INTRODUCTION
Inactivated vaccines are commonly used in poultry as part of a comprehensive vaccination protocol. These vaccines are capable of inducing high antibody titers, which can protect against systemic infections and are transferred from breeder hens to their progeny as maternal antibodies. In order to increase the immunogenicity, these vaccines contain adjuvants. The most common adjuvants in poultry vaccines are aluminum hydroxide gel (Alum) and oil-based emulsions (Schijns and Lavelle, 2011; Fox and Haensler, 2013).
Adjuvants increase immune responses, decreasing the amount of antigen required for vaccines Mount et al. (2013). This is achieved by 3 separate mechanisms (Waksman, 1979; Hunter, 2002). First, they may form a depot of antigens at the injection site, which allows a slow release, inducing a longer-lasting immune response. Second, they form particulate antigens, which are easily detected by antigen-presenting cells. Finally, adjuvants may cause a local inflammation that activates pattern-recognition receptors and stimulates the antigen presentation. Inflammation at the injection site is an adverse effect of adjuvants. This effect can be seen in food animals, including poultry, decreasing meat quality and increasing carcass condemnation (Singh and O’Hagan, 2003; Schat, 2014). In addition, adjuvants substantially increase the price of vaccines Meeusen et al. (2007). Therefore, finding new safe, stable and inexpensive adjuvants is important to improve current vaccines.
We propose that diatoms may act as safe, stable and inexpensive adjuvants. Diatoms are single-cell, eukaryotic microalgae encountered in all water habitats Dolatabadi and de la Guardia (2011). The surface of diatoms possesses a porous, nanostructured silica cell wall or frustule. Diatomaceous earth (DE) or diatomite is a natural siliceous sediment of diatoms. Due to the structural, mechanical, chemical, and optical features of diatoms and their silica cell wall, they have been proposed as drug-delivery carriers (Aw et al., 2013; Wang et al., 2013). In addition, silica-based cationic bilayers have been reported to have higher colloid stability than Alum and to act as an adjuvant Lincopan et al. (2009). Given the demonstrated use of nanostructured silica as an adjuvant, diatom silica may provide an inexpensive and abundant source to use in vaccines. In addition, diatoms can be used as a vaccine vector where the silica-based cell wall can act as an insertion surface for immunogenic proteins (Corbeil et al., 2015). Currently, no information regarding the use of diatoms or DE as adjuvants has been published, to the authors’ knowledge.
In this study, we tested the toxicity of diatoms in embryonated eggs. Subsequently, we used diatomaceous earth (DE) as an adjuvant in the preparation of inactivated Newcastle disease virus (INDV) vaccines in chickens. The objective was to determine if diatoms or DE had any detrimental effect on embryos and/or hatched chicks and their hatchability, quality and body weight after in-ovo administration in an INDV vaccine. In addition, we assessed the humoral immune response against NDV after 1 in-ovo vaccination followed by 2 subcutaneous boosters.
MATERIALS AND METHODS
Welfare Statement
All animal trials were approved by the Institutional Animal Care and Use Committee (IACUC) at the University of California, Davis (protocol #: 177863). All experiments were conducted in accordance with the ILAR Guide for the Care and Use of Laboratory Animals and the UC Davis Animal Welfare Assurance on file with the US Public Health Service.
Diatom Cultivation
Axenic 1.5 L cultures of Thalassiosira pseudonana (CCMP1335) were grown in artificial seawater medium (ESAW, http://www3.botany.ubc.ca/cccm/ NEPCC/esaw.html) at 18°C under continuous light. Cultures were stirred and bubbled with air until a concentration of approximately 1 × 106 cells • mL−1 was achieved. Cells were harvested by filtration on 3 μm polycarbonate filters, resuspended in 1× PBS to remove excess salts and pelleted by centrifugation for 10 min at 3,100 × g. Cell pellets were lyophilized and stored at 4°C.
Experiment I: Effect of In-ovo Application of Diatoms
Specific pathogen-free (SPF) eggs (Sunrise Farms, Catskill, NY) were incubated. At d 18 of embryonation, 150 fertile eggs were divided randomly into 5 groups of 30 eggs each. Group 1, 2, and 3 were inoculated in-ovo with 20, 100, and 500 μg of lyophilized diatoms in a volume of 200 μL of phosphate buffered saline (PBS), respectively. Group 4 was inoculated with 200 μL of Freund’s incomplete adjuvant (FIA) (MP Biomedicals LLC., Solon, OH) and group 5 was inoculated with 200 μL of PBS. The suspensions were delivered in-ovo through the air chamber at 1-inch depth using a 1.5- inch, 21-gauge needle. Hatchability and chick quality were recorded. For the quality assessment, the navel was evaluated and the percentage of chickens with a crusty navel scar was calculated. After hatch, chicks were maintained in BSL2 units at the Teaching and Research Animal Care Services of University of California Davis, where they had free access to feed and water. Chickens were weighed and uniformity was calculated at 1, 3, 5, 7, 11, and 14 d of age.
Experiment II: DE Adjuvant Capabilities
Vaccine Preparation. A live attenuated NDV obtained from a commercial LaSota strain vaccine (Merial Select, Gainesville, GA) was used in the formulation of experimental inactivated vaccines. The virus was reconstituted in PBS and titrated in 10-day-old SPF embryonated eggs via the allantoic sac according to standard procedures Dufou-Zavala (2008). The vaccine was diluted to a titer of 2 × 106 EID50/mL. Five hundred milliliters were inactivated by adding formalin (Fisher Scientific, Fairlawn, NJ) to a concentration of 0.08% and stirring for 6 h at 25°C. The virus was tested for complete inactivation by inoculating 100 μL into the allantoic cavity of embryonated SPF eggs as described for viral titration. Two d post inoculation, allantoic fluid from these eggs was harvested and tested for its hemagglutination (HA) activity. The absence of HA activity confirmed viral inactivation. The inactivated viral stock was stored at 4°C until used.
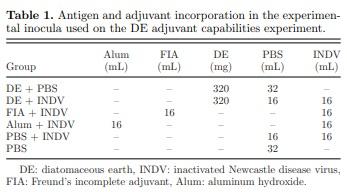
Six experimental inocula were prepared for this study: 320 mg DE (Earthworks Health, Norfolk, NE) in 32 mL PBS (DE+PBS); 320 mg DE in 16 mL PBS + 16 mL of inactivated NDV in tryptose broth (DE+INDV); 16 mL FIA + 16 mL inactivated NDV in tryptose broth (FIA+INDV); 16 mL of 2% aluminum hydroxide gel adjuvant (InvivoGen, San Diego, CA) + 16 mL inactivated NDV in tryptose broth (Alum+INDV); 16 mL PBS + 16 mL inactivated NDV in tryptose broth (PBS+INDV); and 32 mL PBS (PBS) (Table 1). The vaccine containing FIA (FIA+INDV) was prepared using a homogenizer (IKA, Staufen, Germany). The other experimental vaccines were prepared by stirring for 30 minutes at room temperature. All vaccines were stored at 4°C until used.
Vaccine Administration. At 18 d of embryonation, 192 viable SPF eggs were randomly divided into 6 groups of 32 eggs each. Eggs from each group were inoculated in-ovo with 100 μL of one of the previously
described experimental inocula. After vaccine administration, each group was placed in a separate basket and transferred to the hatcher. At 21 and 29 d of age, chicks from each group were inoculated with 200 μL of the respective experimental inocula via subcutaneous route in the base of the neck.
Hatchability, Chick Quality, and Body Weight.
Chicks were placed in BSL2 units at the Teaching and Research Animal Care Services at the University of California, Davis. Hatchability for each group and individual chick quality assessments including navel condition, chick length, and hock condition were recorded. The navel condition was assessed using a scoring system: 1 (clean and closed navel), 2 (presence of a string, discolored button or open navel <2 mm), or 3 (black button or open navel >2 mm) Molenaar et al. (2010). The chick length was measured by stretching the chick along a graduated ruler and recording the chick length from the tip of the beak to the terminal middle toe, excluding the nail Hill (2001). The hock condition was evaluated based on the presence or absence of red discoloration in the hock. Additionally, live body weight was determined weekly for 3 wk. Morbidity and mortality were recorded throughout the experimental period.
Residual Yolk Measurement and Sample Collection.
Five chickens per group were euthanized at 1, 7, 14, and 29 d of age for quality assessment (residual yolk) and tissue collection. At 1 day of age, the percentage of residual yolk from live body weight was calculated. In brief, the yolk sac was collected, weighed and compared to the body weight in grams of the chicks (Van der Pol et al., 2013; Maatjens et al., 2014).
In order to detect the deposition of DE in chicken organs or tissues, tracheas, esophagus, duodenum, skin and bursa of Fabricius were collected at all necropsies and submitted to the transmission electron microscopy (TEM) facility at the California Animal Health and Food Safety (CAHFS) Laboratory. Formalin fixed specimens were post-fixed in 2.5% glutaraldehyde followed by osmium tetroxide before embedding and sectioning according to standard methods Bozzola and Russel (1991).
Blood was collected from all birds via ulnar vein puncture at 7, 14, 21, 29, 36, 39 and 44 d of age. Sera were harvested and stored at −20°C until tested. NDV-specific antibodies were assessed using a commercial NDV enzyme-linked immunosorbent assay (ELISA) (IDEXX, Westbrook, ME) according to the manufacturer’s protocol. Each sample was tested in duplicate. Absorbance at a wavelength of 650 nm was measured using a spectrophotometer (BioTek, Winoosk, VA).
Statistical Analysis
Data was statistically analyzed using Prism Graphpad 5 (San Diego, CA). One-way ANOVA and Student’s t test were used to compare mean values between groups at each time point. Differences were considered significant if P < 0.05. Differences in hatchability and incidence of umbilical scars were analyzed for statistical differences using Chi-square test. Significance was determined at P < 0.05.
RESULTS
Experiment I: Effect of In-ovo Application of Diatoms.
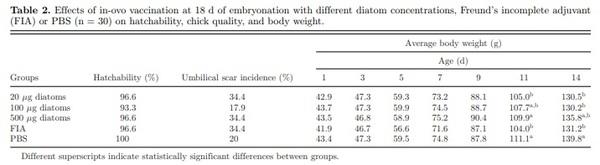
No embryonic mortality attributed to technique was detected following in-ovo inoculation on d 18 of embryonation. Hatchability ranged from 93.3% in the group that received 100 μg of diatoms to 100% in the group that received only PBS (Table 2). Groups that received FIA, 20, and 500 μg of diatoms (34.4%) had the highest percentage of birds with umbilical scars, while birds that received 100 μg diatoms showed the lowest level with 17.9%. No differences in hatchability, uniformities, and umbilical scars were detected between experimental groups (Table 2). No significant differences in live body weight were detected between groups from hatch until 9 d of age. At 11 d of age, higher body weights (P < 0.05) were detected in groups inoculated with 500 μg diatoms and PBS compared with groups inoculated with FIA and 20 μg of diatoms. At 14 d of age, the PBS group exhibited a significantly higher body weight (P < 0.001) followed by the group inoculated with 500 μg of diatoms (Table 2).
Experiment II: DE Adjuvant Capabilities
Hatchability, Chick Quality, and Body Weight. No embryonic mortality was detected following in-ovo vaccination. No statistically significant differences in hatchability were detected between experimental groups (Table 3). The highest hatchability was for the eggs that received DE+PBS and Alum+INDV followed by the groups that received DE+INDV and PBS. The lowest hatchability was for the group PBS+INDV.
Minor variations were detected in residual yolk while chick length was very uniform between groups (Table 3). Additionally, DE+INDV group had the healthiest navels with a score of 1.5 followed by Alum+INDV. Live body weight varied slightly between groups (Table 3). No differences in residual yolk, chick
length, and chick weight were detected between experimental groups. Red hocks were not seen. Finally, there was no morbidity or mortality during the course of the experiment.
Detection of DE by Electron Microscopy. We were not able to detect DE in any of tested tissues or organs submitted to electron microscopy.
NDV Antibody Titers After Vaccination. After in-ovo vaccination, no NDV specific antibody titers were detected at 7, 14, 21, and 29 d of age. Figure 1 displays the NDV antibody titers measured at 36, 39, and 44 d of age after 2 boosters performed at d 21 and 29 of age. None of the chickens from the groups inoculated with DE+PBS, PBS+INDV, or PBS alone showed detectable levels of NDV specific antibodies measured by ELISA at any time point (Figure 1A, B, and C).
Seven d after the second booster at 36 d of age, 6 out of 12 chickens in group DE+INDV (geometric mean titer (GMT) 437.54), 4 out of 10 chickens in group FIA+INDV (GMT 602.54), and 5 out of 12 chickens in group Alum+INDV (GMT 472.22) showed positive antibody titers. No differences were detected in mean titers between these 3 groups or with the other experimental groups, even though GMTs were numerically higher (Figure 1A).
At 10 d post second booster, or 39 d of age, a similar trend was observed in the groups that received adjuvanted inocula containing INDV (Figure 1B). The number of positive birds were 6 out of 12 in group DE+INDV, 5 out of 10 in group FIA+INDV and 7 out of 12 in group Alum+INDV. Birds of the FIA+INDV group displayed higher titers (GMT 1294.34) than the birds of groups DE+INDV (GMT 786.55) and Alum+INDV (GMT 491.41). No differences between these 3 groups were detected, but the titers were higher (P < 0.05) than in the groups that had not received INDV or received INDV alone.
At 15 d post second booster, 44 d of age, group FIA+INDV (8/10 positives, GMT 3475) showed higher titers (P< 0.05) (Figure 1C) than groups DE+INDV (7/12 positives, GMT 1025) and Alum+INDV (8/12 positives, GMT 1072).
DISCUSSION
In-ovo vaccination is a common practice in the United States. These vaccines are often applied during transferring eggs from the incubator to the hatcher at 18 or 19 d of incubation to prevent Marek’s Disease (MD) and infectious bursal disease virus in broiler chickens (Gagic et al., 1999; Williams and Zedek, 2010). Inovo vaccination against NDV has given varying results when live vaccines were utilized (Dilaveris et al., 2007; Kapczynski et al., 2012; Ramp et al., 2012). However, in-ovo vaccination with INDV vaccines Stone et al. (1997) or recombinant vaccines expressing NDV antigens Palya et al. (2012) have shown more success. In the present study, the safety of diatoms or DE for vaccine
adjuvanticity was investigated and INDV experimental vaccines were crafted and inoculated into 18-d-old embryonated SPF eggs.
The first objective was to determine if there were any detrimental effects of diatoms or DE when used as a vaccine adjuvant. Results showed no detrimental effects of in-ovo vaccination of diatoms and DE specifically on hatchability, chick quality, and live body weight during the first 2 to 3 wk of age of the chicks (Tables 2 and 3). This is an important finding since commonly used vaccine adjuvants can cause adverse effects Petrovsky and Aguilar (2004). In food animals, a reduction of the meat quality and increased condemnation at the processing plants associated with the use of these products has been reported (Singh and O’Hagan, 2003; Schat, 2014). These experiments tested as much as 500 μg of diatoms and 1 mg of DE per dose. Considering the fact that DE and diatoms did not cause adverse effects such as decrease in hatchability of embryonated eggs or chick quality and body weight, further studies could focus on the dose response effect of diatoms or DE in the generation of a protective immune response even at doses over 1 mg.
The second objective was to test the capability of a DE adjuvanted INDV vaccine to elicit antibody responses after in-ovo and subcutaneous chicken vaccination. The importance of humoral immunity, specifically immunoglobulin G (IgG) generation after NDV vaccination, is essential to protect flocks against Newcastle disease virus exposure (Kapczynski and King, 2005). This protection is induced by neutralizing antibodies specific for the fusion and hemagglutinin glycoproteins, which are critical for viral attachment to host cells and infection (Boursnell et al., 1990a; Boursnell et al., 1990b; Reynolds and Maraqa, 2000). With weekly sera after in-ovo vaccination alone, none of the experimental vaccinated groups produced any detectable NDV-specific antibodies up to d 29 of age. The reason might be that the NDV titer before inactivation, 105 EID50, was not sufficient to generate a measurable humoral immune response when inoculated in-ovo. Stone et al. (1997) reported that serum antibody titers against NDV were detected 14 d post hatch after in-ovo vaccination with 109.9 EID50 of INDV and the antibody titer increased as the antigen concentration increased. In addition, Toro et al. (2012) demonstrated that a titer of 106 EID50 in a replication deficient adenovirus construct encoding for a S1 gene segment of infectious bronchitis virus vaccine, is not enough to generate protective responses in chickens vaccinated using the in-ovo route. Another potential reason for lack of detectable antibodies could be the egg compartment in which the vaccine was delivered. Others showed that in-ovo vaccination against Marek’s disease is more than 90% effective when vaccines were delivered into the amnion cavity or into the embryo; however, poor protection resulted when the vaccine was delivered into the allantoic cavity (Wakenell et al., 2002). In this study, we did not monitor into which compartment the vaccine was delivered. However, Breedlove et al. (2011) investigated embryo tissues targeted by a replication competent adenovirusfree vector expressing an avian influenza H5 gene after 18-d embryonated egg inoculation using the same technique as in our experiments. Imaging showed that the vaccine was in-ovo inoculated in the amnion cavity, allantoic cavity, or both. The vector’s DNA was detected in the chorioallantoic membrane as well as in the embryonic bursa of Fabricius, esophagus, and thymus suggesting possible swallowing of the inoculum placed in the amnion. In addition, vaccine localization in the allantoic cavity might have allowed the inoculum exposure to the bursa of Fabricius (Breedlove et al., 2011). Thus our methods should have reached adequate targets. In contrast, Stone et al. (1997) reported that the inoculation of an inactivated oil-emulsion NDV and avian influenza vaccine in-ovo 1.5-inch depth induced adequate antibody titers.
In the groups that were vaccinated twice subcutaneously after hatching, chicken groups that received vaccines adjuvanted with FIA, Alum, or DE showed positive antibody titers and had increasing titers until the end of the study (Figure 1). However, some individual chicks in the 3 groups still showed no antibody responses against NDV. This can be explained by the low NDV titer we used in the experimental vaccine preparation in order to test the ability of adjuvants to increase immune responses. Miller reported that NDVspecific antibodies are induced more consistently when chickens are inoculated with INDV viral titers of 109.5 EID50/dose Miller et al. (2013). As expected, birds vaccinated with non-adjuvanted INDV did not generate detectable antibodies during the entire study. This situation allowed us to see the full potential of the tested adjuvants. Consequently, we were able to demonstrate that DE served as an adjuvant in the experimentally tested INDV vaccine. Interestingly the level of adjuvanticity demonstrated by DE 15 d post second booster (GMT: 1025 and 7/12 positive chickens) was similar to the adjuvant capabilities of Alum (GMT: 1072 and 8/12 positive chickens). Alum, a commonly used adjuvant in human vaccination, is not commonly used in livestock due to its elevated price. This makes other adjuvants such as DE an attractive potential alternative due to its low price and availability.
In conclusion, our experimental results suggest that DE could serve as a potential adjuvant for vaccines against poultry diseases. We demonstrated that neither diatoms nor DE had detrimental effects on hatchability, chick quality, and live body weight when used as a vaccine adjuvant. Even though we did not see a clear immune response when vaccines were applied in-ovo, subcutaneous boosters with INDV adjuvanted with DE produced NDV specific antibodies starting at 7 d post second booster in chickens. In addition, the efficacy of DE as an adjuvant for INDV vaccines was comparable to the activity demonstrated by Alum. From an applied perspective we can propose the use of DE as an adjuvant in poultry inactivated vaccines. However, more research is needed to test the effect on the elicited immune response using different antigen, DE concentrations, and distinct vaccine administration routes.
ACKNOWLEDGMENTS
We wish to acknowledge Alejandra Figueroa, Ana Paula Da Silva, Theodore Derksen and Lauren Crum for outstanding help and technical assistance. This work was supported by the School of Veterinary Medicine, University of California, Davis and by USDA, NIFA grant # 2012-67015-30197 at UCSD.
This article was originally published in 2017 Poultry Science 96:288–294. http://dx.doi.org/10.3382/ps/pew250. This work is written by (a) US Government employee(s) and is in the public domain in the US.









.jpg&w=3840&q=75)