1. Introduction
Primarily for the provision of energy density, grain sorghum is quite frequently included in rations for pigs, poultry and feed-lot cattle. However, in a recent meeting held by the Australian Feed Grain Partnership in December 2012, nutritionists from the three industries expressed concern about the efficiency of energy utilisation in animals offered sorghum-based diets. Essentially, the energy provided by sorghum is derived from starch and, indeed, Black, et al. [1] had already drawn attention to this shortcoming.
In order to address the conundrum that is quantifying the true feed quality traits of grain sorghum the concept of the “Bermuda triangle” was developed on the premise that three factors inherent in sorghum are responsible for sub-standard starch and energy utilisation. The three factors nominated, or the apices of the Bermuda triangle, were (i) kafirin, the dominant protein fraction in grain sorghum; (ii) phenolic compounds, potentially including condensed tannin and (iii) phytate or myo-inositol hexaphosphate (IP6). The inference is that sorghum starch is not intrinsically problematic but these three starch-extrinsic factors combine to compromise energy/starch utilisation. It has been proposed that the kafirin proportion of total sorghum protein is escalating in Australia which, if valid, would compromise the nutrient quality of sorghum protein [2]. However, this was not then seen as a major issue from the stand-point of starch utilisation. Almost certainly, contemporary Australian sorghum crops no longer contain condensed tannin; nevertheless, the likelihood is that “non-tannin” phenolic compounds may negatively influence energy utilisation. Phytate may limit starch utilisation and/or glucose absorption thereby negatively influencing energy utilisation. Thus the prime objective of this review is to consider the three apices of the sorghum Bermuda triangle in order to address the conundrum that is grain sorghum as a feedstuff for chicken-meat production.
2. Background
In Australia, sorghum (Sorghum bicolor L.) is a common energy source in diets for pigs, poultry and beef cattle as it can be grown under drier conditions than those suitable for wheat and maize [3]. Grain sorghum may be exported but very little is used for human consumption. In Australia, sorghum is the second feed grain to wheat as the basis of diets for broiler chickens and there has been a considerable research focus on sorghum initiated by the reports of Perez-Maldonado and Rodrigues [4] and Bryden, et al. [5].
Almost certainly, sorghum still suffers from the stigma of condensed tannin (CT), a potent polyphenolic anti-nutritive factor associated with “bird-proof” sorghums. In a recent local survey of pig and poultry nutritionists, the majority believed that sorghum contained some CT and a minority thought CT was present in sufficient concentrations to compromise animal performance. However, the Poultry Research Foundation has subjected more than 60 sorghum samples to the quantal Clorox bleach test [6] and none possessed a pigmented testa; this indicates that they do not contain CT. This has been confirmed with vanillin assays to quantify CT concentrations and local sorghums are now “tannin-free” Type I varieties. Thus our contention, contrary to the report by Perez-Maldonado and Rodrigues [4], is that contemporary Australian sorghum crops do not contain condensed tannin.
The perception is that sorghum is routinely inferior to wheat or even “sub-optimal” but our impression is that this is not necessarily valid. Nevertheless, it is conceded that broilers offered wheat-based diets are advantaged by better pellet quality given the lower starch gelatinisation temperature of wheat in comparison to sorghum. Moreover, broiler chickens offered wheat-based diets are far more likely to respond to feed enzymes, especially non starch polysaccharides (NSP)-degrading enzymes, than their counterparts on sorghum.
Nevertheless, Liu, et al. [7] reported that sorghum-based diets supported similar weight gain and feed conversion efficiency as maize-based diets, and both outperformed wheat-based diets. Ao and Choct [8] found that sorghum-based diets generated superior weight gains than broiler diets based on wheat and barley to 21 days post-hatch. In another comparison, Selle, et al. [9] showed that broilers offered non-supplemented, sorghum-based diets numerically outperformed wheat in growth performance but this was reversed by the addition of an exogenous xylanase; xylanase in sorghum-based diets did not statistically influence apparent metabolisable energy (AME); whereas, in wheat-based diets xylanase significantly improved AME by 1.21 MJ (14.76 versus 13.55 MJ/kg). The failure of a “pure” xylanase feed enzyme to elicit responses in sorghum-based diets is not surprising because sorghum is a “non-viscous” grain containing low levels of NSP that have the capacity to increase digesta viscosity in the gut and impede digestion and absorption of nutrients.
The problematic growth performance of broilers offered sorghum-based diets has been extensively reviewed [2,3,5,10,11]. Briefly, it is recognised that kafirin, the dominant fraction in sorghum protein, is poorly digested due to its hydrophobicity [12,13]. Protein–phytate interactions stem from electrostatic interactions between phytate and basic amino acid residues of arginine, histidine and lysine at pH levels below protein isoelectric points [14]. Kafirin has an unfavourable amino acid profile as it contains a paucity of basic amino acids, especially lysine [15] and, therefore, it is less likely to form protein-phytate complexes. It is well accepted that the potential phytate- protein interaction may reduce the solubility of protein and therefore, impede its digestion [16]. This may partially explain the lack of “extra-phosphoric” effects following the inclusion of phytase in sorghum-based broiler diets.
It is well established that condensed tannin, when present in sorghum, has the capacity to depress protein digestion and compromise bird performance [13]. However, condensed tannin, a polyphenolic compound and potent anti-nutritive factor is almost certainly not present in contemporary Australian sorghum crops. However, the possibility remains that “non-tannin” phenolic compounds in sorghum negatively influence digestion of protein and starch and compromise growth performance in broiler chickens [17]. Liu, et al. [18] reported white sorghum-based diets supported better amino acid digestibilities than red sorghum and it may be that soluble conjugated and insoluble bound phenolic acids hold the key to the apparent superiority of white sorghum. Ferulic acid is the most abundant phenolic acid in sorghum but it certainly is not unique to sorghum.
Selle, et al. [19] reported sorghum contained an average of 2.92 g/kg total phosphorus (P) and the proportion of phytate-bound P of total P was higher in sorghum (82.7%) than barley (67.3%) and wheat (74.9%). Fifteen sorghum varieties contained an average of 2.41 g/kg phytate-P or 8.55 g/kg IP6 phytate. The primary purpose for including phytase in broiler diets is to liberate phytate-bound P [20]. This “phosphoric effect” of phytase in sorghum-based diet is in evidence as Truong, et al. [21] reported that phytase significantly increased apparent disappearance rates of phytate in the proximal jejunum, distal jejunum, proximal ileum and distal ileum in boilers. Also, Liu, et al. [7] showed that phytase significantly increased toe ash from 11.68% versus 12.29% in broiler chickens offered sorghum-based diets. However, the “extra-phosphoric” responses to phytase sorghum-based diets often appear to be muted. The formation of protein-phytase complexes in the gut at pH less than the isoelectric point of protein well may be fundamental to the anti-nutritive properties of phytate and their attenuation generates the “extra-phosphoric” protein and energy responses to phytase [16]. However, from first principles, phytate may not readily complex kafirin because it contains a paucity of basic amino acids [11]. Moreover, this may be compounded by the fact that kafirin is contained in discrete protein bodies in sorghum endosperm with outer hydrophobic layers of cross-linked β- and γ-kafirin. Thus, these chemical and physical factors may limit interactions or complex formation between phytate and kafirin and the pursuant “extra-phosphoric” phytase responses.
Liu and Selle [22] proposed the provision of either rapidly digestible protein or slowly digestible starch will enhance bird performance and that the protein digestion rate may be more important than starch in this reciprocal or bilateral relationship. Compared to other grains, sorghum has relatively slower starch digestion rate [23]. Strategies that enhance protein digestion in sorghum may result in a better balance between the post-enteral availability of glucose and amino acid in chickens and improve growth performance. Therefore, the utilisation of exogenous enzymes and sulfite reducing agents is discussed in this review. Because of the inconsistent growth performance and nutrient utilisation in sorghum-based broiler diets, it is imperative to develop methods to determine sorghum grain quality. Starch pasting profiles determined by rapid visco-analyser (RVA), Promatest protein solubilities and Symes particle size index (PSI) textures have been assessed in a series of studies and their relevance is discussed in this review.
3. Starch
Starch is the most abundant component in sorghum with amylopectin the dominant single polymer at around 75% of total starch, with amylose being around 25%. On average, sorghum contains 70% starch; therefore, it is considered to be the principal grain component used for energy. The utilisation of sorghum starch by broiler chickens as an energy source does appear to be sub-standard in relation to maize. As reviewed by Truong, et al. [24] the distal ileal digestibility of maize starch is superior to that of sorghum. These researchers reported that, across 11 studies, maize starch had an average ileal digestibility coefficient of 0.950 (range: 0.873 to 0.993). Alternatively, across seven studies, sorghum starch had an average ileal starch digestibility coefficient of 0.883 (range: 0.846 to 0.921). On the basis of this comparison, sorghum starch digestibility was inferior to maize by 7.05%. However, this inferiority may not be due to the digestibility of starch per se but to extrinsic factors exerting negative impacts. Nevertheless, starch digestibility per se is influenced by amylose to amylopectin ratios, the two constituents of starch.
Sang, et al. [25] reported that in vitro starch digestion in waxy sorghum (0% amylose) was higher than heterowaxy sorghum (14% amylose) and normal sorghum (23.7% amylose). However, kafirin protein bodies were observed to be more numerous and closely associated with starch granules in the endosperm of non-waxy sorghums [26]. Moreover, sorghum grains are reported to contain about 6.5% resistant starch, and this may be increased following cooling of cooked sorghum due to starch recrystallisation [27].
The amylose:amylopectin ratio in grains may contribute to variations in starch digestion rates. However, inconsistent findings for the impact of amylose on broiler performance and energy utilisation have been reported. Svihus, et al. [28] reported that high amylose contents in cereal grains depress starch digestibility. Ertl and Dale [29] found significant differences in metabolisable energy in different hybrids of maize; however, there was no significant difference of metabolisable energy between normal and waxy maize from the same hybrid. Pirgozliev, et al. [30] investigated the performance of broilers offered wheat-based diets supplemented with three levels of dietary maize starch and three amylose:amylopectin ratios. Theses researchers found no significant differences in feed intake, weight gain and feed conversion ratio between dietary treatments. In contrast, Collins, et al. [31] reported that waxy-maize-based diets had significantly higher weight gain (2136 g/bird versus 1910 g/bird) and feed intake (3882 g/bird versus 3366 g/bird) from 18–42 days post-hatch and this could be due to the pellet durability index in diets based on waxy maize which were significantly higher than normal maize in grower (75.4% versus 54.6%) and finisher diets (77.9% versus 61.3%). Therefore, it remains unclear as to whether or not high amylose content in cereals depresses broiler performance. Based on the discussion above, amylose may retard starch digestion and synchronise starch and protein digestion but, alternatively, high amylose contents may negatively influence pellet quality and depress broiler performance by reducing feed intake. It seems that waxy sorghum may have more rapid protein digestion coupled with more complete starch digestion; therefore, waxy sorghum may perform better in diets for broiler chickens. Unlike maize and wheat, the application of waxy-sorghum in broiler diets is highly limited and it is difficult to clarify its possible impact on growth performance in poultry.
Further to our understanding of starch, amylose and amylopectin until recently has been somewhat limited to measuring the amount of these. A recent reported identified a genetic change in one of the starch synthesis enzymes (namely pullulanase) which resulted in major changes in amylopectin structural properties including digestibility when measure in vitro [32]. This suggests variation in starch structure as well as using specific starch types, such as waxy, may contribute to improved digestibility.
4. Apices of the Sorghum Bermuda Triangle
4.1. Kafirin
Kafirin is the dominant fraction of sorghum protein and Virupaksha and Sastry [33] reported that kafirin constituted 54.1%, glutelin 33.4%, globulin 7.0% and albumin 5.6% of endosperm protein in sorghum. Kafirin is located in discrete protein bodies which are embedded in the glutelin protein matrix of sorghum endosperm where both kafirin and glutelin are intimately associated with starch granules. Subsequently, Taylor, et al. [34] reported average concentrations of 54.0 g/kg kafirin and 27.1 g/kg glutelin in 41 grain sorghum samples with an average crude protein content of 111 g/kg. Thus kafirin and glutelin comprised 48.0% and 27.7% of sorghum protein, respectively. However, kafirin, as a proportion of protein, was positively correlated (r = 0.469; p < 0.005) with sorghum protein concentrations; whereas, the proportion of glutelin was negatively correlated (r = −0.401; p < 0.01). This may suggest that screening sorghum quality by its protein concentration may result in sorghum grains with high kafirin concentrations.
Selle, et al. [35] proposed kafirin concentrations in Australian sorghums have increased in recent decades probably as an unintended consequence of breeding for increased yield and the use of high input nitrogen fertilizers. While this proposal is based on circumstantial evidence, the likely implications are important. In a quest to enhance grain weathering resistance, Henzell [36] stated breeding programs commonly target sorghums with a relatively dense or corneous endosperm. As such, selecting sorghums with hard endosperms will result in higher kafirin contents. Importantly, the textures of Australian sorghums are relatively high by international standards. Our group has determined the Symes PSI texture [37] of 38 sorghum varieties; the mean PSI value was 9.84 ± 1.603 which is in the more rigid end of the “very hard” scale and only two varieties or 5.3% of samples were “softer” in this scale. These findings are in contrast to an extensive global survey recorded by de Alencar Figueiredo, et al. [38]. In this survey the mean PSI texture of 117 sorghum samples was 12.7 but 44% of samples had softer textures than “very hard” as opposed to 5.3% in the local sorghum varieties. Moreover, the likelihood is that “soft” sorghums are superior in performance and nutrient utilisation to “hard” sorghums as the basis for broiler diets [39].
It is instructive to compare the amino acid profiles of local sorghum varieties from related reports compiled by Ravindran, et al. [40] and subsequently by Bryden, et al. [5], where the same analytical methods were adopted. The mean protein content of sorghums in the two studies was very similar (98.7 g/kg versus 103.6 g/kg). However, sorghums in the latter study contained 21.3% less lysine (1.89 g/kg versus 2.40 g/kg; p < 0.015) and 25.7% less arginine (3.32 versus 4.47 g/kg; p < 0.005). Given that kafirin contains low levels of lysine and arginine, these reductions in concentrations of more than 20% are entirely consistent with the proposal that kafirin constitutes a greater proportion of total sorghum protein in the latter study.
This is a disadvantageous development because kafirin contains low levels of lysine but high levels of leucine. Moreover, kafirin is a poor source of digestible amino acids due to its inherent hydrophobicity and the structure of protein bodies. It is generally accepted disulphide cross-linking in the β- and γ-kafirin fractions, which are located in the periphery of the protein body, impedes the digestion of the central α-kafirin core and this is exacerbated by hydrothermal processes as they induce disulphide cross-linkages [41].
Clearly, kafirin compromises sorghum protein quality [2] but this may not be crucial as kafirin constitutes in the order of 15% of total protein in a sorghum-based broiler diet. However, if kafirin levels in local sorghums are indeed increasing then the real possibility that kafirin compromises starch/energy utilisation is pivotal. The mechanisms by which kafirin compromises starch utilisation have not been clarified but arguably their genesis stems from the close physical proximity of kafirin protein bodies, starch granules and glutelin protein matrices in sorghum endosperm. As discussed by Gidley, et al. [42] the quantity of starch in sorghum endosperm exceeds that of kafirin protein, which suggests that the magnitude of biophysical starch-protein interactions is capped. Intuitively, biochemical starch-protein interactions may hold more importance but despite the recognition of the importance [43] they have yet to be precisely defined.
The proposition that kafirin compromises starch/energy utilisation, as enunciated by Taylor [44], is generally accepted although reservations have been expressed by Gidley, et al. [42]. In sorghum endosperm the diameter of starch granules (~15 μm) is considerably larger than kafirin protein bodies (1–3 μm) and the endosperm contains considerably higher concentrations of starch than kafirin. Starch granules and kafirin protein bodies are both discrete structures in sorghum endosperm. The concept that hydrothermally induced disulphide cross-linkages in the cysteine-rich periphery of protein bodies impede the digestion of the central α-kafirin core is straightforward because α-kafirin would be “protected” by a layer of relatively insoluble protein. It is more difficult to conceptualise that these reactions are also a direct impediment to starch digestion, although they may physically hinder starch gelatinisation. Intuitively, it seems any interactions between starch and protein that might impede starch gelatinisation/digestion are more likely to occur between starch and the glutelin protein matrix because starch granules are embedded in the protein matrix. Another possible inherent impediment to starch digestion is the endosperm cell walls. Taylor [44] considered it was probable that these cell walls constitute something of a barrier to digestion. The cell walls are rich in insoluble glucuronoarabinoxylan type pentosans and are intimately bound to the glutelin protein matrix, quite possibly via ferulic acid, a phenolic compound that is abundant in sorghum.
De Mesa-Stonestreet, et al. [45] discussed that kafirins are located primarily in spherical protein bodies, which are embedded in a glutelin protein matrix, and the integrity of which is dependent on disulphide cross-linkages. This is allied to studies using three-dimensional fluorescence microscopy completed by Ezeogu, et al. [46] and their conclusion was that low starch digestibility of cooked sorghum flour was the result of “the more disulfide-bonded protein matrix”. Another possibility, as considered by Truong, et al. [24], are interactions involving disulphide linkages between β- and γ-kafirin on the periphery of protein bodies and starch-granule associated proteins on the surface of starch granules may have the capacity to impede starch digestion. Collectively, it seems possible that disulphide cross-linkages involving kafirin protein bodies, glutelin protein matrices and starch-granule associated proteins in sorghum endosperm are a set of connections forming a web-like protein network that impedes starch utilisation.
Following the quantification of kafirin concentrations in sorghum, a series of feeding studies completed by Poultry Research Foundation indicated that kafirin has negative impact on energy utilisation in broiler chickens (unpublished data). The methodology involved to quantify kafirin has been fully documented by Truong, et al. [47]. In these studies, kafirin significantly depressed ME:GE ratios (p < 0.04) and tended to decrease AMEn (p = 0.074) in broiler chickens from meta-analyses with non-significant (p > 0.10) experimental leverages (Table 1). Thus our tentative conclusion is kafirin content and composition compromises starch/energy utilisation in sorghum-based diets and the real possibility that the chicken-meat industry is being confronted with “high-kafirin” sorghums is a predicament that needs to be considered by sorghum breeding programs. Put simply, the availability of relatively low protein sorghums with lesser kafirin proportions should be substantially beneficial to growth performance and energy utilisation in broiler chickens.
Table 1. The relationship between dietary concentrations of kafirin and energy utilisation (ME:GE ratios and AMEn) in broiler chickens offered sorghum-based diets.
There has been an increasing focus on the inclusion of exogenous proteases in pig and poultry diets. More specifically, Taylor [44] suggested that proteases should be evaluated in sorghum-based broiler diets with the caveat that kafirin and glutelin are not readily hydrolysed by proteolytic enzymes. In this context it is relevant that most or all exogenous proteases are hydrolases and inherently lack the capacity to cleave disulphide cross-linkages which requires reductase activity.
Selle, et al. [48] reported that a Bacillus lichenformis-derived protease improved N digestibility in conventional, sorghum-based broiler diets that were steam-pelleted at an 80 °C conditioning temperature. At the standard inclusion rate (300 units/gram), this protease significantly increased N digestibility coefficients by 16.5% in the distal jejunum (0.627 versus 0.538), by 5.91% in the proximal ileum (0.770 versus 0.727) and by 7.09% in the distal ileum (0.770 versus 0.719). Subsequently, Liu, et al. [49] reported that this protease significantly improved the digestibility of arginine (4.63%), histidine (8.23%), isoleucine (9.87%), leucine (12.7%), phenylalanine (10.5%), threonine (7.36%), valine (10.4%), alanine (12.8%), aspartic acid (9.60%), glutamic acid (9.38%), glycine (9.73%), proline (14.6%), serine (11.5%), and tyrosine (12.1%). Only the responses in lysine and methionine digestibility were not significant and this was probably because the diets contained quite high levels of these two amino acids in “free” or synthetic forms. In addition, protease supplementation significantly (p < 0.05 - < 0.005) increased the rates of digestion of 12 of the 16 amino acids assessed. Moreover, as proposed by Liu and Selle [22], it is probable digestion rates of amino acids were more indicative of broiler performance than static amino acid digestibility coefficients.
Instructively, this protease also significantly increased the digestibility of starch in the distal jejunum by 13.6% (0.770 versus 0.678; p < 0.005) and by 4.80% (0.851 versus 0.812; p < 0.05) in the proximal ileum and numerically enhanced starch digestibility in the proximal jejunum (15.8%) and the distal ileum (3.15%). Based on the presumption that this enzyme preparation did not contain any starch-degrading side-activities, these findings are consistent with the proposition that kafirin and/or glutelin are interfering with starch digestion when sorghum-based diets are offered to poultry.
4.2. “Non-tannin” Polyphenolic Compounds and Phenolic Acids
Phenolic compounds are a diverse group of phytochemicals ranging from highly-polymerised inert lignins to simple C7-C9 phenolic acids [50] and their concentrations in sorghum (170–10,260 mg/100 g) are substantially more than maize (30.9 mg/100 g) and wheat (22–40 mg/100 g) [51]. The polyphenolic compound condensed tannin is an anti-nutritive factor that depresses broiler performance [52] and has been even described as a toxic factor [53]. Sorghum cultivars may be divided into three categories depending on their genotypes and condensed tannin contents [54]. Type I sorghums do not have a pigmented testa and are “tannin-free”, Type II sorghums have a pigmented testa layer that contains condensed tannin and Type III (“bird-proof”) sorghums contain condensed tannin in both the testa and the peripcarp. Dykes and Rooney [55] reported condensed tannin concentrations of 0.28, 4.48 and 11.95 g/kg in Type I, Type II and Type III sorghums, respectively. As discussed, it is most likely Australian sorghums no longer contain condensed tannin; however, there is the very real possibility non-tannin phenolic compounds negatively influence digestion of starch and protein to compromise growth performance in broiler chickens.
Taylor [44] concluded grain sorghum cultivars contain higher levels of phenolic compounds than other cereals and that red (non-tannin) sorghums are highly pigmented with polyphenols, anthocyanins and anthocyanidins and, importantly, these phenols bind strongly to starch. Also ferulic acid has been shown to influence starch pasting profiles which implies phenolic acids have the capacity to interact with starch [17]. Thompson and Yoon [56] found tannic acid depressed wheat starch digestion in vitro and Thompson, et al. [57] reported intakes of phenolic compounds by humans were negatively correlated with blood glycaemic indices.
Phenolic compounds and phytate are both located in the periphery of sorghum grain (pericarp and aleurone layer) rather than in the central endosperm where starch granules are located. Thus in native grain sorghum it seems unlikely any inter-reactions between phenolics and phytate with starch would take place; however, they may be initiated by processing (e.g., hammer-milling, steam-pelleting) and take place in the avian gut, perhaps particularly in the gizzard. Phenolic compounds appear to be susceptible to processing and extrusion has been shown to reduce the molecular weight of phenolic polymers in sorghum [58]. More recently, Dlamini, et al. [59] reported that extrusion reduced average total phenol contents by 51% (14.92 versus 7.37 mg catechin equivalents/g) in the whole grain of five sorghum varieties. The impact of decortication or dehulling was more profound with a reduction of 68% (14.92 versus 4.80 mg catechin equivalents/g) due to the location of phenolics in the outer layers of the grain. The effects of extrusion suggest steam-pelleting sorghum-based diets may reduce the size of phenolic polymers, which may influence any subsequent interactions between phenolics and starch in the avian gut.
Zhu [60] reported non-covalent interactions involving starch and phenolic compounds influence physicochemical and nutritional properties of feedstuffs. Earlier, Tomasik and Schilling [61] and Barros, et al. [62] opined that phenolics readily form starch complexes and are probably more likely to form starch-phenolic complexes with amylose than amylopectin. Kandil, et al. [63] reported that phenolic acids play an important role in the resistance of starch to hydrolysis in a study involving barley, maize, triticale and wheat but not sorghum. Welsch, et al. [64] found phenolic compounds were capable of inhibiting Na+-K+-ATPase or the “sodium pump” which suggested intestinal uptakes of nutrients including glucose via Na+-dependent transporter systems could be compromised.
A wide range of polyphenolic compounds and phenolic acids have been quantified in a number of diverse sorghum samples by this institute. As a consequence, our contention is that the anti-nutritive properties of phenolic compounds are not the sole domain of CT and to some extent are shared by the balance of polyphenols and phenolic acids found in grain sorghum. Instructively, Elkin, et al. [65] concluded that condensed tannin is only partially responsible for variations in the nutrient quality. Also, phenolic compounds and phytate appear to share several anti-nutritive properties [11]. Provisionally, there was a negative linear relationship (r = −0.569; p = 0.042) between dietary levels of total phenolic compounds and ME:GE ratios in broilers offered diets based on nine “tannin-free” sorghum varieties across five of feeding studies. This preliminary outcome supports the contention and prompts identification of which polyphenols or phenolic acids are responsible. Initial indications are that the flavan-4ols (polyphenolic compounds) and soluble conjugated and insoluble bound ferulic acid (a phenolic acid) might be involved.
The colour of red sorghums is attributable to polyphenolic pigments (anthocyanins) and axiomatically red sorghums contain more polyphenols than white sorghum varieties. Indeed, Beta and Corke [66] reported that four red sorghums had a greater concentration of total polyphenols than six white sorghums by more than a three-fold factor. In Australia probably all white sorghums are of the Liberty variety. Interestingly, we have found that Liberty contains lower concentrations of phenolic acids than a number of red sorghum varieties albeit in a limited number of samples. Three categories of phenolic acid concentrations were quantified, free and conjugated phenolic acids which are soluble and the insoluble, bound phenolic acids. In two projects, nine red sorghums contained an average of 500 μg/g total phenolic acids and 392 μg/g total ferulic acid; whereas, in contrast, two white Liberty sorghums contained 328 μg/g phenolic acids and 235 μg/g ferulic acid. Thus red sorghums contained 28% more phenolic acids and 40% more ferulic acid than white sorghums in this comparison. Ferulic acid is the dominant phenolic acid in sorghum, representing 78% of total phenolic acids across the 11 varieties; however, ferulic acid is certainly not unique to sorghum amongst grains as it is present in maize and wheat.
Dykes, et al. [67] reported a positive correlation between total phenols and flavan-4ols (r = 0.94; p < 0.001) in non-tannin sorghums. A recent study at this institute involved offering sorghum-based diets (580 g/kg) based on six different varieties to broiler chickens. In this study, flavan-4ols in sorghum per se was negatively correlated with ME:GE ratios (r = −0.919; p = 0.010) which supports the contention that “non-tannin” polyphenolic compounds in sorghum have the capacity to compromise energy utilisation in poultry.
4.3. Phytate
The inclusion of phytase in broiler diets is now a very common practice and the reciprocal impacts of dietary phytate and exogenous phytase on broiler performance have been reviewed extensively [16,20,68,69]. Briefly, to avoid repetition, sorghum invariably contains phytate at relative and absolute concentrations that are often somewhat higher than other cereal grains [19,70] and it is generally accepted that phytate negatively influences protein and energy utilisation in poultry [20,68,71]. The polyanionic IP6 phytate molecule may form binary or ternary complexes with proteins depending on the isoelectric point of protein and environmental pH [70]. Phytic acid may affect starch digestion by directly binding starch through phosphate linkages or indirectly via starch granule-associated proteins [72]. Also, phytate is a potent chelator of divalent cations including zinc and calcium. However, the fundamental role of phytase is to liberate phytate-bound P and this enhanced P utilisation is of astonishing environmental importance. Nevertheless, the suggestion is that the response of starch and protein digestion to phytase [73] in sorghum-based broiler diets appears to be less robust in comparison to maize- and wheat-based diets [74]. The likelihood is that the “extra-phosphoric” effects of phytase were largely a consequence of phytase attenuating protein-phytate complex formation.
As proposed by Cosgrove [14], electrostatic attractions between positively-charged arginine, histidine, and lysine residues with polyanionic phytate molecules are pivotal to the initiation of binary protein-phytate complex formation. This takes place at pH levels less than the isoelectric point of proteins and the isoelectric point (iP) of kafirin is 5.9 [75] which is considerably higher than pH levels usually found in the crop, proventriculus and gizzard. However, from first principles, it follows that phytate may not readily bind kafirin due to its paucity of basic amino acids. Also, the structure of the discrete, spherical kafirin protein body with its hydrophobic periphery may limit interactions with phytate. Both factors may in turn limit “extra-phosphoric” responses to exogenous phytases in sorghum-based diets simply because kafirin is not readily bound by phytate in binary complexes. It is also follows that if phenolic compounds are limiting performance in sorghum-based broiler diets, then phytase and other standard exogenous enzymes do not have the capacity to counteract this impact.
Both phytate and the various phenolic compounds are located in the outer layers of grain sorghum including the aleurone. Perhaps because of this we are consistently finding that concentrations of phytate and phenolics in sorghums are positively correlated. In this context it is relevant that Selle, et al. [11] gave consideration to the analogous anti-nutritive properties of phytate and phenolic compounds. As one related example, Kreydiyyeh, et al. [76] found that tea extracts reduced intestinal mucosal Na+ -K+ -ATPase or sodium pump activity and uptakes of glucose and sodium in rats. Subsequently, Kreydiyyeh [77] reported that tannic acid, a constituent of tea extracts, inhibited intestinal sodium pump activity in vitro and phenylalanine uptakes in rats. This was attributed to direct inhibition of Na+-K+-ATPase activity by tannic acid and dissipation of the Na gradient required for intestinal uptakes of phenylalanine. Alternatively, Liu, et al. [78] demonstrated the addition of 1000 FTU/kg phytase to corn-soy diets significantly increased Na+-K+-ATPase activity in the duodenum and jejunum of broilers by approximately 20%, and increased glucose concentrations in enterocytes. The implication is phytate depresses sodium pump activity and intestinal uptakes of glucose. Overall, the suggestion is that both phytate and phenolic compounds can impede sodium pump activity along the small intestine thereby compromising intestinal uptakes of glucose and amino acids via Na+ -dependent transport systems.
5. White versus Red Sorghum
White sorghums are considered to be superior to red sorghums by both pig and poultry nutritionists, which was reflected in our survey. One possibility is that higher concentrations of both polyphenolic compounds and phenolic acids in red sorghums are contributing to their apparent inferiority in comparison to white sorghums. While phenolic acids are not related to grain colour their relatively low concentrations in two samples of Liberty may hold relevance. Interestingly, flavan-4ols per se are not responsible for “redness” in sorghum but they are precursors of red polyphenolic pigments. Therefore, it follows that there would be less flavan-4ols in white than red sorghums and our analyses have confirmed this to be the case.
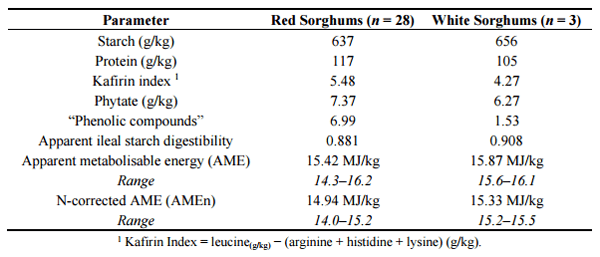
The majority of grain sorghums harvested in Australia possess red pericarps while the minority (Liberty) have white pericarps. This reflects a perception that red sorghums varieties are more viable agronomically than white sorghum and another reason is that white sorghums have a grey appearance following exposure to moisture. Perez-Maldonado and Rodrigues [4] offered broiler chickens diets based on 31 different sorghums including three white sorghums (Liberty) harvested in 2004 and 2005. As shown in Table 2, the mean AME of white sorghum was 0.45 MJ higher (15.87 versus 15.42 MJ/kg dry matter (DM)) than red sorghum and the AMEn was 0.39 MJ higher (15.33 versus 14.94 MJ/kg DM) across both 2004 and 2005 harvest years. A larger number of white sorghum samples would have been preferable; nevertheless, the differences in AME (p < 0.125) and AMEn (p < 0.05) either approached or were statistically valid. Thus the implication is that the energy/starch component of white sorghum is better utilised than red sorghums.
Table 2. Key characteristics of the sorghum varieties assessed by Perez-Maldonado and Rodrigues [4].
Selle, et al. [48] compared a white Liberty (98 g/kg protein) with a red Buster (121 g/kg protein) sorghum as the dietary basis for broiler chickens. Both sorghums were hammer-milled through a 3.2 mm screen prior to being incorporated into diets that were steam-pelleted at a conditioning temperature of 95 °C. Liberty supported better weight gains by 4.69% (1540 versus 1471 g/bird; p < 0.03) and feed conversion ratios by 2.88% (1.485 versus 1.529; p < 0.05) from 7 to 28 days post-hatch. These improvements in growth performance were associated with significantly higher starch digestibility coefficients in four small intestinal segments, higher starch digestion rates, a higher predicted glycaemic response and a greater quantity of potentially digestible starch. Also, from the same comparison, Liu, et al. [18] reported that white sorghum-based diet had significantly higher (p < 0.05) average amino acid digestibility coefficients than red sorghum-based diet in the proximal ileum (0.797 versus 0.769) and distal ileum (0.833 versus 0.806); white sorghum-based diet also had significantly higher apparent digestibility coefficients of leucine, alanine, glutamic acid and proline in the distal jejunum and 14.1% higher average amino acid digestion rate than red sorghum-based diets (2.47 versus 2.17 × 10−2 min−1 ). The more rapid protein digestion rate in white sorghum-based diets may have contributed to the better feed conversion efficiency via synchronizing starch and protein digestion and the availability of glucose and amino acids.
In this comparison, white sorghum contained less phytate (6.74 versus 9.22 g/kg) and less total phenolic compounds (2.02 versus 4.13 GAE/g) than the red sorghum (Buster). However, as discussed, non-tannin phenolic compounds may negatively influence protein and starch digestion and compromise growth performance in broiler chickens. Phenolic acids are found in the outer layers of the kernel (pericarp, testa, and aleurone), and the bound phenolic acids are associated with the cell walls [55]. The white sorghum contained less free, conjugated and bound phenolic acids (Figure 1) and less flavan-4ols, which may play an important role in mould resistance [55]. In broiler diets based on sorghum- or sorghum-wheat blends, sorghum is an important energy source and phenolic compounds may have impeded starch digestion and glucose absorption in this comparison.
Figure 1. The concentrations of free, conjugated and bound phenolic acids in white and red sorghum.
6. Starch and Protein Digestive Dynamics in Sorghum-Based Broiler Diets
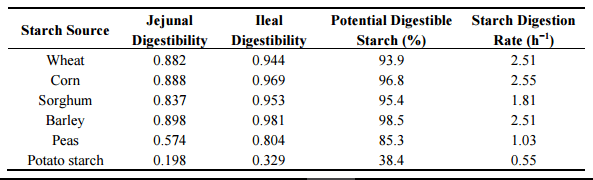
Accumulating evidence suggests that the provision of some slowly digestible starch in broiler diets is beneficial for feed conversion efficiency [79–81]. One possibility is that retarded starch digestion rates provide a greater supply of glucose to the lower small intestine which spares amino acids from catabolism for energy to meet the gut mucosa’s requirements thereby permitting the entry of amino acids into the systemic circulation to become available for protein deposition [22]. Based on an in vitro starch digestion evaluation in various cereal grains, Giuberti, et al. [23] reported that sorghum had the least rapidly digestible starch (119 g/kg), potential digestible starch (70.4 g/100 g dry starch) and the highest resistant starch (275 g/kg). The authors also showed that sorghum had a low starch digestion rate (0.018 min−1 ) and the lowest predicted glycaemic index (Table 3). This is consistent with findings of Weurding, et al. [82] where starch digestion rates in the small intestine of broiler chickens differ across feed ingredients and sorghum had the lowest and slowest starch digestion rate (Table 4).
Table 3. Starch fractions and digestion rates of starch in various cereal grains [23].
Table 4. Starch digestion characteristics in the small intestine of broilers fed diets containing different sources of starch [82].
Liu, et al. [83] found starch and nitrogen digestive dynamics were more indicative of feed conversion efficiency and weight gain than their static ileal digestibility coefficients. These findings suggest that the rate of amino acids absorption in the small intestine in relation to glucose absorption may have a tangible bearing on protein deposition and feed conversion efficiency. Similar investigations have been conducted in pigs; van den Borne, et al. [84] defined nutrient asynchrony as a partial temporal separation of amino acid and glucose availability and assessed the effects of synchronicity of amino acids and glucose on protein and energy metabolism. They found that the efficiency of protein utilisation was significantly enhanced in so-called synchronous diets (56.7%) in comparison to asynchronous diets (47.1%). It was concluded that the asynchronous availability of glucose and amino acids increased amino acid oxidation and compromised protein utilisation. Because sorghum has relatively slower starch digestion than maize and wheat, it may have the potential to provide a diet with better balanced digestion of starch and protein. However, accelerating protein digestion and amino acid absorption in sorghum-based diets is a challenge which may be met by inclusions of sulfite reducing agents and exogenous proteases.
7. Inclusions of Sulfite Reducing Agents
The negative impact of “moist-heat” or “wet-cooking” on sorghum protein digestibility is recognised. Wet cooking has been shown to reduce in vitro protein digestibility from 78%–100% to 45%–55% [85]. This may be associated with the formation of disulphide cross-linkages in kafirin coupled with its inherent hydrophobicity and secondary protein structural changes [11]. Kafirin is less soluble than zein in maize and gluten in wheat [13]. The observed lower in vitro protein digestibility of wet-cooked sorghum in comparison to maize and wheat may be related to the structure of the kafirin protein body. The formation of disulphide cross-linkages in the periphery (β- and γ-kafirin) may generate a relatively insoluble protein barrier which “protects” the central core (α-kafirin) from digestion by limiting the access of enzymes. The inclusion of reducing agents (dithiothreitol and sodium bisulphite) with sorghum during wet-cooking has been shown to significantly improve protein digestion [86]. Interestingly, disulphide-mediated polymerisation of zein formed in cooked maize did not result in reduced protein digestibility [87]; this, coupled with protein body structure, may be attributed to the formation of more disulphide-bounded protein oligomers in sorghum than in maize [88].
Sulphite reducing agents, including sodium sulphite (Na2SO3), sodium bisulphite (NaHSO3) and sodium metabisulphite (Na2S2O5), have the capacity to cleave disulphide cross-linkages and have been shown to improve in vitro starch and protein digestibility in sorghum [86,89,90]. Selle, et al. [91] reported that sodium metabisulphite (SMBS) significantly decreased disulphide cross-linkages and increased free sulphydryl groups in vitro. Moreover, in vivo, SMBS significantly enhanced energy utilisation as it increased AME by up to 0.56 MJ in broilers offered sorghum-based diets containing a range of SMBS inclusion levels. Subsequently, Selle, et al. [92] found that graded SMBS inclusions linearly decreased concentrations of disulphide bonds (r = −0.775; p < 0.001), linearly increased free sulphydryl groups (r = 0.890; p < 0.001) and protein solubility of the diets (r = 0.943; p < 0.001). In the same study, the inclusion of 1.50 g/kg SMBS in conventional sorghum-based broiler diets significantly improved AME by 0.36 MJ (13.81 versus 13.45 MJ/kg; p = 0.005), AMEn by 0.45 MJ (12.30 versus 11.85 MJ/kg; p = 0.001) and feed conversion ratio (FCR) by 2.47% (1.422 versus 1.458; p = 0.025).
From the preliminary study [91], it appears broilers chickens were able to tolerate dietary SMBS inclusions approaching 5.00 g/kg before feed intakes are depressed; however, inclusion levels under 5.00 g/kg have the capacity to enhance nutrient utilisation of sorghum-based diets. With the exception of one sorghum (the red sorghum evaluated in the Truong, et al. [93] study), responses in energy utilisation (AME, ME:GE ratio and AMEn) to SMBS inclusions up to 5.25 g/kg were all positive across five studies as shown in Table 5. In studies 2 to 5 inclusive (complete sorghum-based diets as opposed to sorghum-cased diets used in study 1), the average SMBS inclusion of 2.84 g/kg increased AME by 0.27 MJ, AMEn by 0.32 MJ, and ME:GE ratios by 1.90%. SMBS also improved FCR in broilers offered complete sorghum-based diets by an average of 1.37% or a median response of 1.47%. It would be advantageous to ascertain if these improvements stem mainly from reductions of disulphide cross-linkages and increased solubility of proteins, perhaps especially kafirin, or from starch depolymerisation [94,95]. Given the importance placed on starch-protein interactions in sorghum specifically [43], and in general [24], this is only complicated by the likelihood that the two impacts are interdependent. There is the real possibility SMBS modifies both starch and protein digestive dynamics to benefit energy utilisation in broiler chickens [22]. It seems likely that SMBS responses with certain grain sorghum varieties will be more pronounced than others. Inclusions of SMBS displays promise in commercial practice as it has positive effects on energy utilisation and efficiency of feed conversion and its dietary inclusion cost is largely negated by its replacement of sodium bicarbonate to maintain dietary sodium at intended levels.
Table 5. Responses to sodium metabisulphite (SMBS) inclusions of up to 5.25 g/kg on energy utilisation parameters and feed conversion ratio (FCR) in five studies conducted by Poultry Research Foundation involving a total of eight sorghum varieties1 .
8. Responses of Exogenous Feed Enzymes
The inclusion of exogenous enzymes in sorghum-based diets have been reviewed by Selle, et al. [11] and Liu, et al. [3]. It is generally accepted that phytate negatively influences protein utilisation in poultry [20,68]. This is reflected in the Ravindran, et al. [96] study where Aspergillus niger phytase (1200 FTU/kg) increased mean apparent ileal digestibility (AID) coefficients of 14 amino acids by 6.46% (0.791 versus 0.743) in broilers offered “all-sorghum” diets. However, the low protein content of the sorghum (73.3 g/kg) used in this study probably indicates a relatively small kafirin proportion of total protein. There is the suggestion that the “extra-phosphoric” responses to phytase in sorghum are less pronounced than in other grains [74]. As proposed by Cosgrove [14], at pH levels less than the isoelectric point of protein, electrostatic attractions between positively-charged arginine, histidine, and lysine residues with polyanionic phytate molecules are crucial to binary protein-phytate complex formation. If so, it follows that phytate would not readily bind kafirin due to its paucity of basic amino acids. This may in turn limit “extra-phosphoric” responses to exogenous phytases in sorghum-based diets.
A study completed by the Poultry Research Foundation in 2013 evaluated the effects of an Escherichia coli-derived phytase (1000 FTU/kg) on starch digestibility coefficients in four small intestinal sites in broilers offered conventional sorghum-based diets steam-pelleted at 70 °C. Phytase significantly increased average starch digestibility coefficients by 7.27% (0.812 versus 0.757; p < 0.04) in the four small intestinal sites. Subsequently, Liu, et al. [7] showed that there were numerical improvements in weight gain, feed conversion efficiency and N retention in sorghum-based diets following phytase supplementation (1000 FTU/kg). Truong, et al. [93] found that phytase did not significantly influence nutrient utilisation and apparent digestibility of starch and nitrogen along the small intestine in birds offered sorghum-based diets. Truong, et al. [21] reported that phytase (1000 FTU/kg) increased apparent disappearance rates of phytate in four small intestinal segments which indicates that phytase has the capacity to degrade phytate and generate “phosphoric responses” in sorghum-based diets. Thus phytase inclusions in sorghum-based diets have the potential to reduce levels of inorganic P source and reduce feed costs but the “extra-phosphoric” responses may be somewhat muted.
Investigations into the inclusion of other exogenous enzymes in sorghum-based diets are limited. Liu, et al. [49] reported that a Bacillus lichenformis-derived protease significantly increased the digestibility of 13 amino acids in distal ileum, which ranged from a 4.5% increase in histidine (0.820 versus 0.785, p < 0.05) to a 7.9% increase in proline (0.779 versus 0.722, p < 0.01); it also significantly increased digestion rates of 12 amino acids from 34.2% for histidine (2.57 versus 3.45 × 10−2 min−1 , p < 0.05) to 55.7% for proline (2.28 versus 3.55 × 10−2 min−1 , p < 0.01). Taylor [44] suggested that kafirin may not be readily degraded due to its poor solubility and disulphide linkages. The positive impact of this protease on amino acid digestibility in sorghum-based diets is encouraging; ideally, it should be confirmed in further studies.
Liu, et al. [97] examined an enzyme combination of xylanase, amylase and protease in maize-, wheat- and sorghum- based broiler diets and found that enzyme supplementation did not influence growth performance and nutrient utilization but significantly increased apparent digestibility coefficients of starch in the proximal ileum and apparent digestibility coefficients of nitrogen in the distal jejunum and proximal ileum. Although sorghum contains low concentrations of soluble non-starch polysaccharide (NSP) and is considered to be a “non-viscous” grain, it is often combined with wheat in broiler diets to enhance pellet quality and NSP degrading enzymes should have the potential to improve growth performance and nutrient utilisation in sorghum-wheat blended diets. However, few studies have investigated this issue and future studies are required to determine the effects of exogenous enzymes on sorghum-wheat-blend diets in broiler chickens.
9. Innovative Assessments of Sorghum Quality
The capacity to assess the quality of sorghum as a feedstuff for broiler chickens is clearly a desirable objective. Therefore, in this section consideration is given to three approaches which, in concert, may enable nutritionists to rank the quality of a sorghum shipment and formulate diets accordingly to maximise broiler performance.
9.1. Promatest Protein Solubility
Odjo, et al. [98] has described the Promatest procedure to determine the salt-soluble protein content of maize. Briefly, the solubility of extracted protein is compared to the standard of bovine serum albumin via a colorimetric reaction with coomassie brilliant blue G250 (Merck, Germany). Importantly, Gehring, et al. [99] found that Promatest protein solubilities of 12 maize samples were positively correlated with N-corrected AME (r = 0.81) and ileal digestible energy (r = 0.48) to significant (p < 0.01) extents in broiler chickens. The determination of salt-soluble protein contents in sorghum cultivars in relation to broiler performance could be considered to be innovative and our preliminary investigations are encouraging. For example, Selle, et al. [100] found a significant correlation between protein solubilities of diets based on three sorghum varieties with red, white and yellow pericarps offered to broiler chickens as either unprocessed mash or as re-ground mash following steam-pelleting at a conditioning temperature of 90 °C with N retention, as shown in Figure 2. Overall steam-pelleting reduced protein solubility from 56.3% to 38.3% with a corresponding reduction in N retention from 60.6% to 55.0%. Moreover, Promatest protein solubilities were positively correlated with concentrations of free sulphydryl groups (r = 0.585, p < 0.001) and negatively correlated with disulfide cross-linkages (r = −0.518, p < 0.001).
Figure 2. Linear relationship (r = 0.881 p = 0.021) between protein solubility and N retention in broiler chickens offered sorghum-based diets (adapted from Selle, et al. [100]).
The majority broiler diets are steam-pelleted and sorghum is known to be vulnerable to wet-cooking or hydrothermal processing [41]. Hamaker, et al. [89] reported that wet-cooking resulted a 30.3% reduction of in vitro pepsin digestibility in sorghum in comparison to 13.9% in barley, 10.0% in rice, 5.9% in wheat and 4.9% in maize. Duodu, et al. [13] suggested that this could be due to the factors including disulfide and non-disulfide cross-linking and alterations to the secondary structure of sorghum protein. Given this, it appears that Promatest protein solubility may hold special relevance for sorghum in poultry.
Subsequently, in red sorghum-based diets, Selle, et al. [48] reported that Promatest protein solubility was positively correlated with concentrations of free sulphydryl groups (r = 0.720, p < 0.01). In an investigation of including SMBS into broiler diets, Selle, et al. [92] found that graded inclusions of SMBS linearly decreased disulphide bonds (r = −0.775, p < 0.001) and increased concentrations of free sulphydryl groups (r = 0.890, p < 0.001) and protein solubility (r = 0.943, p < 0.001). Also, protein solubility was correlated with the concentrations of both disulphide bonds and free sulphydryl groups in this study. Therefore, it should be possible to apply Promatest protein solubility as an indicator of protein quality in sorghum per se and sorghum-based broiler diets. The correlation between protein solubility and N retention is promising; nevertheless, further research is required to determine the relevance of Promatest protein solubility with in vivo growth performance and nutrient utilisation in broiler chickens.
9.2. RVA Starch Pasting Profiles
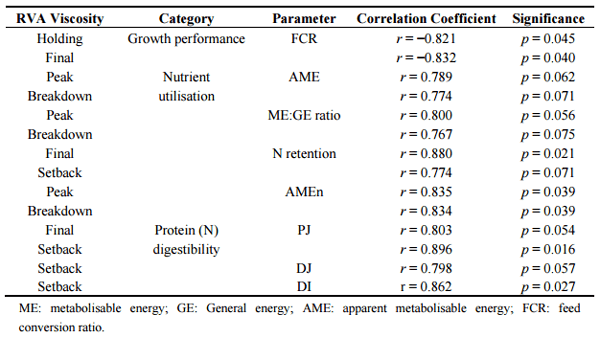
Starch is the major energy source in diets for broiler chickens; therefore, starch pasting profiles of the constituent grain and/or complete diets could be relevant to broiler performance. RVA starch pasting profiles are used extensively for human foods but they have not been widely applied to animal nutrition and their relevance is considered to be somewhat questionable. However, in an as yet unpublished study, sorghum-based diets were offered to broiler chickens and RVA viscosities of the constituent sorghum varieties were found to be correlated with various parameters of growth performance, nutrient utilisation and protein digestibility as documented in Table 6. Here, sorghum-SBM based diets were offered to broiler chickens from 7 to 28 days post-hatch and digesta samples were collected at the end of the study to determine digestibility coefficients in the proximal jejunum, distal jejunum, proximal ileum and distal ileum. Importantly, holding and final RVA viscosities were negatively correlated to feed conversion efficiency to significant extents; thus the implication is as final RVA viscosities of sorghum grains increase FCR is enhanced. This is reflected in the positive correlations between RVA viscosities and parameters of nutrient utilisation and protein digestibility where five observations were significant (p < 0.05) and a further seven approached significance (p < 0.10).
Table 6. Correlations between rapid visco-analyser (RVA) starch pasting profiles and parameters of growth performance, nutrient utilisation and protein digestibility coefficients in broiler chickens (PJ, proximal jejunum; DJ, distal jejunum; DI, distal ileum).
Proteins with disulfide bonds may restrict starch granule swelling during gelatinization, and Zhang and Hamaker [101] reported that dithiothreitol, a reducing agent, treatment of sorghum flour influenced RVA pasting profiles. Moreover, phenolic compound was also reported to influence starch pasting properties [17]. Instructively, Liu, et al. [102] showed that SMBS significantly (p < 0.001) reduced all five starch viscosity parameters and graded SMBS supplementation linearly reduced final starch viscosities (r = −0.986; p < 0.001). The profound impact of SMBS on RVA starch pasting profiles may be attributed directly to the depolymerisation of starch polysaccharides and indirectly to the reduction of disulphide cross-linkages in kafirin, glutelin and possibly starch granule-associated proteins. Therefore, it does appear that RVA starch pasting profiles will provide innovative and valid assessments of sorghum quality.
9.3. Grain Texture: Symes Particle Size Index (PSI)
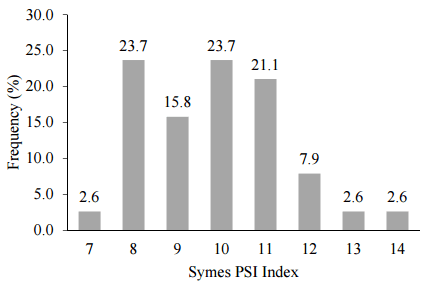
Symes [37] established the particle size index methodology to determine grain texture or “hardness” in wheat. Assessments of grain texture are not straightforward in practice and we have adopted the Symes PSI method to determine the hardness of sorghum [48,100,103]. Poultry Research Foundation has determined the Symes texture of 38 sorghum varieties in Australia as shown in Figure 3. The mean PSI value was 9.84 ± 1.603 (co-efficient of variation: 16.3%) which is at the harder end of the “very hard” spectrum of 8 to 12 inclusive and only three varieties fell outside this category.
Figure 3. Frequency of Symes particle size index (PSI) texture across 38 Australian sorghums.
On the basis of this method Australian sorghums are noticeably “harder” with lower PSI values than those from an extensive global survey recorded by de Alencar Figueiredo, et al. [38]. In this survey the mean texture of 117 sorghum samples was in the order of 12.7% but 44% of samples had softer textures than “very hard” as opposed to 5.3% (2 ex 38) in Australian sorghums. However, the likelihood is that “soft” sorghums are superior in performance and nutrient utilisation to “hard” sorghums as the basis for broiler diets. For example, Cabrera, et al. [39] reported that broilers offered diets based on a soft sorghum with mean particle sizes of 1000, 800 and 600 μm outperformed those offered hard sorghum by 7.14% in weight gain (945 versus 882 g/bird) and 4.38% in FCR (1.639 versus 1.714) in a 21-day feeding study. Kafirin is associated with hard textures of grain sorghums and in 13 sorghum varieties with quantified kafirin concentrations evaluated by Poultry Research Foundation, there was a relationship (r = −0.546; p = 0.054) between kafirin and harder textures that closely approached significance (Figure 4). Chiremba, et al. [104] used a tangential abrasive dehulling device was used to determine grain texture and reported that hard textures were positively correlated with concentrations of phenolic acids in maize and sorghum. This is a potentially important outcome but we have not detected similar relationships between total phenolic acids or ferulic acid with Symes PSI texture. This is may be because the limited number of sorghums (13 sorghums) that have been examined, future research is required to clarify the relationship between sorghum grain texture and intrinsic factors, including phenolic acids and kafirin.
Figure 4. Linear relationship (r = −0.546; p = 0.054) between kafirin concentrations and grain texture (Symes PSI) in 13 Australian sorghums.
10. Implications
Sorghum may be a conundrum as a feedstuff for chicken-meat production because of the relatively inefficient utilisation of dietary energy it provides but the likelihood is that kafirin and non-tannin phenolic compounds may be largely responsible for this situation. Therefore, the real possibility that kafirin levels in local sorghums are increasing as an inadvertent consequence of breeding programs should be addressed. Another viable approach would be the production of more white sorghum varieties with lower concentrations of “non-tannin” phenolic compounds. White sorghums with low protein and kafirin levels and axiomatically low phenolic compound levels should be a highly acceptable feedstuff. The assessment of the nutritive value of a delivery of grain sorghum to a feed-mill is an interesting challenge. However, it seems likely that a combination of Promatest protein solubilities, RVA starch pasting profiles and Symes PSI grain textures would be indicative and certainly merit further investigation as innovative approaches to meet this challenge. Sorghum may be a feed grain with issues for chicken-meat production but this could be, and should be, rectified.
Acknowledgments
The authors gratefully acknowledge the financial support received from the Chicken-meat Committee of the Rural Industries Research and Development Corporation (RIRDC) for the series of sorghum projects entitled “Steam pelleting temperature of sorghum-based broiler diets” (PRJ-003810), “Evaluation of sodium bisulphite in sorghum-based broiler diets” (PRJ-007639) and “The factors influencing sorghum starch digestibility in broiler chickens” (PRJ-008695).
Author Contributions
Sonia Y Liu reviewed the literature, drafted and revised this manuscript; Glen Fox contributed to the discussions on grain texture and functional properties in sorghum; Ali Khoddami contributed to determination of phenolic compounds in sorghum; Karlie A Neilson contributed to quantification of kafirin in sorghum; Ha H Truong and Amy F Moss contributed to collection of growth performance resultes in broiler chickens offered sorghum-based diets in the series of experiemnts; Peter H Selle revised and finalised this manuscript.
Conflicts of Interest
The authors declare no conflict of interest
References
1. Black, J.L.; Hughes, R.J.; Nielsen, S.G.; Tredrea, A.M.; MacAlpine, R.; van Barneveld, R.J. The energy value of cereal grains, particularly wheat and sorghum, for poultry. Proc. Aust. Poult. Sci. Symp. 2005, 17, 21–29.
2. Selle, P.H. The protein quality of sorghum. Proc. Aust. Poult. Sci. Symp. 2011, 22, 147–160.
3. Liu, S.Y.; Selle, P.H.; Cowieson, A.J. Strategies to enhance the performance of pigs and poultry on sorghum-based diets. Anim. Feed Sci. Technol. 2013, 181, 1–14.
4. Perez-Maldonado, R.A.; Rodrigues, H.D. Nutritional Characteristics of Sorghums from Queensland and New South Wales for Chicken Meat Production; RIRDC Publication No. 09/170; Rural Industries Research and Development Corporation: Barton, Australia, 2009.
5. Bryden, W.L.; Liu, X.; Ravindran, G.; Hew, L.I.; Ravindran, V. Ileal digestible amino acids values in feedstuffs for poultry; RIRDC Publication No. 09/71; Rural Industries Research and Development Corporation: Barton, Australia, 2009.
6. Waniska, R.D.; Hugo, L.F.; Rooney, L.W. Practical methods to determine the presence of tannins in sorghum. J. Appl. Poult. Res. 1992, 1, 122–128.
7. Liu, S.Y.; Cadogan, D.J.; Péron, A.; Truong, H.H.; Selle, P.H. Effects of phytase supplementation on growth performance, nutrient utilisation and digestive dynamics of starch and protein in broiler chickens offered maize-, sorghum- and wheat-based diets. Anim. Feed Sci. Technol. 2014, 197, 164–175.
8. Ao, Z.; Choct, M. Effect of early feeding and grain type on growth and performance of broilers. Proc. Aust. Poult. Sci. Symp. 2004, 16, 116–119.
9. Selle, P.H.; Cadogan, D.J.; Ru, Y.J.; Partridge, G.G. Impact of exogenous enzymes in sorghum- or wheat-based broiler diets on nutrient utilization and growth performance. Int. J. Poult. Sci. 2010, 9, 53–58.
10. Bryden, W.L.; Selle, P.H.; Cadogan, D.J.; Liu, X.; Muller, N.D.; Jordan, D.R.; Gidley, M.J.; Hamilton, W.D. A review of the nutritive value of sorghum for broilers; RIRDC Publication No. 09/007; Rural Industries Research and Development Corporation: Barton, Australia, 2009.
11. Selle, P.H.; Cadogan, D.J.; Li, X.; Bryden, W.L. Implications of sorghum in broiler chicken nutrition. Anim. Feed Sci. Technol. 2010, 156, 57–74.
12. Belton, P.S.; Delgadillo, I.; Halford, N.G.; Shewry, P.R. Kafirin structure and functionality. J. Cereal Sci. 2006, 44, 272–286.
13. Duodu, K.G.; Taylor, J.R.N.; Belton, P.S.; Hamaker, B.R. Factors affecting sorghum protein digestibility. J. Cereal Sci. 2003, 38, 117–131.
14. Cosgrove, D.J. Chemistry and biochemistry of inositol polyphosphates. Rev. Pure Appl. Chem. 1966, 16, 209–224.
15. Mosse, J.; Huet, J.C.; Baudet, J. The amino-acid composition of whole sorghum grain in relation to its nitrogen-content. Cereal Chem. 1988, 65, 271–277.
16. Selle, P.H.; Cowieson, A.J.; Cowieson, N.P.; Ravindran, V. Protein-phytate interactions in pig and poultry nutrition: A reappraisal. Nutr. Res. Rev. 2012, 25, 1–17.
17. Beta, T.; Corke, H. Effect of ferulic acid and catechin on sorghum and maize starch pasting properties. Cereal Chem. 2004, 81, 418–422.
18. Liu, S.Y.; Selle, P.H.; Cowieson, A.J. Influence of white- and red-sorghum varieties and hydrothermal component of steam-pelleting on digestibility coefficients of amino acids and kinetics of amino acids, nitrogen and starch digestion in diets for broiler chickens. Anim. Feed Sci. Technol. 2013, 186, 53–63.
19. Selle, P.H.; Walker, A.R.; Bryden, W.L. Total and phytate-phosphorus contents and phytase activity of australian-sourced feed ingredients for pigs and poultry. Aust. J. Exp. Agric. 2003, 43, 475–479.
20. Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41.
21. Truong, H.H.; Yu, S.; Peron, A.; Cadogan, D.J.; Khoddami, A.; Roberts, T.H.; Liu, S.Y.; Selle, P.H. Phytase supplementation of maize-, sorghum- and wheat-based broiler diets, with identified starch pasting properties, influences jejunal and ileal digestibilities of phytate (IP6) and sodium. Anim. Feed Sci. Technol. 2014, 198, 248–256.
22. Liu, S.Y.; Selle, P.H. A consideration of starch and protein digestive dynamics in chicken-meat production. Worlds Poult. Sci. J. 2015, 71, 297–310.
23. Giuberti, G.; Gallo, A.; Cerioli, C.; Masoero, F. In vitro starch digestion and predicted glycemic index of cereal grains commonly utilized in pig nutrition. Anim. Feed Sci. Technol. 2012, 174, 163–173.
24. Truong, H.H.; Liu, S.Y.; Selle, P.H. Starch utilisation in chicken-meat production: The foremost influential factors. Anim. Prod. Sci. 2015, doi:10.1071/AN15056.
25. Sang, Y.J.; Bean, S.; Seib, P.A.; Pedersen, J.; Shi, Y.C. Structure and functional properties of sorghum starches differing in amylose content. J. Agric. Food Chem. 2008, 56, 6680–6685.
26. Wong, J.H.; Lau, T.; Cai, N.; Singh, J.; Pedersen, J.F.; Vensel, W.H.; Hurkman, W.J.; Wilson, J.D.; Lemaux, P.G.; Buchanan, B.B. Digestibility of protein and starch from sorghum (Sorghum bicolor) is linked to biochemical and structural features of grain endosperm. J. Cereal Sci. 2009, 49, 73–82.
27. Taylor, J.R.N.; Emmambux, M.N. Developments in our understanding of sorghum polysaccharides and their health benefits. Cereal Chem. 2010, 87, 263–271.
28. Svihus, B.; Uhlen, A.K.; Harstad, O.M. Effect of starch granule structure, associated components and processing on nutritive value of cereal starch: A review. Anim. Feed Sci. Technol. 2005, 122, 303–320.
29. Ertl, D.; Dale, N. The metabolizable energy of waxy vs. Normal corn for poultry. J. Appl. Poult. Res. 1997, 6, 432–435.
30. Pirgozliev, V.R.; Rose, S.P.; Bedford, M.R. The effect of amylose: Amylopectin ratio in dietary starch on growth performance and gut morphology in broiler chickens. Arch. Fur Geflugelkunde 2010, 74, 21–29.
31. Collins, N.E.; Moran, E.T.; Stilborn, H.L. Performance of broilers fed normal and waxy corn diets formulated with chick and rooster derived apparent metabolizable energy values for the grains. J. Appl. Poult. Res. 2003, 12, 196–206.
32. Gilding, E.K.; Frere, C.H.; Cruickshank, A.; Raba, A.K.; Prentis, P.J.; Mudge, A.M.; Mace, E.S.; Jordan, D.R.; Godwin, I.D. Allelic variation at a single gene increases food value in a drought-tolerant staple cereal. Nat. Commun. 2013, doi:10.1038/ncomms2450.
33. Virupaksha, T.K.; Sastry, L.V.S. Studies on protein content and amino acid composition of some varieties of grain sorghum. J. Agric. Food Chem. 1968, 16, 199–203.
34. Taylor, J.R.N.; Schussler, L.; Vanderwalt, W.H. Fractionation of proteins from low-tannin sorghum grain. J. Agric. Food Chem. 1984, 32, 149–154.
35. Selle, P.H.; Liu, S.Y.; Cowieson, A.J. Sorghum: An enigmatic grain for chicken-meat production. In Sorghum: Production, Growth Habits and Health Benefits; Parra, P.C., Ed.; Nova Publishers Inc.: Hauppauge, NY, USA, 2013; pp. 1–44.
36. Henzell, R.G. Grain sorghum breeding in Australia: Current status and future prospects. In Proceedings of the 2nd Australian Sorghum Conference, Gatton, Australia, 4–6 February 1992; pp. 70–80.
37. Symes, K.J. Inheritance of grain hardness in wheat as measured by particle size index. Aust. J. Agric. Res. 1965, 16, 113–123.
38. De Alencar Figueiredo, L.F.; Davrieux, F.; Fliedel, G.; Rami, J.F.; Chantereau, J.; Deu, M.; Courtois, B.; Mestres, C. Development of nirs equations for food grain quality traits through exploitation of a core collection of cultivated sorghum. J. Agric. Food Chem. 2006, 54, 8501–8509.
39. Cabrera, M.R.; Hancock, J.D.; Bramel-Cox, P.J.; Hines, R.H.; Behnke, K.C. Effects of corn, sorghum genotype, and particle size on milling characteristics and growth performance in broiler chicks. Poult. Sci. 1994, 73, 11.
40. Ravindran, V.; Hew, L.I.; Bryden, W.L. Digestible amino acids in poultry feedstuffs; RIRDC Publication No. 98/9; Rural Industries Research and Development Corporation: Barton, Australia, 1998.
41. Selle, P.H.; Gill, R.J.; Downing, J.A. The vulnerability of sorghum to “moist-heat”. Proc. Aust. Poult. Sci. Symp. 2010, 21, 68–71.
42. Gidley, M.J.; Flanagan, B.M.; Sharpe, K.; Sopade, P.A. Starch digestion in monogastrics— Mechanisms and opportunities. Proc. Recent Adv. Anim. Nutr. Aust. 2011, 18, 207–213.
43. Rooney, L.W.; Pflugfelder, R.L. Factors affecting starch digestibility with special emphasis on sorghum and corn. J. Anim. Sci. 1986, 63, 1607–1623.
44. Taylor, J.R.N. Non-starch polysaccharides, protein and starch: Form function and feed-highlight on sorghum. Proc. Aust. Poult. Sci. Symp. 2005, 17, 9–16.
45. De Mesa-Stonestreet, N.J.; Alavi, S.; Bean, S.R. Sorghum proteins: The concentration, isolation, modification, and food applications of kafirins. J. Food Sci. 2010, 75, R90–R104.
46. Ezeogu, L.I.; Duodu, K.G.; Taylor, J.R.N. Effects of endosperm texture and cooking conditions on the in vitro starch digestibility of sorghum and maize flours. J. Cereal Sci. 2005, 42, 33–44.
47. Truong, H.H.; Neilson, K.A.; McInerney, B.V.; Khoddami, A.; Roberts, T.H.; Liu, S.Y.; Selle, P.H. Performance of broiler chickens offered nutritionally-equivalent diets based on two red grain sorghums with quantified kafirin concentrations as intact pellets or re-ground mash following steam-pelleting at 65 or 97 °C conditioning temperatures. Anim. Nutr. 2015, in press.
48. Selle, P.H.; Liu, S.Y.; Cai, J.; Cowieson, A.J. Steam-pelleting temperatures, grain variety, feed form and protease supplementation of mediumly-ground, sorghum-based broiler diets: Influences on growth performance, relative gizzard weights, nutrient utilisation, starch and nitrogen digestibility. Anim. Prod. Sci. 2013, 53, 378–387.
49. Liu, S.Y.; Selle, P.H.; Court, S.G.; Cowieson, A.J. Protease supplementation of sorghum-based broiler diets enhances amino acid digestibility coefficients in four small intestinal sites and accelerates their rates of digestion. Anim. Feed Sci. Technol. 2013, 183, 175–183.
50. Mangan, J.L. Nutritional effects of tannins in animal feeds. Nutr. Res. Rev. 1988, 1, 209–231.
51. Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333.
52. Nyachoti, C.M.; Atkinson, J.L.; Leeson, S. Sorghum tannins: A review. Worlds Poult. Sci. J. 1997, 53, 5–21.
53. McClymont, G.L.; Duncan, D.C. Studies on nutrition of poultry. III. Toxicity of grain sorghum for chickens. Ausi. Vet. J. 1952, 28, 229–233.
54. Asquith, T.N.; Izuno, C.C.; Butler, L.G. Characterization of the condensed tannin (proanthocyanidin) from a group II sorghum. J. Agric. Food Chem. 1983, 31, 1299–1303.
55. Dykes, L.; Rooney, L.W. Sorghum and millet phenols and antioxidants. J. Cereal Sci. 2006, 44, 236–251.
56. Thompson, L.U.; Yoon, J.H. Starch digestibility as affected by polyphenols and phytic acid. J. Food Sci. 1984, 49, 1228–1229.
57. Thompson, L.U.; Yoon, J.H.; Jenkins, D.J.A.; Wolever, T.M.S.; Jenkins, A.L. Relationship between polyphenol intake and blood-glucose response of normal and diabetic individuals. Am. J. Clin. Nutr. 1984, 39, 745–751.
58. Awika, J.M.; Dykes, L.; Gu, L.W.; Rooney, L.W.; Prior, R.L. Processing of sorghum (Sorghum bicolor) and sorghum products alters procyanidin oligomer and polymer distribution and content. J. Agric. Food Chem. 2003, 51, 5516–5521.
59. Dlamini, N.R.; Taylor, J.R.N.; Rooney, L.W. The effect of sorghum type and processing on the antioxidant properties of African sorghum-based foods. Food Chem. 2007, 105, 1412–1419.
60. Zhu, F. Interactions between starch and phenolic compound. Trends Food Sci. Technol. 2015, 43, 129–143.
61. Tomasik, P.; Schilling, C.H. Complexes of starch with inorganic guests. Adv. Carbohydr. Chem. Biochem. 1998, 53, 263–343.
62. Barros, F.; Awika, J.M.; Rooney, L.W. Interaction of tannins and other sorghum phenolic compounds with starch and effects on in vitro starch digestibility. J. Agric. Food Chem. 2012, 60, 11609–11617.
63. Kandil, A.; Li, J.H.; Vasanthan, T.; Bressler, D.C. Phenolic acids in some cereal grains and their inhibitory effect on starch liquefaction and saccharification. J. Agric. Food Chem. 2012, 60, 8444–8449.
64. Welsch, C.A.; Lachance, P.A.; Wasserman, B.P. Dietary phenolic-compounds—Inhibition of Na+-dependent D-glucose uptake in rat intestinal brush-border membrane-vesicles. J. Nutr. 1989, 119, 1698–1704.
65. Elkin, R.G.; Freed, M.B.; Hamaker, B.R.; Zhang, Y.; Parsons, C.M. Condensed tannins are only partially responsible for variations in nutrient digestibilities of sorghum grain cultivars. J. Agric. Food Chem. 1996, 44, 848–853.
66. Beta, T.; Corke, H. Genetic and environmental variation in sorghum starch properties. J. Cereal Sci. 2001, 34, 261–268.
67. Dykes, L.; Rooney, L.W.; Waniska, R.D.; Rooney, W.L. Phenolic compounds and antioxidant activity of sorghum grains of varying genotypes. J. Agric. Food Chem. 2005, 53, 6813–6818.
68. Selle, P.H.; Ravindran, V.; Caldwell, R.A.; Bryden, W.L. Phytate and phytase: Consequences for protein utilisation. Nutr. Res. Rev. 2000, 13, 255–278.
69. Selle, P.H.; Cowieson, A.J.; Ravindran, V. Consequences of calcium interactions with phytate and phytase for poultry and pigs. Livest. Sci. 2009, 124, 126–141.
70. Doherty, C.; Faubion, J.M.; Rooney, L.W. Semiautomated determination of phytate in sorghum and sorghum products. Cereal Chem. 1982, 59, 373–378.
71. Selle, P.H.; Ravindran, V.; Cowieson, A.J.; Bedford, M.R. Phytate and Phytase; CABI Publishing: Oxon, UK, 2010; pp. 160–205.
72. Oatway, L.; Vasanthan, T.; Helm, J.H. Phytic acid. Food Rev. Int. 2001, 17, 419–431.
73. Ravindran, V.; Sands, J.S.; Hruby, M.; Partridge, G.G. Influence of Phyzyme tm xp Phytase on Nutrient Utilization in Broilers Fed Diets Containing Graded Levels of Phytate; Poultry Research Foundation: Sydney, Australia, 2005; pp. 238–241.
74. Wu, Y.B.; Ravindran, V.; Hendriks, W.H. Influence off exogenous enzyme supplementation on energy utilisation and nutrient digestibility off cereals for broilers. J. Sci. Food Agric. 2004, 84, 1817–1822.
75. Csonka, F.A.; Murphy, J.C.; Jones, D.B. The iso-electric points of various proteins. J. Am. Chem. Soc. 1926, 48, 763–768.
76. Kreydiyyeh, S.I.; Baydoun, E.A.H.; Churukian, Z.M. Tea extract inhibits intestinal-absorption of glucose and sodium in rats. Comp. Biochem. Physiol. C-Pharmacol. Toxicol. Endocrinol. 1994, 108, 359–365.
77. Kreydiyyeh, S.I. Inhibitors in tea of intestinal absorption of phenylalanine in rats. Comp. Biochem. Physiol. C-Pharmacol. Toxicol. Endocrinol. 1996, 113, 67–71.
78. Liu, N.; Ru, Y.J.; Li, F.D.; Cowieson, A.J. Effect of diet containing phytate and phytase on the activity and messenger ribonucleic acid expression of carbohydrase and transporter in chickens. J. Anim. Sci. 2008, 86, 3432–3439.
79. Weurding, R.E.; Enting, H.; Verstegen, M.W.A. The relation between starch digestion rate and amino acid level for broiler chickens. Poult. Sci. 2003, 82, 279–284.
80. Li, T.J.; Dai, Q.Z.; Yin, Y.L.; Zhang, J.; Huang, R.L.; Ruan, Z.; Deng, Z.; Xie, M. Dietary starch sources affect net portal appearance of amino acids and glucose in growing pigs. Animal 2008, 2, 723–729.
81. Enting, H.; Pos, J.; Weurding, R.E.; Veldman, A. Starch digestion rate affects broiler performance. Proc. Aust. Poult. Sci. Symp. 2005, 17, 17–20.
82. Weurding, R.E.; Veldman, A.; Veen, W.A.G.; van der Aar, P.J.; Verstegen, M.W.A. Starch digestion rate in the small intestine of broiler chickens differs among feedstuffs. J. Nutr. 2001, 131, 2329–2335.
83. Liu, S.Y.; Selle, P.H.; Cowieson, A.J. The kinetics of starch and nitrogen digestion regulate growth performance and nutrient utilisation in coarsely-ground, sorghum-based broiler diets. Anim. Prod. Sci. 2013, 53, 1033–1040.
84. Van den Borne, J.; Schrama, J.W.; Heetkamp, M.J.W.; Verstegen, M.W.A.; Gerrits, W.J.J. Synchronising the availability of amino acids and glucose increases protein retention in pigs. Animal 2007, 1, 666–674.
85. Axtell, J.D.; Kirleis, A.W.; Hassen, M.M.; Mason, N.D.; Mertz, E.T.; Munck, L. Digestibility of sorghum proteins. Proc. Natl. Acad. Sci. USA Biol. Sci. 1981, 78, 1333–1335.
86. Choi, S.J.; Woo, H.D.; Ko, S.H.; Moon, T.W. Confocal laser scanning microscopy to investigate the effect of cooking and sodium bisulfate on in vitro digestibility of waxy sorghum flour. Cereal Chem. 2008, 85, 65–69.
87. Batterman-Azcona, S.J.; Hamaker, B.R. Changes occurring in protein body structure and alpha-zein during cornflake processing. Cereal Chem. 1998, 75, 217–221.
88. Duodu, K.G.; Nunes, A.; Delgadillo, I.; Parker, M.L.; Mills, E.N.C.; Belton, P.S.; Taylor, J.R.N. Effect of grain structure and cooking on sorghum and maize in vitro protein digestibility. J. Cereal Sci. 2002, 35, 161–174.
89. Hamaker, B.R.; Kirleis, A.W.; Butler, L.G.; Axtell, J.D.; Mertz, E.T. Improving the in vitro protein digestibility of sorghum with reduing agents. Proc. Natl. Acad. Sci. USA 1987, 84, 626–628.
90. Rom, D.L.; Shull, J.M.; Chandrashekar, A.; Kirleis, A.W. Effects of cooking and treatment with sodium bisulfite on in vitro protein digestibility and microstructure of sorghum flour. Cereal Chem. 1992, 69, 178–181.
91. Selle, P.H.; Liu, S.Y.; Cai, J.; Caldwell, R.A.; Cowiesona, A.J. Preliminary assessment of including a reducing agent (sodium metabisulphite) in “all-sorghum” diets for broiler chickens. Anim. Feed Sci. Technol. 2013, 186, 81–90.
92. Selle, P.H.; Liu, S.Y.; Cai, J.; Caldwell, R.A.; Cowieson, A.J. Graded inclusions of sodium metabisulphite in sorghum-based diets: I. Reduction of disulphide cross-linkages in vitro and enhancement of energy utilisation and feed conversion efficiency in broiler chickens. Anim. Feed Sci. Technol. 2014, 190, 59–67.
93. Truong, H.H.; Cadogan, D.J.; Liu, S.Y.; Selle, P.H. Addition of sodium metabisulfite and microbial phytase, individually and in combination, to a sorghum-based diet for broiler chickens from 7 to 28 days post-hatch. Anim. Prod. Sci. 2015, doi:10.1071/AN14841.
94. Paterson, L.; Mitchell, J.R.; Hill, S.E.; Blanshard, J.M.V. Evidence for sulfite induced oxidative reductive depolymerisation of starch polysaccharides. Carbohydr. Res. 1996, 292, 143–151.
95. Paterson, L.A.; Hill, S.E.; Mitchell, J.R.; Blanshard, J.M.V. Sulphite and oxidative-reductive depolymerization reactions. Food Chem. 1997, 60, 143–147.
96. Ravindran, V.; Cabahug, S.; Ravindran, G.; Bryden, W.L. Influence of microbial phytase on apparent ileal amino acid digestibility of feedstuffs for broilers. Poult. Sci. 1999, 78, 699–706.
97. Liu, S.Y.; Cadogan, D.J.; Péron, A.; Truong, H.H.; Selle, P.H. A combination of xylanase, amylase and protease influences growth performance, nutrient utilisation, starch and protein digestive dynamics in broiler chickens offered maize-, sorghum- and wheat-based diets. Anim. Prod. Sci. 2014, doi:10.1071/AN14657.
98. Odjo, S.; Malumba, P.; Dossou, J.; Janas, S.; Bera, F. Influence of drying and hydrothermal treatment of corn on the denaturation of salt-soluble proteins and color parameters. J. Food Eng. 2012, 109, 561–570.
99. Gehring, C.K.; Bedford, M.R.; Cowieson, A.J.; Dozier, W.A. Effects of corn source on the relationship between in vitro assays and ileal nutrient digestibility. Poult. Sci. 2012, 91, 1908–1914.
100. Selle, P.H.; Liu, S.Y.; Cai, J.; Cowieson, A.J. Steam-pelleting and feed form of broiler diets based on three coarsely ground sorghums influences growth performance, nutrient utilisation, starch and nitrogen digestibility. Anim. Prod. Sci. 2012, 52, 842–852.
101. Zhang, G.Y.; Hamaker, B.R. Sorghum (Sorghum bicolor L. Moench) flour pasting properties influenced by free fatty acids and protein. Cereal Chem. 2005, 82, 534–540.
102. Liu, S.Y.; Selle, P.H.; Khoddami, A.; Roberts, T.H.; Cowieson, A.J. Graded inclusions of sodium metabisulphite in sorghum-based diets: II. Modification of starch pasting properties in vitro and beneficial impacts on starch digestion dynamics in broiler chickens. Anim. Feed Sci. Technol. 2014, 190, 68–78.
103. Selle, P.H.; Liu, S.Y.; Cai, J.; Cowieson, A.J. Steam-pelleting temperatures and grain variety of finely-ground, sorghum-based broiler diets. I. Influence on growth performance, relative gizzard weights, nutrient utilisation, starch and nitrogen digestibility. Anim. Prod. Sci. 2014, 54, 339–346.
104. Chiremba, C.; Taylor, J.R.N.; Rooney, L.W.; Beta, T. Phenolic acid content of sorghum and maize cultivars varying in hardness. Food Chem. 2012, 134, 81–88.
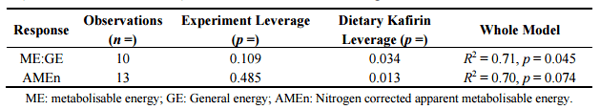
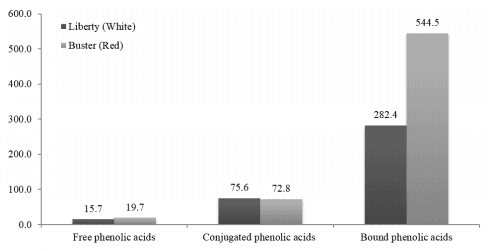
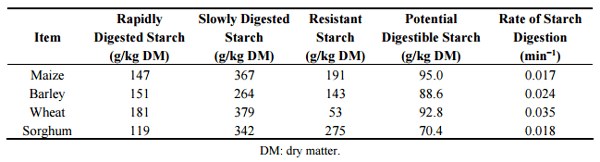
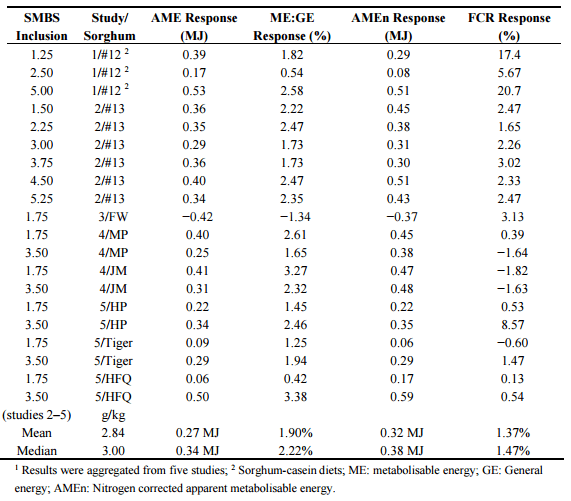
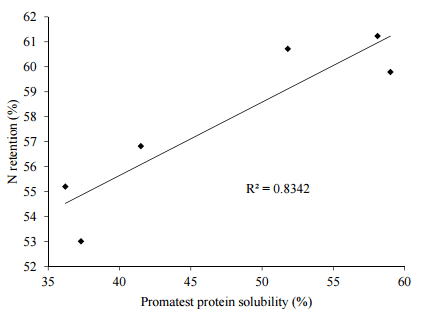
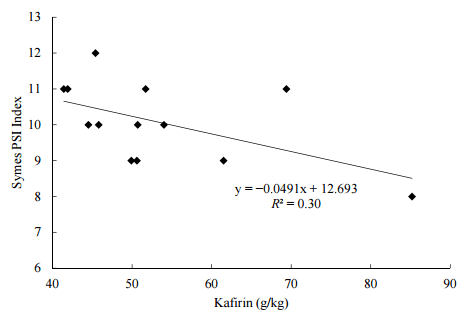










.jpg&w=3840&q=75)