Sow feed intake during lactation
Nutritional Aspects of Sow Longevity
Published: July 14, 2011
By: Dale W. Rozeboom, Michigan State University/ Reviewer: Lee J. Johnston (University of Minnesota)
Introduction
Longevity of a breeding female is influenced by a wide variety of factors, one of which is the nutritional management of the female throughout her productive life. In modern pork production, the female becomes a part of the breeding herd at birth. Hopefully, several reproductive cycles follow before her removal from the breeding herd. A sound feeding program focuses nutrition throughout the whole life span of the female and does not consider any one phase of growth or reproduction significantly more important than others.
Gilts and sows are culled from the breeding herd for several reasons (unacceptable phenotype, anestrus, failure to conceive, failure to maintain pregnancy, lactation failure, locomotive problems, temperament, small litter size, and faster genetic improvement). Several of these decisions to cull may be related to extremes in body composition. Females that are too lean tend to experience low litter weaning weights, poor return to estrus, smaller subsequent litter size, and physical weakness. Similarly problematic, females that are too fat tend to experience anestrus, dystocia, depressed appetite in lactation, poorer milk production, and locomotive failure. Maintenance of body tissue reserves throughout each phase of a female´s lifetime is an important management goal of farrowing and breeding managers in efforts to increase sow longevity. Improper nutrition could be a direct or indirect cause of these conditions that lead to premature culling. Sound nutritional management can improve performance and increase the chance that females will stay in the herd for a longer period of time. Nutrition of the highly productive, maternalline female revolves around her genetic potential to produce pigs and milk, and her ability to consume feed.
Objectives
- To address selected issues related to nutrition of the modern female during rearing, gestation, lactation, and the weaning-to-rebreeding phases of her life.
- Describe how improper nutrition could be a direct or indirect cause of conditions that lead to premature culling.
- Outline sound nutritional management practices that can improve performance and increase the chance that females will stay in the herd for a longer period of time.
Rearing
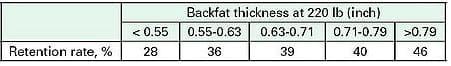
Table 1. Sow retention rate (% to 4th parity) as influenced by backfat taken at 0 lb. Adapted from Gueblez et al., [1]
Body Condition. Numerous researchers have studied the influence of gilt body composition at first breeding on reproductive longevity, thinking that gilt nutrition during rearing and related body nutrient (fat and protein) stores may influence lifetime performance of the sow. The conclusions reached have been variable. Classic survey data [1], shown in Table 1, indicates a positive relationship between gilt backfat depth at 220 pounds body weight and the ability to farrow four litters. These findings are consistent with those found in other studies [2-4]. Consequently, gilt development recommendations to producers from industry and several universities are to delay breeding until gilts have a backfat depth of 0.8 inch or more and weigh about 280 pounds. In contrast, there is ample experimental data, using various genetics and in different production systems, indicating that body condition of gilts at first successful breeding is not related to culling rate [5-9]. Disagreement among the conclusions made in these studies may be due to differing experimental methods. However, Kirkwood [10] wrote that when modern females are subjected to good management that minimizes weight and condition loss during lactation and restores body condition during gestation there is no association between live weight or backfat depth at first successful breeding and subsequent reproductive performance.
The debate about the importance of body composition at first mating and sow longevity is ongoing, with current possible rearing nutrition strategies reflecting to varying degrees, aspects of both sides of the argument. For the most part, current feeding strategies for gilt development reflect the belief that body composition at first breeding is important for sow longevity. Having body tissue minimums (i.e. age, weight, and backfat depth) at gilt breeding is "insurance" against thin sow attrition later, by lessening the impact of possible gaps in sow lactation and gestation management. Consequently, gilt rearing nutrition strategies reflect lean, fat, and skeletal accretion potentials of the varied maternal genotypes being raised.
Feeding Strategies. In large pork production systems, innovative nutritional approaches are being researched and implemented. Breeding gilts are being reared in separate facilities using unique management and nutrition to maximize health and reproductive potential. This is in contrast to the traditional strategy of feeding developing replacement gilts to maximize lean growth rate and efficiency in the grow-finish barn along with barrows and gilts destined for market. Because this has been the default method of rearing breeding gilts for so long, the impact of nutrition during rearing on longevity of the breeding female has gone uninvestigated until recently.
There are no reports describing the direct effect of nursery nutrition (birth to 55 lb) on subsequent lifetime sow productivity. There has however been research done evaluating grow-finish nutrition and longevity. In addition to the decreasingly-used market hog nutrition strategy, there are currently four other researched strategies for feeding the gilt during the grow-finish phase to manipulate body nutrient stores and physiologic systems: (1) market hog nutrition, (2) increase fat/limit lean, (3) limit fat/increase lean, (4) patterned growth, and (5) maximum skeletal development.
Producers with an "ultra-lean" genotype may use greater dietary energy densities and lower protein concentrations in a strategy to increase body fat accumulation and limit lean tissue growth. Producers use this approach because of the importance of body fat reserves for lactation and rebreeding during parity one, and possibly parity two. Certain lean maternal females may only have 0.5-inch backfat at 170 days of age [9,11-12], and if fed primarily for maximum lean gain during rearing may not deposit enough body fat to meet the metabolic needs of subsequent production.
Whether this strategy is correct is being debated. It is supported by a study sponsored by the National Pork Producers Council (currently National Pork Board) [9], in which this strategy in finishing from 150 to 250 lb., positively influenced sow longevity. Gilts fed a moderate lysine (0.6%; about 13 to 16 g/d), high energy (1.59 Mcal/lb) corn-soybean meal diet were more likely to complete 4 parities (56% verses 35%, respectively) than gilts fed a traditional finishing diet to maximize growth (0.95% lysine, 1.48 Mcal/lb corn soybean meal). However, a more recent study suggests not using the "increase fat/limit lean" strategy. Varley and Cole [13] compared 0.55% and 0.85% dietary lysine from 130 lb live weight to first service. After three parities, more females had been culled with the low lysine or limit lean treatment. Their conclusion is supported by the work of King and Williams [14] who first suggested that body protein may be more important than body fat for longevity. The sow catabolizes both body fat and protein stores during lactation. Catabolism of fat and protein stores during lactation is unavoidable in sows nursing large litters and thus both are important for milk production and subsequent return-to-estrus. Limiting the amount of protein stores accumulated during gilt development is potentially dangerous. Amino acids should be provided at or near the gilt´s lean growth potential.
Further caution should be taken if implementing an "increase fat/limit lean" protein- restrictive gilt development program as the study of Cia and coworkers [15] has shown that a low dietary lysine-to-energy ratio (1.34 g lysine per Mcal ME) near breeding compromises estrous expression and ovulation rate in gilts.
Producers with an "average-lean" maternal genotype may use lower dietary energy densities and greater protein concentrations in strategy limit body fatness, while maintaining maximum lean growth. "Averagelean" gilt lines often get too fat and experience poorer lactation feed intakes, less milk production and lower litter growth rates. This "limit fat/increase lean" strategy has been studied more extensively than the "increase fat/limit lean" strategy, and with greater agreement about a positive effect on longevity.
Limiting energy intake of gilts during rearing has increased sow longevity in research conducted in the Netherlands [16-19]. From 12 to 38 wk of age, gilts were fed one of four energy levels: Ad libitum, 83%, 70% and 60% of ad libitum (vitamin, mineral and protein intakes were similar for all treatment groups). Restricting energy intake during rearing delayed pubertal estrus slightly, but did result in a greater parity one conception rate. Litter size did not differ for any parity. Gilts reared on lower energy tended to farrow more litters before being culled. A noticeably large number of gilts (43%) were culled prior to farrowing their first litter, with more (54% and 49%) reared on the two highest energy intakes. For all treatments combined, reproductive disorders (anestrus, no conception, and abortion) were the primary reason for culling gilts. However, for the ad libitum group of gilts alone, leg weakness was noted as an "important" cause for culling. No differences in culling pattern were reported among treatments following parity one. These results are consistent with those found in other studies [20-21].
Other research reports suggest slightly differing results in sow longevity with limited energy during rearing. Snoeyen [22] and van Erp [23] both reported that the number of gilts culled during rearing was decreased with 10 to 20% feed restriction from 55 to 220 pounds compared to ad libitum feeding. However, of the gilts that were not culled before breeding, those given ad libitum access to feed during rearing produced more litters. Similar findings were reported by van de Kerk and de Witt (1983; summarized by te Brake, [19]). Simmins et al. [24] reported no effect of rearing nutrition on culling of gilts prefarrow, but described a detrimental effect of energy restriction during rearing on multiparous sow performance. Conceding that the number of animals included in their study was small, these researchers noted that more sows were culled throughout the eight-parity study for infertility if fed restrictively during rearing. These results may be a consequence of subjecting moderately-grown gilts to inferior gestation and lactation management, which do not minimize weight and condition loss during reproductive cycles.
Long-term improvements in longevity with the "limit fat/increase lean" strategy have been shown by the National Pork Producers Council (currently National Pork Board; [9]) study mentioned above. Gilts fed 4.0 lb/d of a high lysine (1.31%; 23 g/d), moderate energy (1.45 Mcal/lb) corn soybean meal diet, in finishing from 150 to 250 lb., were more likely to complete 4 parities (55% verses 35%, respectively) than gilts fed a traditional finishing diet to maximize growth (0.95% lysine, 1.48 Mcal/lb corn soybean meal). Reasons for culling in this study were not recorded and therefore no explanation is provided as to why gilts experienced different reproductive life times.
Excellent management is required with a restrictive energy development program. Finishing barns need to be equipped with feeding systems that allow restrictive feeding without increasing variation in gilt growth and size within pens. Delayed puberty and estrous irregularity in gilts can be the result of feed restrictions imposed too close to puberty and breeding.
The effects of a growth-altering pre-pubertal feeding regimen, using altering periods of limited growth and maximal growth to achieve patterned growth, on mammary development and first parity milk production have been studied with dairy heifers [25] and swine [26-27]. The periods of limited growth in the swine studies were accomplished by the ad libitum feeding of high fiber diets. During rearing, growth pattern was successfully altered with an overall decrease in ADG. This change was accompanied by an increase in parity-one milk production and parity-one appetite.
The impact of patterned growth during rearing on sow longevity has been studied by Lyvers-Peffer and coworkers [28]. With patterned growth during the pre-pubertal grow-finish period, the incidence of culling prior to completing parity one was decreased with fewer gilts culled for locomotive failure. Sow longevity after parity one was not changed. There is a growing amount of evidence suggesting that slightly-slower patterned growth of modern animals, which have been genetically selected for extremely fast growth, is beneficial for increasing longevity by protecting several physiological systems, including the skeleton [29].
The importance of dietary calcium (Ca) and phosphorus (P) intake by developing gilts for maximizing skeletal development and sow longevity is another aspect of gilt rearing nutrition currently being debated within the swine industry. Nimmo et al. [30] reported that 30% of gilts reared on 0.65% Ca and 0.50% P from weaning to 220 lb had to be culled early in their first gestation because they had become physically disabled. Gilts fed 0.975% Ca and 0.75% P encountered no such problems. These concentrations are considered appropriate for maximum bone mineralization. These results have not been repeated in three different studies conducted subsequently, each involving a far greater number of sows [31-33].
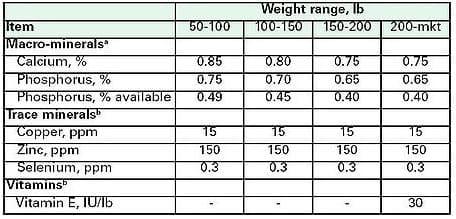
Table 2. Nutrient recommendations for developing gilts [35]aValues are total dietary levels, bValues are supplemental levels
Current recommendations [34] provide the minimal requirements for maximal bone strength, 0.60% Ca and 0.55% P from about 110 to 180 lb live weight, and 0.55% Ca and 0.50% P from 180 to 220 lb live weight. These recommendations are 0.1 percent higher than those provided by NRC for grow-finish hogs of the same weight. The committee authoring the 1998 NRC publication makes this suggestion in spite of stating themselves that "maximization of bone strength does not necessarily improve structural soundness, good health, or longevity." Feeding more Ca and P than that needed to maximize bone mineralization is costly and presents environmental concerns as well.
Most vitamin and mineral requirements of the developing gilt are similar to those of the grow-finish hog raised for market, with recommendations for greater concentrations being suggested [35]. This group of nutritionists suggests that greater copper and zinc concentrations should be fed with higher Ca and P concentration when feeding to maximize bone mineral reserves (Table 2). Greater selenium and vitamin E intake and retention may be beneficial later in the sow´s reproductive life; maximizing the animal´s immuno-response and preventing lactation complications such as mastitis.
Gestation
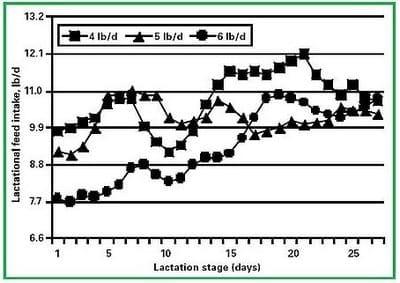
Figure 1. Effect of gestation feed intake on lactational feed intake in gilts (Dourmad, 1991)
Gestation feeding programs are designed to provide sufficient nutrients for development of the fetuses and associated uterine tissues, development of the mammary gland, and modest increases in maternal body weight. If given the opportunity, sows will consume feed far in excess of these needs leading to overly fat sows at farrowing. Several studies worldwide have demonstrated that increasing feed intake during gestation and the associated large increases in sow body weight depress feed intake of lactating sows [36-40] (Figure 1). Low lactation feed intake can depress sow performance and shorten the sow´s reproductive life.
Restricting feed intake during gestation limits excessive pregnancy weight gains of sows. However, severe restriction (60 to 70 % of recommended energy intake) will decrease the sow´s longevity in the breeding herd due to her inability to withstand repeated reproductive cycles [41-42]. One method to restrict energy intake and control weight gain during gestation is to increase the dietary fiber level. In addition to controlling energy intake, dietary fiber may confer some added benefits by increasing litter size at farrowing and improving voluntary feed intake in the subsequent lactation.
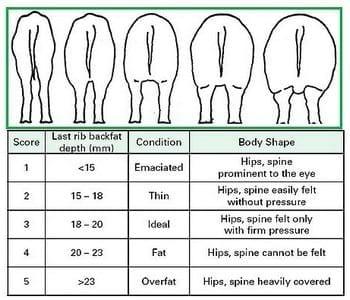
Figure 2. Condition scores of sows (Patience and Thacker, 1989).
Gestating sows should be fed to a target body condition at farrowing and a target weight gain during gestation. Colleagues at the Prairie Swine Center in Saskatchewan have developed sketches of a 5-point body condition scoring system for sows (Figure 2). Sows should enter the farrowing house with a body condition score of about 3. These condition scores can be approximated by last rib backfat depths that can be determined ultrasonically. Backfat depths for younger sows or fatter genotypes will probably be higher for a specific condition score. Target weight gains during gestation should be 80-100 pounds for parity 1, 80-90 pounds for parity 2-5, and 55 pounds for sows´ greater than fifth parity. Recently, two studies have been conducted, in efforts to make backfat depth measurements more useful in commercial production feeding decisions [43-44]. Both successfully demonstrated that monitoring and feeding sows based on backfat measures results in more sows "in" their targeted body condition at given points in the reproductive cycle, but both were unable to identify an improvement in reproductive performance, and indirectly longevity, by doing so. Neither study completed a direct evaluation of longevity.
In the study designed by Straw et al. [43], backfat measurements were taken in eighteen herds using a single point-in-time or snapshot approach to see if there was a relationship between the resulting whole-herd, backfat profile, and reproductive performance (computerized records for the six-month period around the time backfat measurements were obtained). Each herd profile revealed the herd´s average backfat depth, the trend in backfat change with advancing parities, and the gain or loss of backfat between production stages, especially gestation gain and lactation loss. A preliminary summary of the backfat and selected performance measures are shown in Table 3.
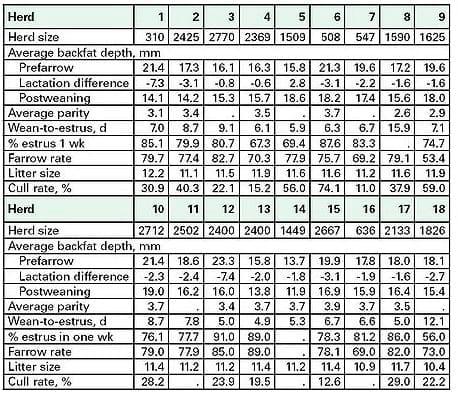
Table 3. Preliminary and partial summary of data for eighteen Michigan swine breeding herds during the summer of 1999 (adapted from Rozeboom, [46]).
Backfat depth in the four fattest herds was 19 to 20 mm, as compared 13 to 15 mm in the three thinnest herds. Backfat depth, across all sows and across all stages of production, ranged from 2.5 to 42 mm. Statistical analysis failed to identify performance differences across herds which could be attributable to the herd´s average backfat depth. It was also observed that reproductive performance was poorly correlated with back depth difference between pre- and post-lactation. In all herds, the use of a snapshot backfat assessment was not appreciably useful in evaluating or improving a given commercial operation´s nutritional program and reproductive performance.
Young and coworkers [44-45] designed an experiment on a large commercial farm to compare three methods of feeding sows in gestation, two of which adjusted feed offered based on backfat depth and weight at weaning of sows or at servicing of gilts:
1. Feed allowance determined by farm manager using traditional body condition scoring and a desired farrowing score of three.
2. Feed allowance set at breeding according to a backfat-weight matrix to achieve a target backfat of 19 mm at farrowing. Each sow´s feed allowance was constant from d 0 to 101 of gestation, and increased 0.9 kg/d from d 102 to farrowing.
3. Feed allowances same as method 2, unless sows were thin (< 15 mm backfat at service), then feed allowance was altered to achieve a backfat depth of 19 mm by d 36 of gestation.
2. Feed allowance set at breeding according to a backfat-weight matrix to achieve a target backfat of 19 mm at farrowing. Each sow´s feed allowance was constant from d 0 to 101 of gestation, and increased 0.9 kg/d from d 102 to farrowing.
3. Feed allowances same as method 2, unless sows were thin (< 15 mm backfat at service), then feed allowance was altered to achieve a backfat depth of 19 mm by d 36 of gestation.
Using weight and backfat in methods 2 and 3 to set sow feed allowance in gestation versus the "manager´s eye" method was successful in getting more sows within a backfat depth of 19 to 21 mm. The number of fat sows (> 21 mm) at farrowing was reduced by both methods as well. However, the numer of thin sows (< 17 mm) at farrowing was similar for all three feeding methods. Thin sows targeted to gain 6 to 9 mm of backfat during gestation, did not do so. Sow performance in lactation and postweaning was not affected by gestation feeding method. Feed intake is often increased to mirror the rapid increase in growth rate of the developing litter so that the sow can maintain a positive energy and nutrient balance in late gestation. This approach can be useful to increase low birth weight of pigs [36]. However, feed intake should only be increased enough to satisfy the growing litter´s demands. One additional pound of a corn-soybean meal diet will provide sufficient nutrients for the increased growth of the litter; amounts greater than this will potentially increase weight gains of the sow which may be detrimental to feed intake during lactation.
The target weight gains and body condition scores listed above can be achieved in many herds if nutrient intake of pregnant sows is limited to specific guidelines (Table 4).
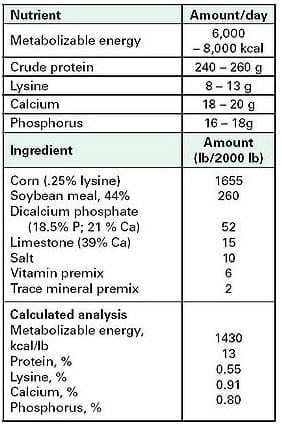
Table 4. Recommended daily intake of energy and selected nutrients for gestating sows, and an example gestation diet with calculated analysis.
Recommended daily intake of nutrients can be achieved by feeding 4.5-6.0 lb of the example corn-soybean meal based diet shown. Exceptions to these guidelines are made for parity one sows and modern high-producing sows. Mahan [47] has reported improved parity-one lactation performance when pregnant gilts had greater daily intakes of a 16% crude protein diet in gestation. The response of primiparous females was attributed to better provision of nutrients for females who were still experiencing significant body growth, and to a quicker adjustment to lactation (greater feed intake and litter gains in week one). Dourmad and coworkers [48] concluded that sows nursing 12 pigs and mobilizing high amounts of body reserves require approximately 8,500 kcal of digestible energy or 6 lb of a corn-soybean meal diet in the following gestation to rebuild protein and fat stores.
Typical corn-soybean meal sow diets are fortified with vitamins and trace minerals for optimal performance. The water soluble vitamin, biotin, may have a direct effect on longevity. Biotin supplementation of sow diets at about 200 mg per ton has been shown to decrease the incidence of hoof cracks and lesions. This improvement in hoof integrity will decrease the incidence of lameness and locomotion problems which are a common reason for culling sows.
Lactation
Unlike gestation, the primary goal during lactation is to maximize energy and nutrient intake of the sow. High nutrient intake is necessary to satisfy the large demands of milk production which typically constitute 70 - 80% of the sow´s total nutrient needs during this period of the reproductive cycle. If these nutrient needs are not satisfied by the diet, sows will break down body tissue to supply nutrients for milk production. Excessive mobilization of body tissues results in low milk production, delayed return to estrus postweaning, and potentially smaller subsequent litter size. High nutrient intake can be achieved by increasing feed intake and/or increasing the nutrient density of the diet. The need for high nutrient intake suggests that feed ingredients like wheat bran, oats, alfalfa meal, and other fibrous feedstuffs are not well suited in most instances for lactating sow diets because they are substantially lower in energy than ingredients such as corn, sorghum, wheat, etc. Choice of nutrient dense feed ingredients is especially important when one realizes that our current nutritional and management programs typically do not allow the sow to express her full genetic potential for milk production [49-50].
Nutrient balance is a critical factor in nutrition of lactating sows. Energy and nutrient demands for body maintenance and milk production must be offset by the energy and nutrients extracted from feed and body tissues. If body tissue is required to make excessive contributions to maintain nutrient balance, the sow will respond by reducing milk output or delaying her return to estrus after weaning.
Researchers are not entirely sure how the sow monitors her nutrient balance during lactation but the existing dogma is that the brain can sense various hormones and metabolites that are involved in the metabolism of nutrients extracted from feed and body tissues. A study conducted at the University of Minnesota demonstrated that low levels of insulin in blood on days 7 and 14 of lactation were associated with negative nutrient balance and delayed return to estrus after weaning [51]. Circulating levels of insulin in the blood have been suggested by several research groups to be an important signal to the brain of the sow´s nutrient balance. Monitoring of body metabolism occurs throughout lactation and it appears that nutrient balance in the first 14 days of lactation plays an important role in determining how rapidly a sow will return to estrus after weaning. Restriction of dietary energy during any week of a 3-week lactation prolongs the weaning to estrus interval (Figure 3; [52]).
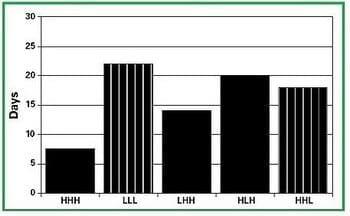
Figure 3. Weaning-to-estrus interval for sows fed high (H) or low (L) energy diets during each week of a 3-week lactation [52].
Delay of reproductive activity after a period of negative nutrient balance may be a defense mechanism developed by the body to wait until there is a more ready supply of nutrients to support another pregnancy and lactation.
Understanding how the sow monitors nutrient balance may have exciting implications for practical swine nutrition. Researchers in Mexico demonstrated that a diet containing 50% cane molasses, a readily available source of carbohydrates, fed during the postweaning period increased circulating insulin levels in sows and resulted in a larger litter size at the next farrowing compared with sows fed an isocaloric diet containing added fat [53]. Other basic research suggests that a diet high in starch elicits higher insulin levels in blood and greater activity of luteinizing hormone in sows compared with a diet at the same energy density but supplemented with fat [54]. Luteinizing hormone plays a key role in initiating occurrence of postweaning estrus. While these basic research studies suggest that composition of the diet can have a direct influence on metabolism of the sow, subsequent experiments have not been able to demonstrate similar responses [55].
In addition to diet composition, a variety of factors influence voluntary feed intake and consequently nutrient balance of lactating sows. In a study of over 25,000 lactations, sows that reached peak daily feed intake earlier had higher average daily feed intake over the entire lactation than sows that took longer to reach peak intake [56]. However, it is a common practice in many herds to gradually increase feed intake during the first week of lactation so that a major transient reduction in feed intake later in lactation can be avoided. This approach may be wise in herds that have documented frequent occurrence of a transient decline in feed intake. However in herds that do not consistently experience this dip in feed intake, undue restriction could place the sow in negative nutrient balance during the early portion of lactation. Every attempt should be made to bring lactating sows up to full feed as soon as possible after farrowing. If this approach causes a depression in feed intake later in lactation, a stepwise increase in the amount of feed offered during the first 4 days of lactation may be warranted. Consider feeding sows 3 lb of feed on the day of farrowing, then increase amount of feed offered 3 lb/day until sows are on full feed about day 4 postfarrowing. This aggressive feeding approach may be particularly important in operations that have short lactations due to SEW production systems. Remember, the primary goal of lactation feeding is to maximize nutrient intake so as to minimize negative nutrient balance.
A large volume of research has focused on the amino acid requirements of lactating sows. It appears that for every pound increase in daily litter weight gain, the sow must consume about 12 grams of lysine to support that weight gain [57]. The sow´s total requirement for lysine and other nutrients must be based on body weight of the sow and the amount of milk being produced (Table 5).
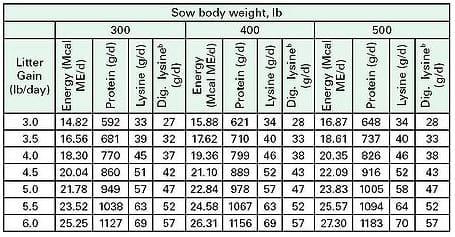
Table 5. Recommended energy, protein, total lysine, and digestible lysine intake for lactating sows by level of production: Adapted from [57]. bAssumes 82% apparent ileal digestibility of total lysine
The values listed in Table 5 are target daily consumption figures assuming sows mobilize 4 g of lysine and 61 g of protein daily. Energy made available in break down of body tissue is not considered in the targets for daily energy intake. In addition to improving milk production, increasing dietary lysine during lactation may increase size of the next litter. The balance of amino acids may be just as important to optimal lactation performance as the quantity of amino acids present in the diet.
Theoretically, ideal protein ratios exactly meet amino acid requirements of an animal with no excesses or deficiencies of amino acids. Ideal protein ratios have been proposed based on experimental evidence for nursery and growing-finishing pigs. To date, ideal protein ratios for lactating sows have been estimated from the amino acid ratios present in sow´s milk. This approach assumes that the profile of amino acids used by the mammary gland to make milk is identical to the profile of amino acids it will extract from blood and that the same profile should be present in the sow´s diet. By directly measuring the amino acids entering and leaving the mammary gland, researchers at the University of Illinois observed that the mammary gland takes up certain essential amino acids, such as arginine, leucine, isoleucine, valine and some others, in larger quantities than are present in milk. This observation suggests the dietary requirement for these amino acids may be different than previously believed based on the amino acid profile of milk [58]. One report suggests that dietary valine should be 117% of lysine to maximize lactation performance of sows [59]. However, without a competitively priced, feed grade source of synthetic valine, this ratio is nearly impossible to achieve in a practical, cost effective diet. Theoretically, improvements in amino acid nutrition of the lactating sow should result in greater preservation of body proteins, primarily muscle mass. It seems reasonable to think that this preservation will increase longevity of the female in the breeding herd. However, this notion awaits direct study. Milk is greater than 80% water so it naturally follows that water plays an important role in lactation. The NCR-89 Committee on Swine Management studied the effects of water flow rate on performance of lactating sows and found that reducing water flow rate from 3 cups to 0.3 cups per minute drastically reduced feed intake, and increased sow weight loss while 21-day litter weight was unaffected (Table 6; [60]).
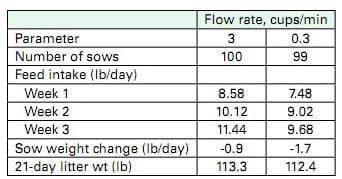
Table 6. Effect of water flow rate from nipple drinkers on feed intake and performance of lactating sows [60]
During hot weather, design and management considerations that provide cool water to lactating sows may increase intake of water and feed and decrease heat stress of sows.
Related to water intake, is salt intake. Interestingly, Seynaeve and coworkers [61] demonstrated the importance of both. Sows fed 0.85% salt verses 0.1% salt in lactation experienced a substantially shorter wean-to-estrus interval (6.2 and 12.6 d, respectively), attributed to greater urine production and improved health.
Calcium and phosphorus nutrition of sows can have a direct impact on longevity of sows through its relationship with the downer sow syndrome. Inadequate calcium and phosphorus absorption due to low dietary concentrations, low feed intake or factors that interfere with absorption of calcium and phosphorus from the gut can increase the sow´s reliance on bone deposits of these minerals. Sows will mobilize minerals from bone stores when dietary intake is insufficient to satisfy lactational demands. The first bones most likely to be affected are the lumbar vertebrae, ribs, and pelvis. If sufficient mobilization occurs, the vertebrae and pelvis become weak increasing the chance of a fracture and the occurrence of downer sow syndrome. As milk production increases so does the demand for calcium and phosphorus [62-63]. Increasing dietary concentration of calcium and phosphorus will not likely improve litter weight gains except in severely deficient diets but it may preserve bone stores of these important macrominerals. Typically, a diet containing .9% calcium, .8% total phosphorus and about 750 IU vitamin D3 per lb will meet the calcium and phosphorus needs of sows consuming 5 lb of feed in gestation and 12 to 14 lb of feed in lactation. Under these conditions, sows will consume 49 - 57 g of calcium and 43 - 50 g of phosphorus daily during lactation. If low feed intake in lactation occurs, increased density of calcium and phosphorus may be necessary.
Although not a nutrient, the addition of the pharmaceutical, chlortetracycline, to the lactation diet has been shown to decrease sow mortality in herds experiencing greater than average mortality [64].
Postweaning
There are no reports describing the direct effect of postweaning sow nutrition on longevity. The primary goal during the postweaning period is to mate the sow and have her conceive a large litter as soon after weaning as possible. For the most part, the timing of events in the postweaning period has been established during lactation and there is little nutritional intervention one can impose after weaning to hasten return to postweaning estrus. Management practices of withholding feed and/or water to "dry off" sows either have no effect or delays return to postweaning estrus. During this period of the reproductive cycle, producers should strive to encourage feed intake so that sows will rebound into positive nutrient balance after the high nutrient demands of late lactation.
Partial or Whole Reproductive Cycle
Longevity may be indirectly improved because of nutritional interventions imposed during more than one phase of reproduction, some fed for a short time and others fed for an extended "loading period." Providing additional omega-3 fatty acids for a short period prior to and during lactation (35 days total) may improve litter size [65], and consequently decrease sow culling due to small litter size. With a six-month loading period (which may include gilt rearing, gestation, and lactation), dietary chromium tripicolinate has been shown to indirectly improve longevity by increasing litter size and percent of sows bred within 7 d of weaning, and decreasing wean-to-estrus interval and sow mortality [66]. In a similar fashion, the addition of L-carnitine at 50 mg/kg of diet in both gestation and lactation, improves wean-to-estrus interval, farrowing rate, and total pigs born over multiple parities [67]; thus longevity. An indirect improvement in sow longevity has been reported by Maxwell and others [68] with the supplementation of mannan oligosaccharide in late gestation and lactation and a resulting decrease in weaning-to-estrus interval.
Summary
Based on the published data directly and indirectly describing the influence of nutrition on longevity of the breeding female, the following recommendations should help improve longevity:
1- The gilt rearing nutrition strategy used should optimize the development of lean, fat, and skeletal systems and not maximize growth rate.
2- With excellent feeding which stores body tissues during gestation and minimizes body tissue losses during lactation, gilts can be bred at various body compositions and still stay in the breeding herd a long time.
3- Control weight gain of sows during gestation by restricting feed intake or by diluting dietary energy density with fibrous feedstuffs.
4- Encourage maximal feed intake during lactation with the goal of maintaining energy and nutrient balance. Target energy and amino acid intakes are based primarily on milk production of the sow.
5- Feed sows aggressively during the first two weeks of lactation as this period is critical in determining success of the rebreeding period.
6- Regularly monitor water flow rate for sows, especially for lactating sows, to ensure that insufficient supply is not depressing voluntary feed intake.
2- With excellent feeding which stores body tissues during gestation and minimizes body tissue losses during lactation, gilts can be bred at various body compositions and still stay in the breeding herd a long time.
3- Control weight gain of sows during gestation by restricting feed intake or by diluting dietary energy density with fibrous feedstuffs.
4- Encourage maximal feed intake during lactation with the goal of maintaining energy and nutrient balance. Target energy and amino acid intakes are based primarily on milk production of the sow.
5- Feed sows aggressively during the first two weeks of lactation as this period is critical in determining success of the rebreeding period.
6- Regularly monitor water flow rate for sows, especially for lactating sows, to ensure that insufficient supply is not depressing voluntary feed intake.
References
1. Gueblez, R, Gestin, JM, Le Henaff, G. Incidence de l´age et de l´ epaisseur de lar dorsal a 100 kg sur la carriere reproductive des truies large white. J. Rech. Porcine en France 1985;17:113.
2. King, RH, Cleary, GV, Maughan, N, Power, C. The effect of initial fat reserves of gilts on their subsequent reproductive performance. Proc. Australian Soc. Anim. Prod. 1984;15:702.
3. Gaughan, JB, Cameron, RDA, McL. Dryden, G, Josey, MJ. Effect of selection for leanness on overall reproductive performance in Large White sows. Anim. Sci. 1995;61:561.
4. Brisbane, JR, Chesnais, JP. Relationship between backfat and sow longevity in Canadian Yorkshire and Landrace pigs. Proc. Nat. Swine Impr. Fed. 1996;21:12.
5. Kirkwood, RN, Mitaru, BN, Gooneratne, AD, Blair, R, Thacker, PA. The influence of dietary energy intake during successive lactations on sow prolificacy. Can. J. Anim. Sci. 1988;68:283.
6. Young, LG, King, GJ, Walton, JS, McMillan, I, Klevorick, M. Age, weight, backfat and time of mating effects on performance of gilts. Can. J. Anim. Sci. 1990;70:469.
7. Newton, EA, Mahan, DC. Effect of initial breeding weight and management system using a high-producing sow genotype on resulting reproductive performance over three parities. J. Anim. Sci. 1993;71:1177.
8. Rozeboom, DW, Pettigrew, JE, Moser, RL, Cornelius, SG, El Kandelgy, SM. Influence of gilt age and body composition at first breeding on sow reproductive performance and longevity. J. Anim. Sci. 1996;74:138.
9. Long, TE. Effects of gilt nutrition and body composition on subsequent reproductive performance. Swine Health Management Certificate Seminar Series. October. International Video and Satellite Series developed by: Michigan State University, University of Nebraska, Iowa State University, University of Minnesota, North Carolina State University, and the University of Guelph. Editor: B. Straw. Released by Michigan State University, Large Animal Clinical Sciences, East Lansing, MI. 1998.
10. Kirkwood, RN. Early puberty, mating reasonable goal. Intern. Pigletter 1990;10(No. 3):9.
11. Close, WM. Managing and feeding the breeding gilt and sow. International Nutrition Supplement: International Pig Topics. 1997; October. Vol. 12, No. 7. pp. ii-iv.
12. Jordan, J. Gilt-edged strategy yields flying start and a lifetime of production. Pig Farming. 1998; March. Vol. 46, No. 3. pp. 33-35.
13. Varley, M, Cole, M. The effect of age at first boar contact, feeding regime and lysine concentration in the diet on lifetime performance in female swine. J. Anim. Sci. 2000; 78 (Supp. 1 ):308 (Abstr.).
14. King, RH, Williams, IH. The effect of nutrition on the reproductive performance of first-litter sows. 2. Protein and energy intakes during lactation. Anim. Prod. 1984;38:249.
15. Cia, MC, Edwards, SA, Glasgow, VL, Shanks, M, Fraser, H. Modification of body composition by altering the dietary lysine to energy ratio during rearing and the effect on reproductive performance of gilts. Anim. Sci. 1998;66:457.
16. den Hartog, LA. The effect of energy intake on development and reproduction of gilts and sows. Ph.D. Dissertation. Agricultural Univ., Wageningen, The Netherlands. 1984.
17. den Hartog, L. A., and G. J. Noordewier. 1984. Effect of energy intake on age at puberty in gilts. Neth. J. Agric. Sci. 32:264.
18. den Hartog, LA. Energy intake and fertility. Pig Int. 1985; April, p.16.
19. te Brake, JHA. Culling sows and the profitability of piglet production. Neth. J. Agric. Res. 1986;34:427.
20. Kirchgessner, VM, Roth-Maier, DA, Neumann, FJ. Zum EinfluB der aufzuchtintensitat auf die spatere reproduktionsleistung von sauen. Zuchtungskunde 1984;56:176.
21. Nielsen, HE, Danielson, V. Nutrition and breeding lifetime. In: 35th Annu. Meeting EAAP, Commission on Anim. Nutr./Commission on Pig Prod., August 6-9, 1984, The Hauge.
22. Snoeyen, P. De invloed van het voerniveau van opfokzeugen op de latere gebruiksduur van fokzuegen. Internal report. Dept. of Anim. Nutr., Agric. Univ. Wageningen, The Netherlands. 1979.
23. van Erp, M. De invloed van het voederniveau van opfokzeugen op de latere gebruikseigenschappen. Internal report. Dept. Anim. Nutr., Agric. Univ. Wageningen, The Netherlands. 1980.
24. Simmins, PH, Edwards, SA, Spechter, HH. Growth and body condition of sows given different feeding regimes during the rearing stage and through eight parities when housed in groups with straw bedding. Anim. Prod. 1994;58:271.
25. Park, CS, Erickson, GM, Choi, YJ, Marx, GD. Effect of compensatory growth on regulation of growth and lactation: Response of dairy heifers to a stair-step growth pattern. J. Anim. Sci. 1987;64:1751-1758.
26. Crenshaw, JD. Feeding gilts to enhance lactational performance. North Central Swine Conf., March 14, Fargo, North Dakota, North Dakota State University, 1990, pp. 33-40.
27. Lyvers-Peffer, PA, Rozeboom, DW. The effects of a growth-altering pre-pubertal feeding regimen on mammary development and parity-one lactation potential in swine. Livest. Prod. Sci. 2001;70:167-173.
28. Lyvers-Peffer, PA, Peng, JJ, Snedegar, JA, Rozeboom, DW. Effects of a growth-altering pre-pubertal feeding regimen on gilt growth and reproductive longevity. J. Anim. Sci. 2003;81(Supp. 1):(Abstr.).
29. Orth, M. Skeletal biology and sow longevity. Proc. Midwest Swine Nutrition Conference, September 9, Indianapolis, IN, 2003, pp 1-8.
30. Nimmo, RD, Peo, ER, Moser, BD, Lewis, AJ. Effect of level of dietary calcium-phosphorus during growth and gestation on performance, blood and bone parameters of swine. J. Anim. Sci. 1981;52:1330.
31. Arthur, SR, Kornegay, ET, Thomas, HR, Veit, HP, Notter, DR, Webb, KE, Baker, JL. Restricted energy intake and elevated calcium and phosphorus intake for gilts during growth. IV. Characterization of metacarpal, metatarsal, femur, humerus and turbinate bones of sows during three parities. J. Anim. Sci. 1983;57:1200.
32. Grandhi, RR, Strain, JH. Dietary calcium-phosphorus levels for growth and reproduction in gilts and sows. Can. J. Anim. Sci. 1983;63:443.
33. Kornegay, ET, Diggs, BG, Hale, OM, Handlin, DL, Hitchock, JP, Barczewski, RA. Reproductive performance of sows fed elevated calcium and phosphorus levels during growth and development. A cooperative study S-145 Committee on Nutritional Systems for Swine to Increase Reproductive Efficiency. J. Anim. Sci. 1984;59 (Suppl. 1):253 (Abstr.).
34. NRC. Nutrient Requirements of Swine (10th Ed.). National Academy Press, Washington, DC. 1998.
35. Tri-State Swine Nutrition Guide. Adeoli, L, Cline, T, Forsyth, D, Hill, G, Mahan, D, Richert, B, Rozeboom, D, Trottier, N. Michigan State University, Ohio State University, Purdue University Cooperating. Michigan State University Extension Publication 1998;E2603.
36. Cromwell, GL, Hall, DD, Clawson, AJ, Combs, GE, Knabe, DA, Maxwell, CV, Noland, PR, Off, DE, Prince, TJ. Effects of additional feed during late gestation on reproductive performance of sows: A cooperative study. J. Anim. Sci. 1989;67:3.
37. Dourmad, JY. Effect of feeding level in the gilt during pregnancy on voluntary feed intake during lactation and changes in body composition during gestation and lactation. Livest. Prod. Sci. 1991;27:309.
38. Weldon, WC, Lewis, AJ, Louis, GF, Kovar, JL, Giesemann, MA, Miller, PS. Postpartum hypophagia in primiparous sows: I. Effects of gestation feeding level on feed intake, feeding behavior, and plasma metabolite concentrations during lactation. J. Anim. Sci. 1994;72:387.
39. Coffey, MT, Diggs, BG, Handlin, DL, Knabe, DA, Maxwell, CV, Noland, PR, Prince, TJ, Cromwell, GL. Effects of dietary energy during gestation and lactation on reproductive performance of sows: A cooperative study. J. Anim. Sci. 1994;72:4.
40. Revell, DK, Williams, IH, Mullan, BP, Smits, RJ. Body fatness influences voluntary feed intake and live weight loss during lactation in primiparous sows. J. Anim. Sci. 1994;72 (Suppl. 1):389 (Abstr.).
41. Simmins, PH, Edwards, SA, Spechter, HH, Riley, JE. Lifetime performance of sows given different rearing and pregnancy feeding regimes. Anim. Prod. 1992;54:457(Abstr.).
42. Dourmad, JY, Etienne, M, Prunier, A, Noblet, J. The effect of energy and protein intake of sows on their longevity: A review. Livest. Prod. Sci. 1994;40: 87.
43. Straw, BE, Dewey, CE, Rozeboom, DW, Bates, RO. Sow Body Condition Scoring, Reproductive Performance and Sow Longevity. Western States Veterinary Conference, Las Vegas, NV, Feb 11-15. Presentation only (no proceedings). 2001.
44. Young, MG, Tokach, MD, Aherne, FX, Main, RG, Dritz, SS, Goodband, RD, Nelssen, JL. Comparison of three methods of feeding sows in gestation. J. Anim. Sci. 2003;81(Supp. 1):59(Abstr.).
45. Young, MG, Tokach, MD, Aherne, FX, Main, RG, Dritz, SS, Goodband, RD, Nelssen, JL. Effect of gestation feeding method on sow performance in lactation. J. Anim. Sci. 2003;81(Supp. 1):59(Abstr.).
46. Rozeboom, DW. Breeding herd energetics. Proc. Midwest Swine Nutrition Conference, September 9, Indianapolis, IN, 2003, pp 1a-6a.
47. Mahan, DC. Relationship of gestation protein and feed intake level over a five-parity period using a high-producing sow genotype. J. Anim. Sci. 1998;76:533.
48. Dourmad, JY, Etienne, M, Noblet, J. Reconstitution of body reserves in multiparous sows during pregnancy: Effect of energy intake during pregnancy and mobilization during the previous lactation. J. Anim. Sci. 1996;74:2211.
49. Stauber, TE, Stahly, TS, Ewan, RC, Williams, NH. Maximum lactational capacity of sows with a high and low genetic capacity for lean tissue growth. J. Anim. Sci. 1994;72(Suppl. 1):364(Abstr.).
50. Boyd, RD, Kensinger, RS, Harrell, RJ, Bauman, DE. Nutrient uptake and endocrine regulation of milk synthesis by mammary tissue of lactating sows. J. Anim. Sci. 1995;73(Suppl. 2):36.
51. Tokach MD, Pettigrew, JE, Dial, GD, Wheaton, JE, Crooker, BA, Johnston, LJ. Characterization of luteinizing hormone secretion in the primiparous, lactating sow: Relationship to blood metabolites and return-to-estrus. J. Anim. Sci. 1992;70:2195.
52. Koketsu, Y, Dial, GD, Pettigrew, JE, Marsh, WE, King, VL. Influence of imposed feed intake patterns during lactation on reproductive performance and on circulating levels of glucose, insulin, and luteinizing hormone in primiparous sows. J. Anim. Sci. 1996;74:1036.
53. Rodriquez-Marquez, MC, Cuaron, JA. Dietary energy source on ovulation in swine. J. Anim. Sci. 1990;68(Suppl. 1):367(Abstr.).
54. Kemp, B, Soede, NM, Helmond, FA, Bosch, MW. Effects of energy source in the diet on reproductive hormones and insulin during lactation and subsequent estrus in multiparous sows. J. Anim. Sci. 1995;73:3022.
55. Johnston, LJ, Pettigrew, JE,Wilson,ME, Walker, RD, Crooker, BA. Influence of dietary molasses on reproductive performance of lactating sows. J. Anim. Sci. 1994;73(Suppl. 1):334(Abstr.).
56. Koketsu, Y, Dial, GD, Pettigrew, JE, Marsh, WE, King, VL. Characterization of feed intake patterns during lactation in commercial swine herds. J. Anim. Sci. 1996;74:1202. 57. Pettigrew, J E. Amino acid nutrition of gestating and lactating sows. Biokyowa Tech. Rev. 5. 1993.
58. Trottier, N. Protein metabolism in the lactating sow. PhD Dissertation. Univ. of IL. 1995.
59. Richert, BT, Tokach, MD, Goodband, RD, Nelssen, JL, Pettigrew, JE, Walker, RD, Johnston, LJ. Valine requirement of the high-producing lactating sow. J. Anim. Sci. 1996;74:1307.
60. NCR-89 Committee on Swine Confinement Management, and Leibbrandt, VD. Effect of nipple drinker water flow rate and season on swine lactation performance. J. Anim. Sci. 1991;69(Suppl. 1):482(Abstr.).
61. Seynaeve, M, De Wilde, R, Janssens, G, De Smet, B. The influence of dietary salt level on water consumption, farrowing and reproductive performance of lactating sows. J. Anim. Sci. 1996;74:1047.
62. Maxson, PF, Mahan, DC. Dietary calcium and phosphorus for lactating swine at high and average production levels. J. Anim. Sci. 1986;63:1163.
63. Mahan, DC, Newton, EA. Effect of initial breeding weight on macro- and micromineral composition over a three-parity period using a high-producing sow genotype. J. Anim. Sci. 1995;73:151.
64. Deen, J, Keay, S, Boyd, RD, Appleton, D, Matzat, P, Wolff, T. Strategic application of Aureomycin chlortetracycline in lactation diets reduces sow mortality and improves reproductive performance. J. Anim. Sci. 2000;78(Suppl.1):173(Abstr.).
65. Webel, SK, Otto-Tice, ER, Moser, RL, Orr, DE. Effect of feeding duration on protected n-3 polyunsaturated fatty acids (FertiliumTM) on litter size and embryo survival in sow. J. Anim. Sci. 2004;82(Suppl. 2):51(Abstr.).
66. Hagen, CD, Lindemann, MD, Purser, KW. Dietary chromium tripicolinate increases sow productivity under commercial conditions. J. Anim. Sci. 1999;77(Suppl.1):67(Abstr.).
67. Real, DE, Nelssen, JL, Tokach, MD, Goodband, RD, Dritz, SS, Owen, KQ. Effects of dietary L-carnitine and chromium picolinate on sow reproductive performance. J. Anim. Sci. 2002;72(Suppl. 2):59(Abstr.).
68. Maxwell, CV, Ferrell, K, Dvorak, RA, Johnson, ZB, Davis, ME. Efficacy of mannan oligosaccharide supplementation through late gestation and lactation on sow and litter performance. J. Anim. Sci. 2003;81(Suppl.2):69(Abstr.).
Originally published on the eXtension Foundation Site. Engormix.com thanks for this contribution.
Related topics:
Authors:


Recommend
Comment
Share

Would you like to discuss another topic? Create a new post to engage with experts in the community.










