke SC (2003) Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult Sci 82:632–639
26. Jones FT, Ricke SC (2003) Observations on the history of the development of antimicrobials and their use in poultry feeds. Poult Sci 82:613–617
27. Emami NK, Daneshmand A, Naeini SZ, Graystone EN, Broom LJ (2017) Efects of commercial organic acid blends on male broilers challenged with E. coli K88: performance, microbiology, intestinal morphology, and immune response. Poult Sci 96:3254–3263
28. Hassan HMA, Mohamed MA, Youssef AW, Hassan ER (2010) Efect of using organic acids to substitute antibiotic growth promoters on performance and intestinal microfora of broilers. Asian-Australas J Anim Sci 23:1348–1353
29. Van Immerseel F, De Buck J, Pasmans F, Velge P, Bottreau E, Fievez V, Haesebrouck F, Ducatelle R (2003) Invasion of Salmonella enteritidis in avian intestinal epithelial cells in vitro is infuenced by short-chain fatty acids. Int J Food Microbiol 85:237–248
30. Gantois I, Ducatelle R, Pasmans F, Haesebrouck F, Hautefort I, Thompson A, Hinton JC, Van Immerseel F (2006) Butyrate specifcally down-regulates salmonella pathogenicity island 1 gene expression. Appl Environ Microbiol 72:946–949
31. Van Immerseel F, Russell JB, Flythe MD, Gantois I, Timbermont L, Pasmans F, Haesebrouck F, Ducatelle R (2006) The use of organic acids to combat Salmonella in poultry: a mechanistic explanation of the efcacy. Avian Pathol 35:182–188
32. Van Immerseel F, Fievez V, de Buck J, Pasmans F, Martel A, Haesebrouck F, Ducatelle R (2004) Microencapsulated short-chain fatty acids in feed modify colonization and invasion early after infection with Salmonella enteritidis in young chickens. Poult Sci 83:69–74
33. Liu Y, Yang X, Xin H, Chen S, Yang C, Duan Y, Yang X (2017) Efects of a protected inclusion of organic acids and essential oils as antibiotic growth promoter alternative on growth performance, intestinal morphology and gut microfora in broilers. Anim Sci J 88:1414–1424
34. Upadhaya SD, Lee KY, Kim IH (2014) Protected organic acid blends as an alternative to antibiotics in fnishing pigs. Asian-Australas J Anim Sci 27:1600–1607
35. Lund P, Tramonti A, De Biase D (2014) Coping with low pH: molecular strategies in neutralophilic bacteria. FEMS Microbiol Rev 38:1091–1125
36. Rivera-Chavez F, Zhang LF, Faber F, Lopez CA, Byndloss MX, Olsan EE, Xu G, Velazquez EM, Lebrilla CB, Winter SE, Baumler AJ (2016) Depletion of butyrate-producing Clostridia from the gut microbiota drives an aerobic luminal expansion of Salmonella. Cell Host Microbe 19:443–454
37. Kaczmarek SA, Barri A, Hejdysz M, Rutkowski A (2016) Efect of diferent doses of coated butyric acid on growth performance and energy utilization in broilers. Poult Sci 95:851–859
38. Chen JL, Zheng P, Zhang C, Yu B, He J, Yu J, Luo JQ, Mao XB, Huang ZQ, Chen DW (2016) Benzoic acid benefcially afects growth performance of weaned pigs which was associated with changes in gut bacterial populations, morphology indices and growth factor gene expression. J Anim Physiol Anim Nutr (Berl) 101:1137–1146
39. Messens W, Goris J, Dierick N, Herman L, Heyndrickx M (2010) Inhibition of Salmonella typhimurium by medium-chain fatty acids in an in vitro simulation of the porcine cecum. Vet Microbiol 141:73–80
40. Ferrara F, Tedin L, Pieper R, Meyer W, Zentek J (2017) Infuence of medium-chain fatty acids and short-chain organic acids on jejunal morphology and intra-epithelial immune cells in weaned piglets. J Anim Physiol Anim Nutr (Berl) 101:531–540
41. Van Immerseel F, De Buck J, Boyen F, Bohez L, Pasmans F, Volf J, Sevcik M, Rychlik I, Haesebrouck F, Ducatelle R (2004) Medium-chain fatty acids decrease colonization and invasion through hilA suppression shortly after infection of chickens with Salmonella enterica serovar Enteritidis. Appl Environ Microbiol 70:3582–3587
42. Boyen F, Haesebrouck F, Vanparys A, Volf J, Mahu M, Van Immerseel F, Rychlik I, Dewulf J, Ducatelle R, Pasmans F (2008) Coated fatty acids alter virulence properties of Salmonella Typhimurium and decrease intestinal colonization of pigs. Vet Microbiol 132:319–327
43. Zentek J, Buchheit-Renko S, Ferrara F, Vahjen W, Van Kessel AG, Pieper R (2011) Nutritional and physiological role of medium-chain triglycerides and medium-chain fatty acids in piglets. Anim Health Res Rev 12:83–93
44. Omonijo FA, Ni L, Gong J, Wang Q, Lahaye L, Yang C (2018) Essential oils as alternatives to antibiotics in swine production. Anim Nutr, in press https://doi.org/10.1016/j.aninu.2017.09.001
45. Mehdi Y, Dufrasne I (2016) Selenium in cattle: a review. Molecules 21:545
46. Heo JM, Opapeju FO, Pluske JR, Kim JC, Hampson DJ, Nyachoti CM (2013) Gastrointestinal health and function in weaned pigs: a review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J Anim Physiol Anim Nutr (Berl) 97:207–237
47. Debski B (2016) Supplementation of pigs diet with zinc and copper as alternative to conventional antimicrobials. Pol J Vet Sci 19:917–924
48. Prasai TP, Walsh KB, Bhattarai SP, Midmore DJ, Van TT, Moore RJ, Stanley D (2017) Zeolite food supplementation reduces abundance of enterobacteria. Microbiol Res 195:24–30
49. Jahanbakhsh S, Letellier A, Fairbrother JM (2016) Circulating of CMY-2 beta-lactamase gene in weaned pigs and their environment in a commercial farm and the efect of feed supplementation with a clay mineral. J Appl Microbiol 121:136–148
50. Lee JA, Jung BG, Kim TH, Kim YM, Koh HB, Lee BJ (2015) Improvement of bacterial clearance and relief of clinical signs of Salmonella enterica serovar Typhimurium infection in pigs through upregulation of Th 1-specifc responses by administration of a combination of two silicate minerals, biotite and bentonite. J Vet Med Sci 77:1087–1094
51. Wu DW, Wang LC, Wen C, Hooge DM, Liang C, Zhou YM (2013) Efects of replacing a dietary antibacterial agent (zinc bacitracin) with copper salts in Cherry Valley Pekin meat ducks. Br Poult Sci 54:112–119
52. Smith JW 2nd, Tokach MD, Goodband RD, Nelssen JL, Richert BT (1997) Efects of the interrelationship between zinc oxide and copper sulfate on growth performance of early-weaned pigs. J Anim Sci 75:1861–1866
53. Sales J (2013) Efects of pharmacological concentrations of dietary zinc oxide on growth of post-weaning pigs: a meta-analysis. Biol Trace Elem Res 152:343–349
54. Starke IC, Pieper R, Neumann K, Zentek J, Vahjen W (2014) The impact of high dietary zinc oxide on the development of the intestinal microbiota in weaned piglets. FEMS Microbiol Ecol 87:416–427
55. Wang C, Zhang L, Su W, Ying Z, He J, Zhang L, Zhong X, Wang T (2017) Zinc oxide nanoparticles as a substitute for zinc oxide or colistin sulfate: efects on growth, serum enzymes, zinc deposition, intestinal morphology and epithelial barrier in weaned piglets. PLoS One 12:e0181136
56. Mehdi Y, Hornick JL, Istasse L, Dufrasne I (2013) Selenium in the environment, metabolism and involvement in body functions. Molecules 18:3292–3311
57. Walk CL, Wilcock P, Magowan E (2015) Evaluation of the efects of pharmacological zinc oxide and phosphorus source on weaned piglet growth performance, plasma minerals and mineral digestibility. Animal 9:1145–1152
58. Vahjen W, Pietruszynska D, Starke IC, Zentek J (2015) High dietary zinc supplementation increases the occurrence of tetracycline and sulfonamide resistance genes in the intestine of weaned pigs. Gut Pathog 7:23
59. Di Cesare A, Eckert E, Corno G (2016) Co-selection of antibiotic and heavy metal resistance in freshwater bacteria. J Limnol 75:59–66
60. Wales AD, Davies RH (2015) Co-selection of resistance to antibiotics, biocides and heavy metals, and its relevance to foodborne pathogens. Antibiotics (Basel) 4:567–604
61. Yazdankhah S, Rudi K, Bernhoft A (2014) Zinc and copper in animal feed-development of resistance and co-resistance to antimicrobial agents in bacteria of animal origin. Microb Ecol Health Dis 25:25862
62. Perri A, Friendship R, Harding J, O’Sullivan T (2016) An investigation of iron defciency and anemia in piglets and the efect of iron status at weaning on post-weaning performance. J Swine Health Prod 24:10–20
63. Wang MM, Wang YC, Wang XN, Liu Y, Zhang H, Zhang JW, Huang Q, Chen SP, Hei TK, Wu LJ, Xu A (2015) Mutagenicity of ZnO nanoparticles in mammalian cells: role of physicochemical transformations under the aging process. Nanotoxicology 9:972–982
64. Westendarp H (2005) Essential oils for the nutrition of poultry, swine and ruminants. Dtsch Tierarztl Wochenschr 112:375–380 (in German)
65. Helander IM, Alakomi HL, Latva-Kala K, Mattila-Sandholm T, Pol I, Smid EJ, Gorris LG, von Wright A (1998) Characterization of the action of selected essential oil components on Gram-negative bacteria. J Agric Food Chem 46:3590–3595
66. Gutiérrez-Fernández J, García-Armesto MR, Álvarez-Alonso R, del Valle P, de Arriaga D, Rúa J (2011) Antimicrobial activity of binary combinations of natural and synthetic phenolic antioxidants against Enterococcus faecalis. J Dairy Sci 96:4912–4920
67. Wei HK, Xue HX, Zhou Z, Peng J (2017) A carvacrol–thymol blend decreased intestinal oxidative stress and infuenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 11:193–201
68. Li P, Piao X, Ru Y, Han X, Xue L, Zhang H (2012) Efects of adding essential oil to the diet of weaned pigs on performance, nutrient utilization, immune response and intestinal health. Asian-Australas J Anim Sci 25:1617–1626
69. Tiihonen K, Kettunen H, Bento MHL, Saarinen M, Lahtinen S, Ouwehand AC, Schulze H, Rautonen N (2010) The efect of feeding essential oils on broiler performance and gut microbiota. Br Poult Sci 51:381–392
70. Hussein MMAH, Hassan WH, Moussa IM (2013) Potential use of allicin (garlic, Allium sativum Linn, essential oil) against fsh pathogenic bacteria and its safety for monosex Nile tilapia (Oreochromis niloticus). J Food Agric Environ 11:696–699
71. Huang RH, Qiu XS, Shi FX, Hughes CL, Lu ZF, Zhu WY (2011) Efects of dietary allicin on health and growth performance of weanling piglets and reduction in attractiveness of faeces to fies. Animal 5:304–311
72. Roselli M, Britti MS, Le Huërou-Luron I, Marfaing H, Zhu WY, Mengheri E (2007) Efect of diferent plant extracts and natural substances (PENS) against membrane damage induced by enterotoxigenic Escherichia coli K88 in pig intestinal cells. Toxicol In Vitro 21:224–229
73. Chowdhury AK, Ahsan M, Islam SN, Ahmed ZU (1991) Efcacy of aqueous extract of garlic & allicin in experimental shigellosis in rabbits. Indian J Med Res 93:33–36
74. Robyn J, Rasschaert G, Hermans D, Pasmans F, Heyndrickx M (2013) Is allicin able to reduce Campylobacter jejuni colonization in broilers when added to drinking water? Poult Sci 92:1408–1418
75. Namkung H, Li J, Gong M, Yu H, Cottrill M, De Lange C (2004) Impact of feeding blends of organic acids and herbal extracts on growth performance, gut microbiota and digestive function in newly weaned pigs. Can J Anim Sci 84:697–704
76. Manzanilla E, Nofrarias M, Anguita M, Castillo M, Perez J, Martin-Orue S, Kamel C, Gasa J (2006) Efects of butyrate, avilamycin, and a plant extract combination on the intestinal equilibrium of early-weaned pigs. J Anim Sci 84:2743–2751
77. Nofrarias M, Manzanilla E, Pujols J, Gibert X, Majo N, Segalés J, Gasa J (2006) Efects of spray-dried porcine plasma and plant extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J Anim Sci 84:2735–2742
78. Huang Y, Yoo J, Kim H, Wang Y, Chen Y, Cho J, Kim I (2010) Efects of dietary supplementation with blended essential oils on growth performance, nutrient digestibility, blood profles and fecal characteristics in weanling pigs. Asian-Australas J Anim Sci 23:607
79. Li SY, Ru YJ, Liu M, Xu B, Péron A, Shi XG (2012) The efect of essential oils on performance, immunity and gut microbial population in weaner pigs. Livest Sci 145:119–123
80. Molnar C, Bilkei G (2005) The infuence of an oregano feed additive on production parameters and mortality of weaned piglets. Tierärztliche Praxis Großtiere 33:42–47
81. Zou Y, Wang J, Peng J, Wei H (2016) Oregano essential oil Induces SOD1 and GSH expression through Nrf2 activation and alleviates hydrogen peroxide-induced oxidative damage in IPEC-J2 cells. Oxid Med Cell Longev 2016:5987183
82. Wondrak GT, Villeneuve NF, Lamore SD, Bause AS, Jiang T, Zhang DD (2010) The cinnamon-derived dietary factor cinnamic aldehyde activates the Nrf2-dependent antioxidant response in human epithelial colon cells. Molecules 15:3338–3355
83. Gessner DK, Fiesel A, Most E, Dinges J, Wen G, Ringseis R, Eder K (2013) Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κB and Nrf2 in the duodenal mucosa of pigs. Acta Vet Scand 55:18
84. Wei H-K, Chen G, Wang R-J, Peng J (2015) Oregano essential oil decreased susceptibility to oxidative stress-induced dysfunction of intestinal epithelial barrier in rats. J Funct Foods 18:1191–1199
85. Zhao CY, Tan SX, Xiao XY, Qiu XS, Pan JQ, Tang ZX (2014) Efects of dietary zinc oxide nanoparticles on growth performance and antioxidative status in broilers. Biol Trace Elem Res 160:361–367
86. Li MZ, Huang JT, Tsai YH, Mao SY, Fu CM, Lien TF (2016) Nanosize of zinc oxide and the efects on zinc digestibility, growth performances, immune response and serum parameters of weanling piglets. Anim Sci J 87:1379–1385
87. Milani NC, Sbardella M, Ikeda NY, Arno A, Mascarenhas BC, Miyada VS (2017) Dietary zinc oxide nanoparticles as growth promoter for weanling pigs. Anim Feed Sci Technol 227:13–23
88. Wessels S, Ingmer H (2013) Modes of action of three disinfectant active substances: a review. Regul Toxicol Pharmacol 67:456–467
89. Xue X, Chen X, Mao X, Hou Z, Zhou Y, Bai H, Meng J, Da F, Sang G, Wang Y, Luo X (2013) Amino-terminated generation 2 poly(amidoamine) dendrimer as a potential broad-spectrum, nonresistance-inducing antibacterial agent. AAPS J 15:132–142
90. Lienkamp K, Madkour AE, Musante A, Nelson CF, Nusslein K, Tew GN (2008) Antimicrobial polymers prepared by ROMP with unprecedented selectivity: a molecular construction kit approach. J Am Chem Soc 130:9836–9843
91. Sadiq S, Imran M, Habib H, Shabbir S, Ihsan A, Zafar Y, Hafeez FY (2016) Potential of monolaurin based food-grade nano-micelles loaded with nisin Z for synergistic antimicrobial action against Staphylococcus Aureus. LWT–Food Sci Technol 71:227–233
92. Silvaa MD, Ning C, Ghanbar S, Zhanel G, Logsetty S, Liu S, Kumar A (2015) Evidence that a novel quaternary compound and its organic N-chloramine derivative do not select for resistant mutants of Pseudomonas aeruginosa. J Hosp Infect 91:53–58
93. Ghanbar S, Fumakia M, Ho EA, Liu S (2018) A new strategy for battling bacterial resistance: turning potent, non-selectively and potentially non-resistance-inducing biocides into selective ones. Nanomedicine 14:471–481
94. Gifre L, Aris A, Bach A, Garcia-Fruitos E (2017) Trends in recombinant protein use in animal production. Microb Cell Fact 16:40
95. Cooper RA (2013) Inhibition of bioflms by glucose oxidase, lactoperoxidase and guaiacol: the active antibacterial component in an enzyme alginogel. Int Wound J 10:630–637
96. Muizzuddin N, Schnittger S, Maher W, Maes DH, Mammone T (2013) Enzymatically generated hydrogen peroxide reduces the number of acne lesions in acne vulgaris. J Cosmet Sci 64:1–8
97. Wyns H, Plessers E, De Backer P, Meyer E, Croubels S (2015) In vivo porcine lipopolysaccharide infammation models to study immunomodulation of drugs. Vet Immunol Immunopathol 166:58–69
98. Drago-Serrano ME, de la Garza-Amaya M, Luna JS, Campos-Rodriguez R (2012) Lactoferrin-lipopolysaccharide (LPS) binding as key to antibacterial and antiendotoxic efects. Int Immunopharmacol 12:1–9
99. Vaishnava S, Hooper LV (2007) Alkaline phosphatase: keeping the peace at the gut epithelial surface. Cell Host Microbe 2:365–367
100. Buchet R, Millan JL, Magne D (2013) Multisystemic functions of alkaline phosphatases. Methods Mol Biol 1053:27–51
101. Melo AD, Silveira H, Luciano FB, Andrade C, Costa LB, Rostagno MH (2016) Intestinal alkaline phosphatase: potential roles in promoting gut health in weanling piglets and its modulation by feed additives-a review. Asian-Australas J Anim Sci 29:16–22
102. Melo AD, Silveira H, Bortoluzzi C, Lara LJ, Garbossa CA, Preis G, Costa LB, Rostagno MH (2016) Intestinal alkaline phosphatase and sodium butyrate may be benefcial in attenuating LPS-induced intestinal infammation. Genet Mol Res, in press https://doi.org/10.4238/gmr15 048875
103. Jiang WD, Xu J, Zhou XQ, Wu P, Liu Y, Jiang J, Kuang SY, Tang L, Tang WN, Zhang YA, Feng L (2017) Dietary protein levels regulated antibacterial activity, infammatory response and structural integrity in the head kidney, spleen and skin of grass carp (Ctenopharyngodon idella) after challenged with Aeromonas hydrophila. Fish Shellfsh Immunol 68:154–172
104. Xu J, Feng L, Jiang WD, Wu P, Liu Y, Jiang J, Kuang SY, Tang L, Tang WN, Zhang YA, Zhou XQ (2016) Efects of dietary protein levels on the disease resistance, immune function and physical barrier function in the gill of grass carp (Ctenopharyngodon idella) after challenged with Flavobacterium columnare. Fish Shellfsh Immunol 57:1–16
105. Zuo J, Ling B, Long L, Li T, Lahaye L, Yang C, Feng D (2015) Efect of dietary supplementation with protease on growth performance, nutrient digestibility, intestinal morphology, digestive enzymes and gene expression of weaned piglets. Anim Nutri 1:276–282
106. Guggenbuhl P, Wache Y, Wilson JW (2012) Efects of dietary supplementation with a protease on the apparent ileal digestibility of the weaned piglet. J Anim Sci 90(Suppl 4):152–154
107. Mahmood T, Mirza MA, Nawaz H, Shahid M, Athar M, Hussain M (2017) Efect of supplementing exogenous protease in low protein poultry by-product meal based diets on growth performance and nutrient digestibility in broilers. Anim Feed Sci Tech 228:23–31
108. Lahaye L, Philippe F, Bodin J (2015) Protease use in grower-fnisher pigs allows gain in feed costs for similar performance. In: 13th digestive physiology in pigs symposium, vol 104. p 1.38
109. Tactacan GB, Cho SY, Cho JH, Kim IH (2016) Performance responses, nutrient digestibility, blood characteristics, and measures of gastrointestinal health in weanling pigs fed protease enzyme. Asian-Australas J Anim Sci 29:998–1003
110. Yang Y, Iji PA, Kocher A, Thomson E, Mikkelsen LL, Choct M (2008) Efects of mannanoligosaccharide in broiler chicken diets on growth performance, energy utilisation, nutrient digestibility and intestinal microfora. Br Poult Sci 49:186–194
111. Van Nevel CJ, Decuypere JA, Dierick NA, Molly K (2005) Incorporation of galactomannans in the diet of newly weaned piglets: efect on bacteriological and some morphological characteristics of the small intestine. Arch Anim Nutr 59:123–138
112. Li WF, Feng J, Xu ZR, Yang CM (2004) Efects of non-starch polysaccharides enzymes on pancreatic and small intestinal digestive enzyme activities in piglet fed diets containing high amounts of barley. World J Gastroenterol 10:856–859
113. Xiao R, Power RF, Mallonee D, Routt K, Spangler L, Pescatore AJ, Cantor AH, Ao T, Pierce JL, Dawson KA (2012) Efects of yeast cell wall-derived mannan-oligosaccharides on jejunal gene expression in young broiler chickens. Poult Sci 91:1660–1669
114. Ramirez-Hernandez A, Rupnow J, Hutkins RW (2015) Adherence reduction of Campylobacter jejuni and Campylobacter coli strains to HEp-2 cells by mannan oligosaccharides and a high-molecular-weight component of cranberry extract. J Food Prot 78:1496–1505
115. Brufau MT, Martin-Venegas R, Guerrero-Zamora AM, Perez-Vendrell AM, Vila B, Brufau J, Ferrer R (2015) Dietary beta-galactomannans have benefcial efects on the intestinal morphology of chickens challenged with Salmonella enterica serovar Enteritidis. J Anim Sci 93:238–246
116. White LA, Newman MC, Cromwell GL, Lindemann MD (2002) Brewers dried yeast as a source of mannan oligosaccharides for weanling pigs. J Anim Sci 80:2619–2628
117. Bucekova M, Valachova I, Kohutova L, Prochazka E, Klaudiny J, Majtan J (2014) Honeybee glucose oxidase-its expression in honeybee workers and comparative analyses of its content and H2O2-mediated antibacterial activity in natural honeys. Naturwissenschaften 101:661–670
118. Suleman L (2016) Extracellular bacterial proteases in chronic wounds: a potential therapeutic target? Adv Wound Care (New Rochelle) 5:455–463
119. Kim MK, Zhao A, Wang A, Brown ZZ, Muir TW, Stone HA, Bassler BL (2017) Surface-attached molecules control Staphylococcus aureus quorum-sensing and bioflm development. Nat Microbiol 2:17080







United States





