INTRODUCTION
Variability in the nutritional value of feed ingredients for poultry (and indeed other livestock species) has been widely reported over several decades and has been the impetus behind a sustained body of research work. For example, Sibbald and Slinger (1962), Mollah et al. (1983), and Leeson et al. (1993) reported variation in the apparent metabolizable energy (AME) content of corn (3,476–3,960 kcal/kg dry matter [DM]), wheat (2,948–3,960 kcal/kg DM), and soybean meal (SBM; 2,520–2,960 kcal/kg DM) when fed to broiler chickens. Leeson et al. (1993) also noted significant variation in the digestibility coefficients of amino acids (AA) in corn, for example, with the apparent digestibility coefficient of lysine varying from 0.28 to 1.08. Douglas et al. (2000) reported variation in the ileal digestible energy concentration of corn/SBM-based diets associated with 12 different SBM sources from 2,848 to 3,104 kcal/kg DM. Ravindran et al. (2007) reported an apparent ileal AA digestibility coefficient of 0.81 in maize (8 samples) with a standard deviation of 0.03, of 0.83 in wheat (8 samples) with a standard deviation of 0.02, and of 0.82 in SBM (10 samples) with a standard deviation of 0.01. However, although variation at the ingredient level is well reported and even occasionally understood, the contribution of the animal to this variability is much less clear.
Hughes and Choct (1997) raised the concept of the “low AME chicken” and asked the question whether individuality of digestive physiology in birds may contribute to the variance in the AME of wheat with different arabinoxylan concentrations. One wheat-based diet was fed to 40 individually housed broilers, and a range of 2,103–3,561 kcal/kg DM in AME was observed, around a mean of 2,892 kcal/kg DM. The authors then subdivided the birds into ’low-, medium-, or high’-AME groups and observed that the low-AME group had higher excreta energy concentration, higher feed conversion ratio, and lower weight gain than the high-AME group, which was clear evidence of significant heterogeneity of digestive capacity and efficiency within a population of broilers fed with the same diet.
Thus, it is apparent that variance exists in the nutritional value (AME and AA) within classes of feed ingredients such as corn, SBM, or wheat. It is further apparent that individuality of bird digestive physiology contributes to this variance, at least for AME and starch digestibility, and that this may be particularly relevant when ingredient quality is inherently low. However, the contribution of individual animal heterogeneity to AA digestibility is not clear and has not received much attention in the literature. Therefore, the purpose of the study presented herein was to explore variance in AA digestibility coefficients for two distinct batches of SBM fed without or with an exogenous protease on an individual bird basis. The hypothesis was that SBM with an inherently low digestibility of AA would generate higher variability at an individual animal level than SBM with an inherently high digestibility and that exogenous protease may reduce this variance, perhaps particularly by augmenting digestibility in birds with compromised intestinal capacity.
MATERIALS AND METHODS
The experiment was approved by the Animal Ethics Committee of the University of New England (approval no.: AEC18-068). Health and animal husbandry practices complied with the Code of Practice for the Use of Animals for Scientific Purposes issued by the Australian Bureau of Animal Health (National Health and Medical Research Council, 2014). The experiment was conducted as per the method used by Ravindran et al. (2017), with some minor modifications (detailed in the following section).
Housing, Birds, and Diets
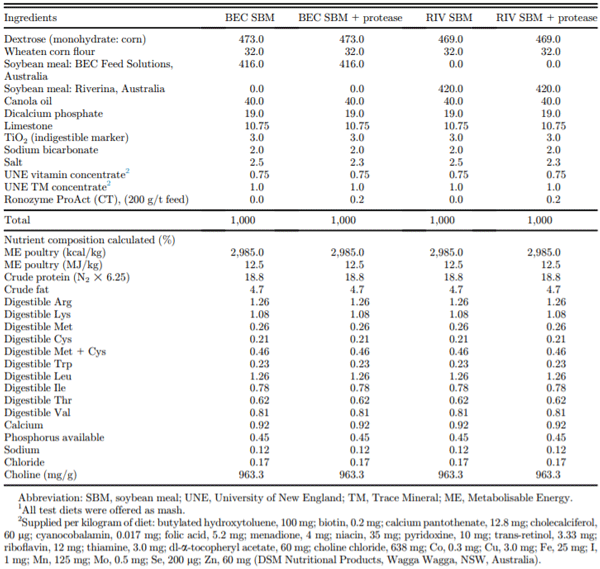
Ross 308-day-old chicks (initial weight, 41.8 ± 0.01 g/ bird) were obtained from Aviagen Hatchery Pty Ltd. (Goulburn, NSW, Australia). The chicks were reared in a climate-controlled room for 29 D on standard wheat-based starter (days 1–15; AME: 3,050 kcal/kg, CP: 22.5%) and grower (days 15–33; AME: 3,100 kcal/ kg, CP: 21.0%) diets that were formulated as per breeder guidelines. The temperature at day 1 was 33 °C, and this was reduced in a stepwise manner to 21 °C on day 25. On day 30, the birds were randomly distributed to 72 individual cages (1 bird per cage) with the commercial grower diet being fed for a further 3 D to permit acclimatization within the new battery cage environment. The 4 assay diets (Table 1) were introduced at the age of 34 D and continued until 37 D. These diets were semisynthetic and comprised 2 alternative SBM sources (BEC; BEC Feed Solutions, Australia, or RIV; Riverina, Australia), fed without or with a monocomponent exogenous protease in a 2*2 factorial arrangement of treatments. The SBM batches used herein were commercially available in Australia at the time the research was conducted and were not specifically selected based on any particular chemical or mechanical properties. Water was available ad libitum throughout the trial period. The experiment was concluded on day 37, at which point the ileal collection was carried out. The exogenous protease used was RONOZYME ProAct (DSM Nutritional Products, Kaiseraugst, Switzerland), a serine protease expressed in Bacillus licheniformis. One protease (PROT) unit is defined as the amount of enzyme that releases 1 mmol of p-nitroaniline from 1 mmol substrate (Suc-Ala-Ala-Pro-Phe-pNA per min at pH 9.0 and 37 °C). The enzyme was added to provide 15,000 PROT/kg of feed. Chemical characteristics of the SBM (solubility in KOH, urease activity, and trypsin inhibitor activity) were determined by Eurofins (Eurofins Agro Testing Denmark A/S, Vejen, Denmark; AOAC, 2000).
Monitoring
The birds were inspected twice daily, in the morning and late afternoon, particularly to check the general health status, feed, and water supply as well as room temperature and ventilation.
Ileal Digesta Collection, Drying, and Storage
On day 37, the birds’ live weight was recorded, and the birds were then euthanized by carbon dioxide asphyxiation. The ileum was excised, and ileal digesta were collected from each bird in a labeled plastic container, which was placed in ice and frozen (−20 °C) immediately. The sample was collected from the distal half of the ileum by flushing with distilled water. The ileum was defined as that portion of the small intestine extending from Meckel’s diverticulum to a point of 40 mm proximal to the ileocaecal junction. After freezing, the ileal samples were freeze-dried, ground (0.5 mm), and stored in plastic containers at room temperature for titanium (Ti) and AA analysis.
Determination of Ti Concentration
The concentration of Ti in the ileal digesta and the diets was determined using the method described by Short et al. (1996). In brief, a 100-mg sample of freeze-dried ileal contents and 200-mg samples of diets were weighed in porcelain crucibles in duplicate and ashed for 13 h at 580 °C. Then, 5 ml of 7.4 mol sulfuric acid was added after cooling, and the mixture was boiled on a heating plate for 30 min at 200 °C; the temperature was then raised to 250 °C for another 30 min until the sample was completely dissolved. The mixture was quantitatively transferred into a 50-ml volumetric flask after filtration through a filter paper. Subsequently, 10 ml of hydrogen peroxide (30%, v/v) was added to each flask, and the contents were further diluted up to 50 ml with distilled water and thoroughly mixed. Absorbance was read using a UV spectrophotometer at 410 nm. The Ti concentration was determined from the standard curve and converted to milligram per gram of the sample.
Table 1. Composition of (g/kg, as-fed basis) of assay diets1 (days 34–37) for the measurement of apparent amino acid digestibility of soybean meals.
Amino Acid Profile Analysis
Sample Preparation Approximately 180 mg of the sample was hydrolyzed in 20% HCl for 24 h at 110 °C. An internal standard (DL-Norvaline and α-aminobutyric acid) was added to each sample after hydrolysis. After a 1:25 dilution in water, 10 μL of the solution was derivatized using an AccQ Tag Ultra Derivatization Kit (Waters, NSW, Australia). HPLC analysis was based on the method used by Cohen (2001) but was adapted for use with an ACQUITY Ultra Performance LC (Waters Corp., Rydalmere, NSW, Australia) system. The column used was an ACQUITY Ultra Performance LC BEH C18 1.7-μm column (Waters, NSW, Australia) with detection at 260 nm and a flow rate of 0.7 mL/min. (AOAC, 2000).
After first performing acid hydrolysis, the concentrations of AA were determined by precolumn derivatization AA analysis using 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate, followed by separation of the derivatives and quantification by reversed-phase HPLC (Cohen and Michaud, 1993; Cohen, 2001). Amino acids were detected by UV absorbance.
Dry Matter Digestibility and Apparent Ileal Digestibility Calculation
The DM digestibility was calculated based on the change in concentration of the Ti marker in DM of feed vs. DM of ileal digesta using the following formula:
DM digestibility = [(Ti (g=kg DM) digesta) -
(Ti (g=kg DM) feed) / (Ti (g=kg DM) digesta).
where Ti g/kg DM digesta is the concentration of the Ti marker in ileal digesta and Ti g/kg DM feed is the concentration of the Ti marker in feed.
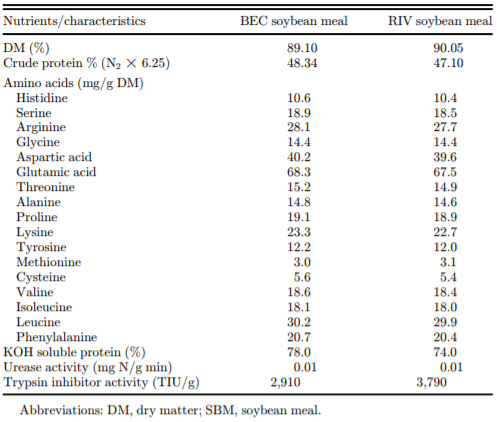
Table 2. Amino acid composition, solubility in potassium hydroxide (KOH), protein solubility (%), and urease activity (mg N/g/min) analyzed in the two SBM samples.
The apparent ileal digestibility (AID) coefficients were calculated from the dietary ratio of AA to the Ti marker relative to the corresponding ratio in the ileal digesta by the following formula:
AID coefficient = 1-[(AA/Ti) I / (AA/Ti) D]
where (AA/Ti) D is the ratio of AA to Ti in the diet and (AA/Ti) I is the ratio of AA to Ti in ileal digesta.
Statistical Analysis
Data were subjected to 2-way ANOVA analysis using JMP version 12.2.0 (SAS Institute, Cary, NC), with the model terms being SBM and protease. Significance was set at P < 0.05, with trends being noted at P < 0.1. As the focus of this work was on the contribution of individual animals to AA digestibility of SBM, coefficient of variation (CV) of AA digestibility within treatments was also assessed and reported. No outliers were detected, and all the results are based on 18 individual animal observations per treatment.
RESULTS
The chemical characteristics of the two SBM batches used in the present study are presented in Table 2. No obvious differences existed between the two SBM batches in terms of the AA profile although the BEC SBM had a marginally higher protein concentration (48.3% vs. 47.1%, respectively) and a commensurately higher AA concentration (2.33% vs. 2.27% lysine, respectively). Solubility in KOH and urease activity were similar between the two SBM batches. The BEC SBM had a lower trypsin inhibitor concentration (2,910 vs. 3,790 TIU/g, respectively) than the RIV SBM.
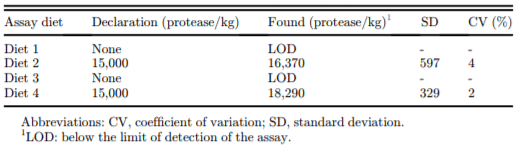
Recovery of protease activity in the experimental diets is presented in Table 3. No protease activity was detected in the two control diets, whereas protease activity from approximately 16,000 to 18,000 PROT/kg was detected in the diets in which protease had been added. These values compared well with the expectations (15,000 PROT/kg) and are within normal ranges for this product.
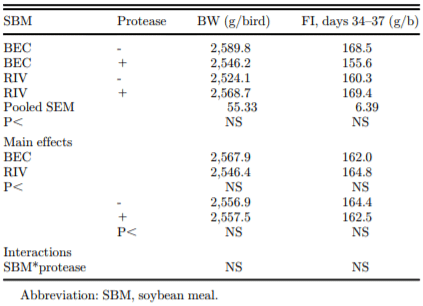
To set the context for the digestibility results, the body weight of the birds (at the termination of the experiment on day 37) and the feed intake of the experimental diets (days 34–37) are presented in Table 4. There were no significant treatment effects on body weight or on feed intake. It should be noted that all birds received a common starter (days 1–15) and grower (days 15–33) diet and that the experimental diets were only offered, for the purpose of assessment of ileal AA digestibility, from day 34 to 37. However, these performance characteristics are important and show that feed intake during the experimental phase was not responsible for the digestibility responses that were observed. Feed intake at an individual bird level was not correlated (P > 0.05; data not shown) with AA digestibility.
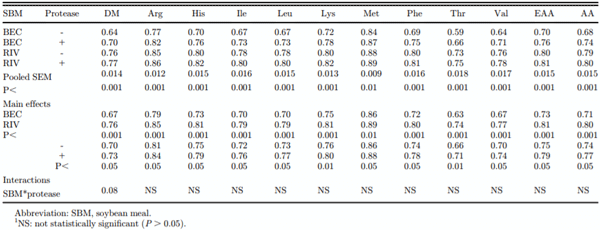
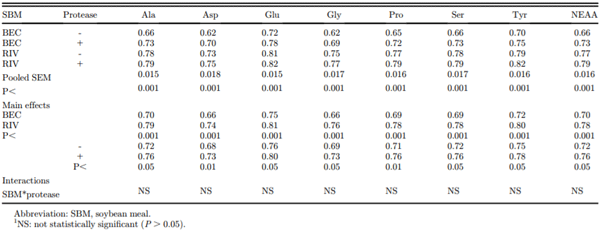
The effect of SBM origin and protease addition on the AID of nonessential AA is presented in Table 5. There were no interactions between SBM and protease for any AA. The BEC SBM had lower AID coefficients for all AA (for all nonessential AA, the mean values = 0.70 vs. 0.78; P < 0.001). Addition of exogenous protease increased AA digestibility coefficients from an average of 0.72 to 0.76 (+5.5%; P < 0.05).
Table 3. Protease recovery of assay diets (PROT/kg).
The effect of SBM origin and protease addition on the AID of essential AA and DM is presented in Table 6. There were no interactions between SBM and protease for any of the AA, although a tendency for an interaction was observed for DM digestibility as protease effect was more evident for the BEC SBM than for the RIV SBM (interaction P = 0.08). The BEC SBM had a lower AID coefficient for essential AA than the RIV SBM (0.73 vs. 0.81, respectively; P < 0.001). Addition of exogenous protease resulted in an increase in the digestibility coefficient for all essential AA, on average from 0.75 to 0.79 (+5.3%; P < 0.05) although this ranged from +2.3% for Met to +7.5% for Thr.
Although there were no significant interactions between the SBM batch and protease addition, the BEC SBM (with an inherently lower AA digestibility) appeared more responsive to protease addition. In fact, as presented in Figure 1, there was a significant second-order polynomial correlation between the inherent digestibility of AA in the two SBM batches and the percentage effect of protease on the same.
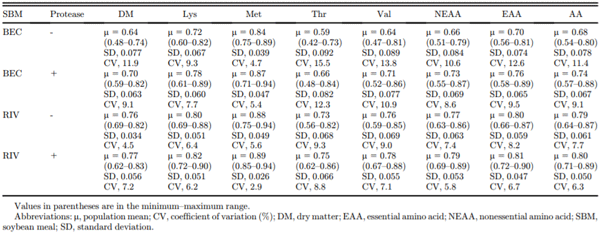
Finally, a major purpose of the present study was to explore the contribution of individual broilers to variance in the digestibility of AA in SBM. Variance analysis is presented in Table 7. In addition to the BEC SBM having a lower inherent AA digestibility, variability at the individual bird level was also higher. For example, the standard deviation of AA digestibility for the BEC SBM (without added protease) was 0.078, based on a population mean of 0.68 (CV of 11.4%), whereas the same values for the RIV SBM were 0.061, on a mean of 0.79 (CV of 7.7%). Variation in AA digestibility at the individual bird level was lower when exogenous protease was added compared with the SBM fed without protease. For example, CV for AA digestibility in the BEC SBM was reduced from 11.4 to 9.1% and in the RIV SBM was reduced from 7.7 to 6.3% by the addition of exogenous protease.
Table 4. Effect of soybean origin and protease addition on the body weight (BW, g/b) of birds on day 37 (immediately before ileal digesta collection) and feed intake (FI, g/b) per treatment during the digestibility assay (days 34–37).
DISCUSSION
The fact that there were no differences in feed intake during the experimental phase is important as feed intake (and digestible protein intake) is known to influence digestibility values (Fan et al., 1994; Angkanaporn et al., 1997). When low-protein diets are fed and/or low protein intake results from low feed intake per se, it is common for apparent ileal AA digestibility values to decrease as endogenous AA loss makes up a larger proportion of the AA flow in the intestine (Adeola et al., 2016; Ravindran, 2016). The fact that feed intake was not influenced by treatment in the present experiment gives confidence that differences associated with protease addition and/or SBM origin are genuine and not artifacts of the assay conditions.
The AID of AA in the BEC SBM was lower than in the RIV SBM. The reason for this difference is not clear and is apparently unrelated to solubility in KOH, trypsin inhibitor concentration, or urease activity. Solubility in KOH and urease activity in SBM are known to influence nutritional value, but most reports on this topic involve ranges that are substantially higher than those observed herein. For example, Heger et al. (2016) noted a significant impairment of broiler performance when diets (based on full-fat SBM) were offered with a range of urease activity from 0.01 to 4.15 mg N/g and KOH solubility from 80.2 to 94.2. Importantly, Parsons et al. (1991) noted that broiler performance was not influenced by solubility of SBM in KOH when solubility values were higher than 59% but that values lower than this threshold resulted in an impaired gain-to-feed ratio. In the present work, both SBM batches had solubility values in excess of 70%, and this was unrelated to differences in ileal AA digestibility, which is in agreement with the observations of Parsons et al. (1991). Generally, solubility values (in KOH) between 78 and 84% are optimal and would be suggestive of adequate processing conditions, that is, neither underprocessed nor overprocessed (Dozier and Hess, 2011). Thus, the two batches of SBM in the present work appear to be well processed, and the general solubility is not a critical factor in explaining the difference in nutritional value. The higher trypsin inhibitor concentration in the RIV SBM was not associated with reduced AA digestibility. It is possible that birds compensated for the higher inhibitor concentration by increasing pancreatic enzyme production. This has previously been noted across a wider range of trypsin inhibitor concentration than was the case herein (Clarke and Wiseman, 2005).
Table 5. Effect of soybean origin and protease addition on the apparent ileal digestibility of nonessential amino acids (NEAA) in broilers (n = 18).1
A recent work (Cowieson et al., 2016) used proteomics to explore the undigested peptides from the ileum of broilers fed with corn/SBM- or corn/SBM/raw SBM– based diets. In this work, it was observed that a substantial proportion of the protein in the ileum of broilers was from undigested SBM storage proteins (glycinin and beta-conglycinin) and endogenous proteins such as enzymes and mucin. It is possible that the differences between the two SBM batches in the present experiment are associated with differences in the concentration of these recalcitrant proteins and/or changes to specific endogenous loss associated with antigenic proteins, particle size, or, for example, phytate concentration. Indeed, on an individual AA level, there is a strong suggestion that the BEC SBM may have lower apparent digestibility owing to higher endogenous AA flow. Instructively, the difference between the two SBM batches in AID of Met (4.5%), Arg (9.4%), and Lys (10%) is considerably less than that for AA that are more associated with endogenous proteins such as Gly (17.3%) and Thr (19.2%). Similar patterns have been found to be associated with the effect of dietary antinutrients on AA digestibility in poultry such as nonstarch polysaccharides (Angkanaporn et al., 1994) and phytate (Cowieson and Ravindran, 2007). Nevertheless, there were clear differences in inherent digestibility between the two SBM batches used herein, and this was relevant to both exogenous protease effect and variability at the individual bird level.
Table 6. Effect of soybean origin and protease addition on the apparent ileal digestibility of essential amino acids (EAA), total amino acids (AA), and dry matter (DM) in broilers (n = 18).1
Figure 1. Correlation between the inherent digestibility coefficient of amino acids in the two soybean meal (SBM) batches and the percentage change in the digestibility coefficient associated with exogenous protease addition (R2 = 0.88; P < 0.001). Equation of the trend line: y = 110.6x2 − 211x + 100.46.
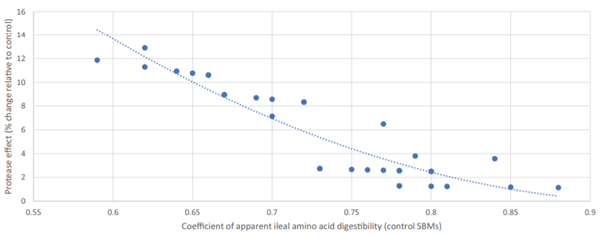
Exogenous protease increased the AID of AA, on average, by 5.7%. However, this varied from 1.1% for Met in the RIV SBM to 12.9% for Asp in the BEC SBM. Generally, the effect of protease was inversely proportional to the inherent digestibility of AA in the two SBM batches (Figure 1), and this explained 85% of the variance in protease response. Similar effects have been reported previously for protease (Cowieson and Roos, 2014, 2016) and also for carbohydrases (Cowieson and Bedford, 2009) and phytase (Cowieson et al., 2017ab). These patterns of response are suggestive of two major principles. First, the magnitude of effect of exogenous enzymes is directly related to the quantity of undigested nutrients in the small intestine, and second, the effect of exogenous enzymes is linked, in part, to alternations in the flow of endogenous protein (and possibly other nutrients).
As far as the authors are aware, this is the first experiment that has reported variance in AA digestibility at the individual bird level under ad libitum feeding conditions. Angkanaporn et al. (1996) used individually caged 5-week-old broiler chickens to assess the apparent and true digestibility of AA in various feed ingredients, but this was assessed using the precision feeding technique, and the authors did not report variance at the individual bird level. Some work has been reported on the contribution of individual birds to variation in the AME of a wheat-based diet (Hughes and Choct, 1997), and it is logical that a similar variation would be expected for digestibility of AA (and also for phosphorus and other nutrients). The reason why some individual birds do not adequately digest energy or AA is not clear but could be related to natural variation in absorptive capacity, digestion rate (perhaps endogenous enzyme function or acidification potential), variable microbiome influences, feed intake or feeding behavior, subclinical disease status, or others. Importantly, when a diet or feed ingredient with an inherently low digestibility is encountered, the natural variability in digestive capacity within a population appears to be significantly exaggerated, leading to a highly deleterious outcome for flock uniformity. In the present work, the relatively poor-quality BEC SBM returned a CV for AA digestibility coefficients of 11.4% with a range of 0.54–0.80 across 18 individual birds. This contrasts with the same for the RIV SBM of a CV of 7.7% and a range of 0.64–0.87. Importantly, the addition of exogenous protease to these SBM batches reduced the variation in AA digestibility coefficients (from 11.4 to 9.1% in the BEC SBM and from 7.7 to 6.3% in the RIV SBM). This suggests that exogenous protease supports birds in the population with a relatively poor digestive capacity and mitigates some of the disadvantages associated with variability in feed ingredient quality.
Table 7. Variability in apparent ileal digestibility coefficients of selected amino acids associated with individual birds fed with two distinct SBM batches, without or with exogenous protease addition (n = 18 per group; 72 birds in total).
Finally, there are significant correlations between AA for all birds in the present experiment (data not shown), which suggests that the variability in digestive potential is generic and not specifically associated with certain proteins or AA. For example, the correlation between AID coefficients of Met and Lys was highly significant and had an r2 of 0.86. In the case of nonessential AA to essential AA, the r2 was 0.99. These relationships indicate that a bird with a compromised capacity to digest AA has similar digestive constraints for all AA, not just for specific AA, and so may benefit from generic support to enhance protein solubility in the intestine or to improve absorptive function.
It can be concluded that SBM quality varies and that this variation is not necessarily always fully explained by conventional metrics of quality such as solubility in KOH or urease activity. Furthermore, a secondary, but significant, consequence of ingestion of low-quality SBM by broilers is an exaggeration of interbird variability. These issues in broiler heterogeneity require further exploration to establish whether specific dietary, management, or genetic interventions may mitigate the risks associated with individuality of digestive physiology and the interaction of this with diet variance. Importantly, the use of exogenous protease in the present work significantly increased the ileal digestibility of AA and reduced the variability at the individual bird level. Thus, exogenous protease may offer a unique opportunity to both reduce nutritional input cost (depending on shadow prices of the displaced nutrients in formulation relative to the cost of the enzyme) and also improve the productivity and uniformity of bird populations with undesirable levels of phenotypic heterogeneity. Exogenous protease may then form part of a nutritional solution package that improves the resilience of birds with poor digestive capacity and reduces the negative consequences on production when such birds are inadvertently fed with a diet of relatively poor nutritional value.
ACKNOWLEDGMENT
The work was financially supported by DSM Nutritional Products, Kaiserausgt, Switzerland.