Climate, Soil Management, and Cultivar Affect Fusarium Head Blight Incidence and Deoxynivalenol Accumulation in Durum Wheat of Southern Italy
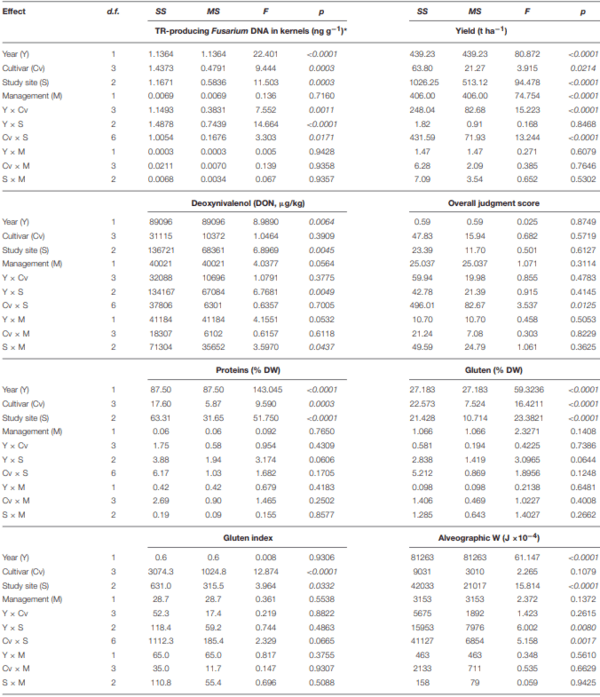
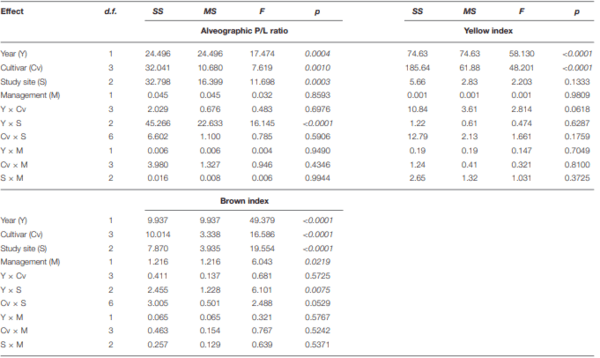
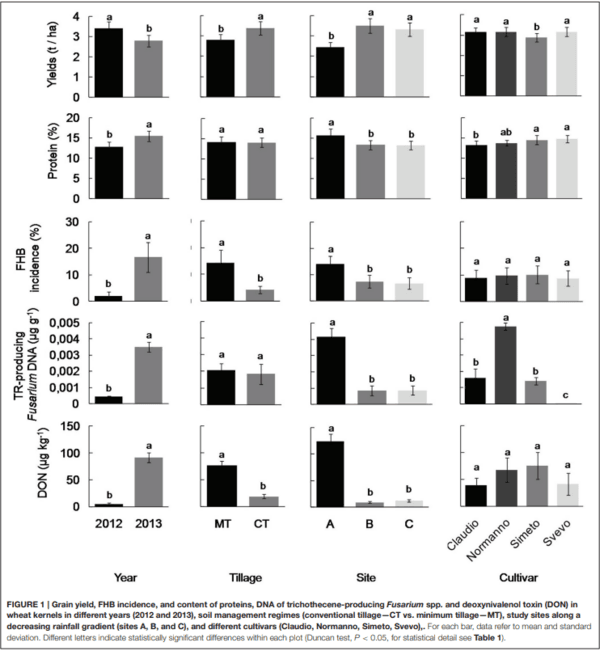
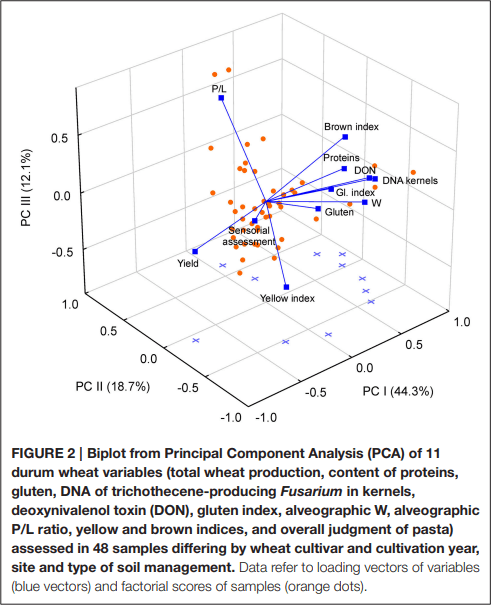
Ames, N. P., Clarke, J. M., Marchylo, B. A., Dexter, J. E., and Woods, S. M. (1999). Effect of environment and genotype on durum wheat gluten strength and pasta viscoelasticity. Cereal Chem. 76, 582–586. doi: 10.1094/CCHEM.1999.76.4.582
Aureli, G., Amoriello, T., Belocchi, A., D’Egidio, M. G., Fornara, M., Melloni, S., et al. (2015). Preliminary survey on the so-occurrence of DON and T2+HT2 toxins in durum wheat in Italy. Cereal Res. Commun. 43:3. doi: 10.1556/0806.43.2015.010
Bernhoft, A., Torp, M., Clasen, P. E., Løes, A. K., and Kristoffersen, A. B. (2012). Influence of agronomic and climatic factors on Fusarium infestation and mycotoxin contamination of cereals in Norway. Food Addit. Contam. 29, 1129–1140. doi: 10.1080/19440049.2012.672476
Berthiller, F., Schuhmacher, R., Adam, G., and Krska, R. (2009). Formation, determination and significance of masked and other conjugated mycotoxins. Anal. Bioanal. Chem. 395, 1243–1252. doi: 10.1007/s00216-009-2874-x
Blanco, A., Colasuonno, P., Gadaleta, A., Mangini, G., Schiavulli, A., Simeone, R., et al. (2011). Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds on durum wheat. J. Cereal Sci. 54, 255–264. doi: 10.1016/j.jcs.2011.07.002
Boutigny, A. L., Forget, F. R., and Barreau, C. (2008). Natural mechanisms for cereal resistance to the accumulation of Fusarium trichothecenes. Eur. J. Plant Pathol. 121, 411–423. doi: 10.1007/s10658-007-9266-x
Brzozowski, B., Dawidziuk, K., and Bednarski, W. (2008). Gliadin degradation by proteases of Fusarium genus fungi in different in vivo and in vitro conditions. Polish J. Nat. Sci. 23, 188–206. doi: 10.2478/v10020-008-0014-2
Burlakoti, R. R., Estrada, R., Rivera, V. V., Boddeda, A., Secor, G. A., and Adhikari, T. B. (2007). Real-time PCR quantification and mycotoxin production of Fusarium graminearum in wheat inoculated with isolates collected from potato, sugar beet, and wheat. Phytopathology 97, 835–841. doi: 10.1094/PHYTO-97-7- 0835
Chandelier, A., Nimal, C., André, F., Planchon, V., and Oger, R. (2011). Fusarium species and DON contamination associated with head blight in winter wheat over a 7-year period (2003–2009) in Belgium. Eur. J. Plant Pathol. 130, 403–414. doi: 10.1007/s10658-011-9762-x
Covarelli, L., Beccari, G., Prodi, A., Generotti, S., Etruschi, F., Juan, C., et al. (2015). Fusarium species, chemotype characterisation and trichothecene contamination of durum and soft wheat in an area of central Italy. J. Sci. Food Agric. 95, 541–551. doi: 10.1002/jsfa.6772
Czaban, J., Wróblewska, B., Sułek, A., Mikos, M., Boguszewska, E., Podolska, G., et al. (2015). Colonisation of winter wheat grain by Fusarium spp. and mycotoxin content as dependent on a wheat variety, crop rotation, a crop management system and weather conditions. Food Addit. Contam. 32, 874–910. doi: 10.1080/19440049.2015.1019939
D’Egidio, M. G., Mariani, B. M., Nardi, S., and Novaro, P. (1993).Viscoelastograph measures and total organic matter test: suitability in evaluating textural characteristics of cooked pasta. Cereal Chem. 70, 67–72.
De Wolf, E. D., Madden, L. V., and Lipps, P. E. (2003). Risk assessment models for wheat Fusarium head blight epidemics based on within season weather data. Phytopathology 93, 428–435. doi: 10.1094/PHYTO.2003.93.4.428
Dexter, J. E., Marchylo, B. A., Clear, R. M., and Clarke, J. M. (1997). Effect of Fusarium head blight on semolina milling and pasta-making quality of durum wheat. Cereal Chem. 74, 519–525. doi: 10.1094/CCHEM.1997.74.5.519
Dill-Macky, R., and Jones, R. K. (2000). The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis. 84, 71–76. doi: 10.1094/PDIS.2000.84.1.71
Edwards, S. G. (2009). Fusarium mycotoxin content of UK organic and conventional wheat. Food Addit. Contam. 26, 496–506. doi: 10.1080/02652030802530679
Edwards, S. G., Pirgozliev, S. R., Hare, M. C., and Jenkinson, P. (2001). Quantification of trichothecene-producing Fusarium species in harvested grain by competitive PCR to determine efficacies of fungicides against Fusarium head blight of winter wheat. Appl. Environ. Microbiol. 67, 1575–1580. doi: 10.1128/AEM.67.4.1575-1580.2001
Fagnano, M., Fiorentino, N., D’Egidio, M. G., Quaranta, F., Ritieni, A., Ferracane, R., et al. (2012). Durum wheat in conventional and organic farming: yield amount and pasta quality in Southern Italy. Sci. World J. 2012:973058. doi: 10.1100/2012/973058
Fedak, G., Cao, W., Xue, A., Savard, M., Clarke, J., and Somers, D. J. (2007). “Wheat production in stressed environments,” in Development in Plant Breeding, eds H.T. Buck, J. E. Nisi, and N. Salomón (Dordrecht: Springer Netherlands), 91–95.
Flagella, Z. (2006). Qualità nutrizionale e tecnologica del frumento duro. Ital. J. Agron. 1, 203–239. doi: 10.4081/ija.2006.s1.203
Flagella, Z., Giuliani, M. M., Giuzio, L., Volpi, C., and Masci, S. (2010). Influence of water deficit on durum wheat storage protein composition and technological quality. Eur. J. Agron. 33, 197–207. doi: 10.1016/j.eja.2010.05.006
Fois, S., Schlichting, L., Marchylo, B., Dexter, J., Motzo, R., and Giunta, F. (2011). Environmental conditions affect semolina quality in durum wheat (Triticum turgidum ssp. durum L.) cultivars with different gluten strength and gluten protein composition. J. Sci. Food Agric. 91, 2664–2673. doi: 10.1002/jsfa.4509
Gauthier, T., Waché, Y., Laffitte, J., Taranu, I., Saeedikouzehkonani, N., Mori, Y., et al. (2013). Deoxynivalenol impairs the immune functions of neutrophils. Mol. Nutr. Food Res. 57, 1026–1036. doi: 10.1002/mnfr.201 200755
Giorni, P., Dall’Asta, C., Reverberi, M., Scala, V., Ludovici, M., Cirlini, M., et al. (2015). Open field study of some Zea mays hybrids, lipid compounds and fumonisins accumulation. Toxins 7, 3657–3670. doi: 10.3390/toxins7 093657
Häller, G. B., Munich, M., Kleijer, G., and Mascher, F. (2008). Characterisation of kernel resistance against Fusarium infection in spring wheat by baking quality and mycotoxin assessments. Eur. J. Plant Pathol. 120, 61–68. doi: 10.1007/s10658-007-9198-5
Hijmans, R. J., S. E., Cameron, J. L., Parra, P. G., Jones, and Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978. doi: 10.1002/joc.1276
Hooker, D. C., Schaafsma, A. W., and Tamburic-Ilincic, L. (2002). Using weather variables pre-and post-heading to predict deoxynivalenol content in winter wheat. Plant Dis. 86, 611–619. doi: 10.1094/PDIS.2002.86.6.611
Jolliffe, I. (2002). Principal Component Analysis. Springer Series in Statistics. New York, NY: Springer-Verlag.
Kelly, A. C., Clear, R. M., O’Donnell, K., McCormick, S., Turkington, T. K., Tekauz, A., et al. (2015). Diversity of Fusarium head blight populations and trichothecene toxin types reveals regional differences in pathogen composition and temporal dynamics. Fungal Genet. Biol. 82, 22–31. doi: 10.1016/j.fgb.2015.05.016
Magan, N., Hope, R., Colleate, A., and Baxter, E. S. (2002). Relationship between growth and mycotoxin production by Fusarium species, biocides and environment. Eur. J. Plant Pathol. 108, 685–690. doi: 10.1023/A:1020618728175
Mangini, G. (2006). Analisi genetica del contenuto proteico delle cariossidi di frumento duro. Tecnica Molitoria Gennaio 2006, 1–12.
Mariani, B. M., D’Egidio, M. G., and Novaro, P. (1995). Durum wheat quality evaluation: influence of genotype and environment. Cereal Chem. 72, 194–197.
Miller, J. D., Culley, J., Fraser, K., Hubbard, S., Meloche, F., Ouellet, T., et al. (1998). Effect of tillage practice on Fusarium head blight of wheat. Can. J. Plant Pathol. 20, 95–103. doi: 10.1080/07060669809500450
Nightingale, M. J., Marchylo, B. A., Clear, R. M., Dexter, J. E., and Preston, K. R. (1999). Fusarium head blight: effect of fungal proteases on wheat storage proteins. Cereal Chem. 76, 150–158. doi: 10.1094/CCHEM.1999.76. 1.150
Pancaldi, D., Tonti, S., Prodi, A., Salomoni, D., Dal Prà, M., Nipoti, P., et al. (2010). Survey of the main causal agents of Fusarium head blight of durum wheat around Bologna, northern Italy. Phytopathol. Mediterr. 49, 258–266. doi: 10.14601/Phytopathol_Mediterr-3442
Pasquali, M., and Migheli, Q. (2014). Genetic approaches to chemotype determination in type B-trichothecene producing Fusaria. Int. J. Food Microbiol. 189, 164–182. doi: 10.1016/j.ijfoodmicro.2014.08.011
Pinheiro, N., Costa, R., Almeida, A. S., Coutinho, J., Gomes, C., and Maçãs, B. (2013). Durum wheat breeding in Mediterranean environment – influence of climatic variables on quality traits. Emir. J. Food Agric. 25, 962–973. doi: 10.9755/ejfa.v25i12.16732
Pitt, J. (1979). The Genus Penicillium. London: Academic Press.
Quaranta, F., Amoriello, T., Aureli, G., Belocchi, A., and D’Egidio, M. G., Fornara, et al. (2010). Grain yield, quality and deoxynivalenol (DON) contamination of durum wheat (Triticum durum Desf.): results of national networks in organic and conventional cropping systems. 2010. Ital. J. Agron. 4, 353–366. doi: 10.4081/ija.2010.353
Raciti, C. N., Doust, M. A., Pattavina, A., Gallina, O., Fiscella, T., Strano, L., et al. (2005). Atti 6◦ Convegno AISTEC “Cereali: biotecnologie e processi innovativi.” Giugno 16–18, 251–257.
Raper, K. B., and Fennell, D. I. (1965). The Genus Aspergillus. Malabar: Krieger Publishing Company Inc.
Reverberi, M., Punelli, M., Scala, V., Scarpari, M., Uva, P., Mentzen, W., et al. (2013). Genotypic and phenotypic versatility of Aspergillus flavus during maize exploitation. PLoS ONE 8:e68735. doi: 10.1371/journal.pone.00 68735
Rocha, O., Ansari, K., and Doohan, F. M. (2005). Effects of trichothecene mycotoxins on eukaryotic cells: a review. Food Addit. Contam. 22, 369–378. doi: 10.1080/02652030500058403
Schaafsma, A. W., Tamburic-Ilincic, L., and Hooker, D. C. (2005). Effect of previous crop, tillage, field size, adjacent crop, and sampling direction on airborne propagules of Gibberella zeae/Fusarium graminearum, Fusarium head blight severity, and deoxynivalenol accumulation in winter wheat. Can. J. Plant Pathol. 27, 217–224. doi: 10.1080/07060660509507219
Shephard, G. S. (2008). Impact of mycotoxins on human health in developing countries. Food Addit. Contam. 25, 146–151. doi: 10.1080/02652030701567442
Soil Survey Staff (1998). Keys to Soil Taxonomy. Washington, DC: United States Department of Agriculture Natural Resources Conservation Service.
Summerell, B. A., Salleh, B., and Leslie, J. F. (2003). A utilitarian approach to Fusarium identification. Plant Dis. 87, 117–128. doi: 10.1094/PDIS.2003.87.2.117
Sutton, J. C. (1982). Epidemiology of wheat blight and maize ear rot caused by Fusarium graminearum. Can. J. Plant Physiol. 4, 195–2019.
Visconti, A., and Pascale, M. (2010). An overview on Fusarium mycotoxins in the durum wheat pasta production chain. Cereal Chem. 87, 21–27. doi: 10.1094/CCHEM-87-1-0021
West, J. S., Holdgate, S., Townsend, J. A., Edwards, S. G., Jennings, P., and Fitt, B. D. (2012). Impacts of changing climate and agronomic factors on fusarium ear blight of wheat in the UK. Fungal Ecol. 5, 53–61. doi: 10.1016/j.funeco.2011.03.003
Xu, X. M., Nicholson, P., Thomsett, M. A., Simpson, D., Cooke, B. M., Doohan, F. M., et al. (2008). Relationship between the fungal complex causing Fusarium head blight of wheat and environmental conditions. Phytopathology 98, 69–78. doi: 10.1094/PHYTO-98-1-0069
Zain, M. E. (2011). Impact of mycotoxins on human and animals. J. Saudi Chem. Soc. 15, 129–144. doi: 10.1016/j.jscs.2010.06.006







.jpg&w=3840&q=75)