Mycotoxins in the food chain: a look at their impact on immunological responses
Published: December 13, 2006
Source : RAGHUBIR P. SHARMA - The University of Georgia (Courtesy of Alltech Inc.)
The immune system is an important defensive mechanism against invading parasitic organisms or foreign cells. The system is highly evolved in mammals and birds. In general, its complexity correlates with the evolutionary level of various animal species. In higher organisms, the system consists of specialized cells found throughout the body; these cells are localized in large quantities in certain organs such as the thymus, spleen, and lymph nodes. Cells of the immune system and cells of the hemopoietic system all originate in the bone marrow. The bone marrow stem cells later differentiate to perform specialized immunologic functions.
Immunotoxicology is a relatively new discipline, although the allergic responses to various chemicals have long been recognized. Chemicals, including mycotoxins, can either suppress or stimulate the immune system. Immunosuppression likely decreases resistance to a variety of infectious diseases and may even predispose the host to the expression and dispersion of cancerous cells. Stimulation of the immune system is not always desirable because it may lead to hypersensitivity (allergic) reactions. The mycotoxin-induced immunotoxicity has been reviewed earlier in detail (Bondy and Pestka, 2000; Sharma, 1985; 1991; 1993).
Complexities of the immune system
The immune system of mammals is highly complex, and various cells of this system interact to produce the desired effect. The immune system includes the innate immune functions that are inherent in different organs and do not require pre-exposure to antigens or chemicals. Some lymphocytes are natural killer cells that generally require no priming or proliferation to exert their effects. On the other hand, the generalized immune system usually confers acquired immunity after the organism has been in contact with infectious organisms or other antigenic determinants.
Lymphocytes and macrophages are cellular units of both types of immune systems. The two major forms of lymphocytes, T cells and B cells, differentiate in the thymus and fetal liver, respectively. The T cells are involved in cell-mediated immune responses, such as delayed hypersensitivity reaction and immune surveillance against foreign or altered cells. Several subpopulations of T cells exist, e.g., cytotoxic T cells, helper T cells, and suppressor T cells. The B cells are primarily involved in the production of a variety of antibodies; however, these cells are often under the control of T cells. The T cells interact with one another or with other cells of the immune system via a variety of soluble factors called cytokines. In some instances direct cell-cell interactions are necessary to mount the optimal immune responses.
Macrophages are derived from monocytes and are present in various body cavities, such as pulmonary alveoli and peritoneum; they are also found in the lymph nodes and liver, as components of the innate immune system. Other peripheral leukocytes are often involved in various immunopathologic mechanisms. For acquired immunity macrophages are phagocytes that concentrate antigens and confer specific immunologic responses to various T or B cells and also remove cell debris.
The immune system interacts with other systems and is profoundly influenced by the central nervous system, both directly via innervations of lymphatic organs and indirectly via neuroendocrine mechanisms. The cells of the immune system produce factors that influence the nervous system. Hormones, such as somatotrophin (growth hormone) and thymosin (thymic maturation factor), stimulate the immune responses, whereas steroids, including sex hormones, generally suppress the immune responses.
Mycotoxins and public health
Mycotoxins are metabolites produced by certain fungi that infest food crops and processed foods. The first recorded episode of mycotoxicosis in public health was recognized in the middle of the 19th century when ergotism was shown to be produced by ingestion of rye infected with Claviceps purpurea. In the first half of the 20th century, the ‘moldy bread disease’ or alimentary toxic aleukia (ALA) was associated with over-wintered wheat or grain in the former Soviet Union infected with various Fusaria species.
The interest and intensive research in the problems with mycotoxins started with the incidence of Turkey-X disease in England, where the peanut meal used in poultry rations was found to be contaminated with
aflatoxins produced by Aspergillus flavus.
Since the 1950s extensive research in the field of mycotoxins has provided information on a number of fungal products that are commonly associated with foods. The most common mycotoxins found in various foods are aflatoxins, fumonisins and trichothecenes, with occasional contamination with other toxins. Commonly encountered mycotoxins associated with various food commodities are listed in Table 1. Although the molds that produce these toxins are ubiquitous, the problem in foods can be easily managed by avoiding the fungal contamination and not consuming moldy or spoiled foods. Toxigenic fungi do not always produce mycotoxins. Their production is often aided by certain temperatures and humidity and stressful conditions to the fungi.
Avoiding storage of foods at conditions favorable to mycotoxin production is highly valuable in controlling the toxic levels of these agents.
Additionally, development of fungus-resistant crops has been somewhat useful in preventing the growth
of toxigenic fungi.
In public health, the greater problem with mycotoxins is in developing countries rather than in the developed countries. In industrialized countries the food resources are plentiful, handling and preservation technology is well developed, and various regulations limiting the exposure to mycotoxins are in effect. On the other hand in developing countries, where food supplies are inadequate and storage facilities are not yet optimal, there is a greater chance for food spoilage and contamination by toxigenic molds. Even in developed countries; however, problems with mycotoxins frequently occur in livestock health.
Table 1. Common mycotoxins contaminating foods a

aOnly common mycotoxins and representative fungi andcommodities associated with them are indicated.
Prevalence of mycotoxins in foods
Mycotoxins are inadvertent contaminants in foods as the fungi that produce them are fairly widespread.
In spite of a large amount of research available for various mycotoxins, information regarding their occurrence in foods is limited. In many cases, the information is limited to periods when episodes of adverse human or animal health effects have been observed. Only a few of the mycotoxins are routinely analyzed in foods during surveys. The US Department of Agriculture and Food and Drug Administration have established limits for various mycotoxins in foods and feeds (Food and Drug administration, 2004). Still, food commodities sometimes exceed the allowed limits and may in fact be of concern in public health. Table 2 lists the levels of selected mycotoxins associated with different food commodities and their action levels established by US Food and Drug Administration. Additional information and primary sources of mycotoxin levels in foods are available from the recent reviews or other publications (Campbell et al., 2003; Council for Agricultural Science and Technology, 2003; Dutten, 1996; Gilbert and Vargas, 2003; Harwig et al., 1973; Osborne, 1980; Shepard, 2003; Shotwell et al., 1971; Wilson and Nuovo, 1973).
One of the public health concerns with the presence of mycotoxins in foods is that these toxins are relatively stable; once they are formed they can persist for a long time. Only a few of them are partially destroyed by cooking, pasteurization, or storage. The concentration of a mycotoxin may indeed increase during the fermentation process or storage if the toxigenic fungi are present in food. The mere presence of fungi however is not always indicative of the occurrence of mycotoxins, since the organisms produce these toxic metabolites only under certain conditions that are determined by temperature, humidity or a lack of normal nutrients for the mold.
The occurrence of mycotoxins in agricultural commodities depends on factors such as geographical region, season, and the conditions under which a certain crop is grown, harvested and stored. There are no definite rules for if and when a mold will produce a mycotoxin. The fungal spoilage of crops and grains may be enhanced by drought, insect damage, cracked kernels during harvesting, and presence of excessive chaff in the harvested grain.
Table 2. Mycotoxin levelsa encountered in foods and their FDA action levels 1.
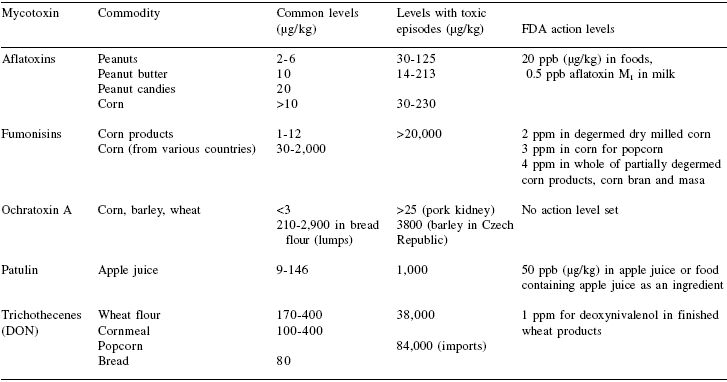
Click to see the image in real size.
aLevels are only from some representative reports. Wide ranges in concentrations have been reported, particularly in contaminated samples.
1FDA, 2004.
Common health problems with mycotoxins in foods
AFLATOXINS
Metabolic products of Aspergillus flavus and A. parasiticus occur in foods in a number of susceptible commodities. Peanuts, corn, other nuts and grains are possible food commodities that may be contaminated.
Aflatoxin B1 is the most potent and prevalent of this group. Aflatoxin B1 is activated in vivo by metabolizing enzymes to an epoxide, which is very reactive and binds to various biological molecules, including specific bases in the DNA. After dietary exposure aflatoxin B1 is metabolized in the liver, the organ with a high level of metabolizing enzymes, and produces damage to this organ, including hepatocarcinogenesis. Indeed in some species, especially the rainbow trout, aflatoxin B1 is the most potent carcinogen known. In the short term, aflatoxin B1 causes necrosis in the liver and possibly damage to other organs such as kidney, heart, spleen and pancreas. In animals exposed to aflatoxin B1 decreased productivity and high mortality due to infections are often the consequence.
Several episodes of aflatoxin B1 poisoning in humans have been reported. These include both general effects of this mycotoxin and carcinogenesis.
Symptoms of suspected outbreaks of poisoning include jaundice, rapidly developing ascites, partial hypotension and death. In many parts of the world where hepatocellular carcinoma is prevalent, a high level of aflatoxin in the diet has been documented.
However, many epidemiological surveys suggest the concurrent presence of hepatitis B virus and other mycotoxins such as fumonisins that may be contributory factors in addition to the aflatoxin B1 (see later).
FUMONISINS
Fumonisins are toxins produced by Fusarium verticillioides and other Fusarium species commonly found on corn (Riley et al., 2001). Fumonisin B1 is the most toxic and prevalent of these mycotoxins; its presence has been demonstrated in cornmeal, breakfast cereals and corn tortillas. Fumonisins have produced fatal diseases in livestock, including equine leukoencephalomalacia (ELEM, a rapidly developing brain degeneration) and porcine pulmonary edema (PPE or hydrothorax, lethal in a few days). An association of the presence of the fungus producing fumonisins and the presence of mycotoxins themselves has been reported with human esophageal cancer and primary liver cancer. The latter has been implicated when fumonisins are co-contaminants of foodstuffs with aflatoxins. Recently, carcinogenicity of fumonisin B1 has been demonstrated in laboratory animals, including mice (hepatoma in females) and rats (renal carcinoma in males).
The species and gender specificity of fumonisin B1 is puzzling and the basis for it has not been discerned. Fumonisin B1 is a structural analog of one of the primary sphingolipid bases, sphinganine, and thereby competes for the incorporation of sphinganine into ceramide and ultimately in complex sphingolipids (Riley et al., 1998, 2001). A rise in cellular levels of sphinganine and sphingosine (dehydrosphinganine, another important free sphingoid base that is also a known signaling agent) is a uniform observation in all tissues exposed to this mycotoxin. These free sphingolipid bases (sphingosine and sphinganine) as well as their phosphate derivations are major signaling molecules that often possess paradoxical effects, characterized by either death or survival signals in cells.
The occurrence of fumonisins also depends on the fraction of corn produced during processing. For example, dry milling of corn kernels results in bran, flaking grits, grits, meal and flour. Fumonisins are found in greatest quantity in the bran fraction, followed in order by flour, meal, grits and flaking grits (lowest). This happens because fumonisins are concentrated in the hull and germ of corn kernel and milling fractions may contain different amounts of these parts. Degermed corn usually contains lower concentration of fumonisins than whole corn.
OCHRATOXIN
Ochratoxin A is a nephrotoxic fungal metabolite produced by certain species of Aspergillus and Penicillium that mainly contaminate cereals like corn, barley, wheat and oats. This mycotoxin, along with another nephrotoxic mycotoxin, citrinin, was implicated in Balkan endemic nephropathy that affected thousands of people in the middle of the 20th century in Eastern Europe. The disease was characterized by anemia, tubular proteinuria and hematuria. This toxin has been shown to be carcinogenic in rats and mice.
Toxicity to ochratoxin A has been reported in pigs after feeding grain with concentrations of this toxin as low as 0.2 ppm. Ochratoxin A is a potent inhibitor of protein synthesis and hence is immunosuppressant.
Its teratogenic and mutagenic potential has also been shown in laboratory animals.
PATULIN
This mycotoxin is produced by Penicillium, Aspergillus and Byssochylamys molds that generally grow on apples. It can occur in significant amounts in apple juice or apple products. Although no major human or animal episodes have been recorded, patulins are toxic in animal feeding studies. The toxin is stable even after pasteurization, cooking or storage.
Its presence has been reported in other food commodities, including bread, legumes, pecans, various fruits (including apricots, pears, grapes, etc.), fruit juices and cheese. It was once considered a potential antibiotic; however, it was abandoned due to lack of effectiveness and possible toxic consequences. It produces gastric irritation, nausea and vomiting upon ingestion. It is uniformly toxic in all mammalian species tested. The biochemical effects of patulin in cells include effects on mitochondrial respiration ultimately causing inhibition of electron transport systems.
Hazards to patulin exposure can easily be avoided by using only tree-picked fruits. Apples or other fruits that are damaged or rotten may not be used for making juice or other food products. The Food and Drug Administration established an action level of 50 µg/kg (50 parts per billion) in apple juice or any other foods that contain apple juice as one of the ingredients. This action level is based on single strength apple juice, one which is not concentrated, or the single strength apple juice component of the food, if the food contains apple juice as an ingredient.
TRICHOTHECENES, INCLUDING DEOXYNIVALENOL (DON)
One of the so-called trichothecenes, deoxynivalenol (DON, also commonly known as vomitoxin), is produced by molds of the genus Fusarium. F. graminearum is a common contaminant of grains including wheat, corn, barley and rye. Another major trichothecene, T-2 toxin, was implicated in the outbreak of alimentary toxic aleukia (ATA) that killed thousands of people in post-World War II Soviet Union. Trichothecene mycotoxins were also involved in the ‘red-mold disease’ intoxication in Japan. Contamination of grains with trichothecenes has been reported in the United States and Canada.
The symptoms of trichothecene toxicity include damage to skin or mucous membranes shortly after contact. Weakness, dizziness and incoordination may ensue shortly after the exposure. These symptoms are followed by bloody diarrhea, difficulty in breathing and bleeding from lungs or mucous membranes after exposure to aerosol containing trichothecenes. In livestock after a low level exposure, loss of appetite and decreased productivity are often reported.
Trichothecenes act by interfering with the ribosomes that are important in protein synthesis. Since protein synthesis is vital for many functions, including that of the immune system, the toxic manifestations of these mycotoxins are of importance in rapidly dividing cells and tissues. Damage to liver, spleen and other lymph nodes is often observed in cases of DON or other trichothecene poisonings.
Mechanisms involved in immunotoxicity
Little information exists on how various mycotoxins produce immunotoxicity. Some mycotoxins, such as aflatoxin Bl and fusarium T-2 toxin, inhibit protein synthesis and cell proliferation (Table 3). This inhibition may not be the primary mechanism involved in their immunotoxic effects; both have selective effects on various subpopulations of lymphocytes.
Several mycotoxins are cytotoxic to lymphocytes in vitro, perhaps because of their effects on membranes (including those involving lymphocytic receptors) or interference with macromolecular synthesis and function.
Cytochalasins (mycotoxins isolated from moldy rice) are highly cytotoxic and act on cytokinesis (perhaps by binding to the filamentous actin), but their immunotoxic potential has not been ascertained.
Table 3. Toxic effects of common mycotoxins and relationship to immune responses. a
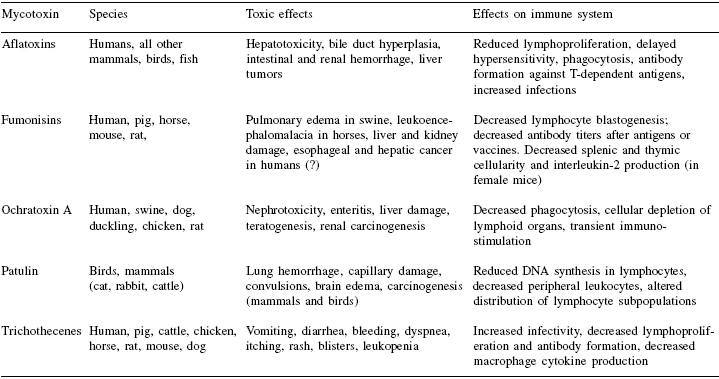
Click to see the image in real size.
aOnly selected symptoms and effects on immunological responses have been indicated. Many mycotoxins are also likely to stimulateproduction of specific antibodies.
Mycotoxins can indirectly influence the immunologic functions. Some of the compounds are neurotoxic or cause other organ pathology, and these compounds may activate the endocrine mechanisms. The stress-induced release of corticosteroids inhibits immune functions. Fusarium T -2 toxin, which acts via such mechanisms, is discussed later.
Some mycotoxins or their metabolites may be highly reactive in mammals and may bind to or destroy tissues. The immune system can also respond to altered proteins or to other biological molecules formed by binding with reactive chemicals, although no experimental evidence exists of this mechanism involving mycotoxins.
Antibodies against mycotoxins conjugated with proteins have been produced and are utilized for analyses for mycotoxins using immunoassays.
The influence of exogenous chemicals on immune responses may be highly variable and a mycotoxin may increase, decrease, or fail to affect the response, depending on the testing protocol and dose.
Impact of mycotoxins on immune responses
AFLATOXINS
Immunomodulation by aflatoxin B1 has been investigated in detail. A comprehensive description may be found elsewhere (Bondy and Pestka, 2000; Sharma, 1991). In most species, resistance to infection is reduced by simultaneous exposure to aflatoxin B1. Effects of aflatoxin B1 are primarily on the cell-mediated immune functions; however, T celldependent humoral responses are also adversely affected (Reddy et al., 1987; Reddy and Sharma, 1989). Generally the humoral responses that are T cell-independent are not affected by low doses of aflatoxins. The immunosuppressive effects of aflatoxin B1 can be explained on the basis of DNA binding of the resulting epoxide derivative of aflatoxin B1, thereby interfering with cell proliferation and protein synthesis.
In humans, the investigations dealing with aflatoxin B1 and impaired immune function are limited. In some parts of the world the exposure to this mycotoxin can be considerably high. Exposure to aflatoxin B1 was reported to be high in Thailand, Swaziland, and Mozambique (45, 43, and 222 ng/kg/day, respectively). In Mozambique the mortality due to hepatoma in hepatoma B virus-antigen carriers was twice that observed in other countries. The actual risk of similar carriers is the same throughout the world. It was proposed that since Mozambique had a high average intake of aflatoxin, the resulting loss of immunoprotection may account for higher viralinduced hepatoma incidence (Lutwick, 1979). A similar increase in primary hepatic carcinoma was suggested in certain provinces of China when people were exposed to high levels of aflatoxin B1 and fumonisin B1 (Ueno et al. 1997); the involvement of immune system in this carcinogenesis is uncertain as both of these mycotoxins are complete carcinogens.
There have been limited studies on the effect of aflatoxin B1 on cytokine production. The expression of macrophage-derived cytokines, namely interleukin (IL)-1α, IL-6 and tumor necrosis factor α (TNFα), was suppressed in mitogen-stimulated macrophages derived from aflatoxin B1 exposed mice (Dugyala and Sharma, 1996). This report suggested that the effect of aflatoxin B1 was greater on macrophages than on other types of immune cells. Macrophages as scavengers may engulf large amounts of aflatoxinbound macromolecules and thereby are selectively sensitive to the toxin.
FUMONISINS
There have been a number of investigations involving fumonisin B1 and immune responses (reviewed by Bondy and Pestka, 2000). Fumonisin B1 generally inhibited lymphocyte blastogenesis in cells obtained from exposed mammals; poultry was relatively resistant. Inconsistent effects of fumonisin on immune functions have been reported. Decreased antibody formation to injected antigens has been reported in turkeys, pigs, calves or even rodents; however, the levels of fumonisins used in these studies have been fairly large.
Much of the studies in laboratory animals and cell lines have been inconclusive with regards to fumonisin-induced immunomodulation. The results depend on the protocol of testing and sequence of exposure to toxin and antigens. In a recent report it was observed that fumonisin B1 altered immune functions in female BALB/c mice, but the males were totally refractory (Johnson and Sharma, 2001). It has been established that fumonisin B1 causes accumulation of free sphingoid bases, sphinganine and sphingosine, by interfering with their conversion to ceramide (Riley et al., 2001) and somehow interrupts the cell cycle (Johnson et al., 2003). The free sphingoid bases and their phosphates are important signaling agents in cells, the bases and their phosphates usually having opposite outcomes for cell survival. Cellular signaling is critical in mounting immune responses; indeed fumonisin B1 has been used as a tool to define the role of ceramide in signaling in immunocompetent cells (lymphocytes). Sphingoid signaling in various physiological processes is becoming relevant and should be further investigated.
The possible involvement of immunological responses in the pathological outcome after fumonisin B1 treatment is discussed later in this report.
OCHRATOXINS
Ochratoxin A, a nephrotoxicant widely encountered as a food contaminant, has been investigated for its
immunologic effects. The biochemical mechanism of ochratoxin poisoning involves interference with macromolecular synthesis, increased lipid peroxidation and diminished mitochondrial respiration. This mycotoxin is fairly cytotoxic and causes atrophy of gastrointestinal lymph nodes after oral ingestion (reviewed by Bondy and Pestka, 2000).
Systemic investigations suggest that exposure to ochratoxin A results in the inhibition of cellular, humoral or innate immune responses; however, many of these effects are produced at exposure levels that are also nephrotoxic. The immunotoxic effects have been demonstrated in poultry, pigs, rats and mice. In some studies immunostimulation, perhaps as a consequence of tissue damage, was also reported.
PATULIN
Patulin has been investigated with regard to the immune system only to a limited extent (Llewellyn et al., 1998). It is less toxic than other mycotoxins; however, effects such as altered number of splenic T lymphocytes, diminished serum immunoglobulin concentrations, decreased delayed hypersensitivity responses and increased neutrophil numbers were reported. The observed changes in cellular phenotype distribution of immune cells may or may not indicate ultimate effects on immune functions. It is suggested that a limited human exposure to foods contaminated with patulin may be of little consequence for the immunologic functions.
TRICHOTHECENE MYCOTOXINS
The prevalent trichothecenes, T-2 toxin, deoxynivalenol (DON), oxynivalenol, and diacetoxyscirpenol, have been evaluated for their immunologic effects to a great extent (reviewed by Sharma and Kim, 1991).
This group of mycotoxins was important for immunological investigations as T-2 toxin was implicated in the onset of alimentary toxic aleukia in the Russian population. Leukopenia is a consistent observation after trichothecene exposures. T-2 toxin is perhaps the only mycotoxin with known immunologic dysfunction in humans.
Exposure to T-2 toxin and deoxynivanol results in severe depletion of T cells. In routine immunotoxicological testing in rodents, both increase and decrease of immunologic functions have been reported depending on the protocol employed. T-2 toxin was immunosuppressive in murine in vivo and in vitro models (Taylor et al., 1985; 1987). Although it is presumed that trichothecene mycotoxins are immunotoxic, the mechanism of their action is not well understood. These mycotoxins are inhibitors of protein synthesis; however, the high susceptibility of T cell function cannot be totally explained on this basis. The T-independent responses are generally less sensitive to T-2 toxin effects. Oral exposure to T-2 toxin in mice results in inflammatory lesions in the forestomach (Taylor et al., 1989), leading to systemic endotoxemia. These effects were associated with increased hypothalamic catecholamine levels and increased peripheral corticosterone concentrations, thereby implicating a possible role of hypothalamicpituitary-adrenal axis in the resulting immunosuppressive responses.
Deoxynivalenol and T-2 toxin have been investigated for their effects on cytokine production by immunocompetent cells. However, effects were not consistent. T-2 toxin caused increased expression and production of IL-2, IL-3 and interferon γ in splenocytes from T-2 treated mice, but a decrease in IL-1α, TNFα and IL-6 was noticed (Dugyala and Sharma, 1997). Deoxynivalenol, however, caused an increase in TNFα production by macrophages (Sugita-Konishi and Pestka, 2001; Yang and Pestka, 2002).
The deoxynivalenol-induced nephropathy in mice can be demonstrated by passive injection of deoxynivalenol-induced IgA monoclonal antibodies (Yan et al., 1998). The findings imply that increased systemic IgA after deoxynivalenol may be implicated in pathogenesis of this mycotoxin in kidney, suggesting the involvement of an immune mechanism in the ultimate pathologic outcome to this mycotoxin.
Immune mechanisms in fumonisin B1-induced murine hepatotoxicity
We have recently investigated the role of immune pathways in the induction of hepatotoxicity in mice after treatment of animals with fumonisin B1.
Systemic treatment of mice with fumonisin B1 caused consistent induction of the pro-inflammatory cytokines, TNFα and interferon γ, in liver (Bhandari and Sharma, 2002; Bhandari et al., 2002b). Indeed the exposure to fumonisin B1 caused localized activation of the cytokine network, implying that innate immune responses are important in the hepatotoxic outcome (Bhandari et al., 2002). The toxicity of fumonisin B1 in murine liver was decreased in mice lacking either tumor necrosis factor receptor (TNFR)-1 or TNFR-2 (Sharma et al., 2000; 2001).
The hepatotoxicity was also ameliorated in mice lacking interferon γ expression (Sharma et al., 2003).
Recently we have also observed that fumonisin B1 hepatotoxicity in mice is totally abolished in NZB/NZW-F1 mice that are spontaneously prone to systemic lupus erythematosis (unpublished observations). The accumulation of free sphingolipid bases was identical in both lupus-prone mice and wild-type controls. The lupus-prone mice are deficient in the production of inflammatory cytokines, including TNFα and interferon γ, thereby further supporting the hypothesis that production of cytokines by innate immune cells in liver is responsible for the ultimate pathological outcome of hepatotoxicity in susceptible animals. Similarly, when mice were injected with anti-thymine antibody (anti-Thy 1.2) together with fumonisin, no hepatotoxicity was observed (unpublished observations).
Miscellaneous mycotoxins in food
There are a number of other mycotoxins that can occur in certain circumstances as food contaminants. Many of these are toxic and also influence the immunologic functions. These include citrinin (a nephrotoxic mycotoxin), rubratoxins (that cause liver and kidney toxicity and diffuse hemorrhage), macrocyclic trichothecenes (a large family of mycotoxins with varied toxicity), secalonic acid (an antineoplastic but teratogenic toxin), cytochalasins (mycotoxins that bind to actin and also inhibit hexose transport in cells), ergotoxins (potent vasoconstrictors), penicillic acid, etc. Few or conflicting reports of these miscellaneous mycotoxins with respect to their effects on immune functions are found in literature. Because of low probability of their occurrence in foods at present, these are not discussed here.
References
Bhandari, N. and R.P. Sharma. 2002. Modulation of selected cell signaling genes in mouse liver by fumonisin B1. Chem-Biol. Interact. 139:317-331.
Bhandari, N., C.C Brown and R.P. Sharma. 2002. Fumonisin B1-induced localized activation of cytokine network in mouse liver. Food Chem. Toxicol. 40:23-31.
Bondy, G.S. and J.J. Pestka. 2000. Immunomodulation by fungal toxins. J. Toxicol. Environ. Health B 3:109-143.
Campbell, B.C., R.J. Molyneux and T.P. Schatzki. 2003. Current research on reducing pre- and postharvest aflatoxin contamination of US almond, pistachio and walnut. J. Toxicol. Toxin Rev. 22:225-266.
Council for Agricultural Science and Technology. 2003. Mycotoxins: risks in plant, animal and human systems. Council for Agricultural Science and Technology, Ames, Iowa, p. 199.
Dugyala, R.R. and R.P. Sharma 1996. The effect of aflatoxin B1 on cytokine mRNA and corresponding protein levels in peritoneal macrophages and splenic lymphocytes. Internat. J. Immunopharmacol. 18:599-608.
Dugyala, R.R. and R.P. Sharma. 1997. Alteration of major cytokines produced by mitogen-activated peritoneal macrophages and splenocytes in T-2- toxin treated male CD-1 mice. Environ. Toxicol. Pharmacol. 3:73-81.
Dutten, M.F. 1996. Fumonisins, mycotoxins of increasing importance: their nature and their effects. Pharmacol. Ther. 70:137-161.
Food and Drug Administration. 2004. Compliance Program Guidance Manual, Chapter 7. Molecular Biology and Natural Toxins. Mycotoxins in domestic foods. p. 4.
Gilbert, J. and E.A. Vargas. 2003. Advances in sampling and analysis for aflatoxins in food and animal feed. J. Toxicol. Toxin Rev. 22:381-422.
Harwig, J., Y.K. Chen, B.P.C. Kennedy and P.M. Scott. 1973. Occurrence of patulin and patulinproducing strains of Penicillium expansum in natural rots of apple in Canada. Can. Inst. Food. Sci. Tech. 6:22-25.
Johnson, V.J. and R.P. Sharma 2001. Genderdependent immunosuppression following subacute exposure to fumonisin B1. Internat. Immunopharmacol. 1:2023-2034.
Johnson, V.J., Q. He, S.H. Kim, A. Kanti and R.P. Sharma. 2003. Increased susceptibility of renal epithelial cells to TNFα-induced apoptosis following treatment with fumonisin B1. Chem.- Biol. Interact. 145:297-309.
Llewellyn, G.C., J.A. McCay, R.D. Brown, D.L. Musgrove, L.F. Butterworth, A.E. Munson and K.L. White, Jr. 1998. Immunological evaluation of the mycotoxin patulin in female B6C3F1 mice. Food Chem Toxicol. 36:1107-1115.
Lutwick, L.I. 1979. Relation between aflatoxin, hepatitis B virus and hepatocellular carcinoma. Lancet 1(8119):755-757.
Osborne, B.G. 1980. The occurrence of ochratoxin A in mouldy bread and flour. Food Cosmet. Toxicol. 18:615-617.
Reddy, R.V. and R.P. Sharma. 1989. Effects of aflatoxin B1 on murine lymphocytic functions. Toxicology 54:31-44.
Reddy, R.V., M.J. Taylor and R.P. Sharma. 1987. Studies of immune function of CD-1 mice exposed to aflatoxin B1. Toxicology 43:123-132.
Riley, R.T., KA. Voss, W.P. Norred, R.P. Sharma, E. Wang and A.H. Merrill. 1998. Fumonisins: mechanisms of mycotoxicity. Revue Med. Vet. 149, 617-626.
Riley, R.T., E. Enongene, K.A. Voss, W.P. Norred, F.I. Meredith, R.P. Sharma, D. Williams, J. Spitsbergen, D.E. Williams, D.B. Carlson and A.H. Merrill. 2001. Sphingolipid perturbations as mechanisms for fumonisin carcinogenesis. Environ. Health Perspect. 109(Supp. 2):301-308.
Sharma, R.P. 1985. Immunotoxicology of food constituents. Food Technol. 39:94-105.
Sharma, R.P. 1991. Immunotoxic effects of mycotoxins. In: Mycotoxins and Phytoalexins (R.P. Sharma and D.K. Salunkhe, eds). CRC Press, Boca Raton, pp. 81-99.
Sharma, R.P. 1993. Immunotoxicity of mycotoxins. J. Dairy Sci. 76:892-897.
Sharma, R.P. and Y.W. Kim. 1991. Trichothecenes. In: Mycotoxins and Phytoalexins (R.P. Sharma and D.K. Salunkhe, eds). CRC Press, Boca Raton, pp. 339-359.
Sharma, R.P., N. Bhandari, R.T. Riley, KA. Voss and F.I. Meredith. 2000. Tolerance to fumonisin toxicity in a mouse strain lacking the P75 tumor necrosis factor receptor. Toxicology 143:183-194.
Sharma, R.P., N. Bhandari, Q. He, R.T. Riley and K.A. Voss. 2001. Decreased fumonisin hepatotoxicity in mice with a targeted deletion of tumor necrosis factor receptor 1. Toxicology 159:69-79.
Sharma, R.P., Q. He and V.J. Johnson. 2003. Deletion of interferon γ reduces fumonisin-induced hepatotoxicity in mice via alterations in inflammatory cytokines and apoptotic factors. J. Interferon Cytokine Res. 23:13-23.
Shepard, G.S. 2003. Aflatoxin and food safety: recent African perspectives. J. Toxicol. Toxin Rev. 22:267-286.
Shotwell, O.L., C.W. Hesseltine, E.E. Vandegraft and M.L. Goulden. 1971. Survey of corn from different regions for aflatoxin, ochratoxin and zearalenone. Cereal Sci. Today 16:266-273.
Sugita-Konishi, Y. and J.J. Pestka. 2001. Differential upregulation of TNF-α, IL-6, and IL-8 production by deoxynivalenol (vomitoxin) and other 8-ketotrichothecenes in a human macrophage model. J. Toxicol. Environ. Health 64:619-636.
Taylor, M.J., R.V. Reddy and R.P. Sharma. 1985. Immunotoxicity of repeated low level exposure to T-2 toxin, a trichothecene mycotoxin, in CD-1 mice. Mycotox. Res. 1:57-64.
Taylor, M.J., B.J. Hughes, and R.P. Sharma. 1987. Dose and time related effects of T-2 toxin on mitogenic response of murine splenic cells in vitro. Int. J. Immunopharmacol. 9:107-113.
Taylor, M.J., R.A. Smart and R.P. Sharma. 1989. Relationship of the hypothalamic-pituitary-adrenal axis with chemically-induced immunomodulation. I. Stress-like response after exposure to T-2 toxin. Toxicology 179-195.
Ueno, Y., K. Lijima, S.D. Wang, Y. Sugiura, M. Sekijima, T. Tanaka, C. Chen, S.Z. Yu. 1997. Fumonisins as a possible risk contributory factor for primary liver cancer: a 3-year study of corn harvested in Haimen, China, by HPLC and ELISA. Food Chem. Toxicol. 35:1143-1150.
Wilson, D.M. and G.J. Nuovo. 1973. Patulin production in apples decayed by Penicillium expansum. Appl. Microbiol. 26:124-125.
Yan, D., W.K. Rumbeiha and J.J. Pestka. 1998. Experimental murine IgA nephropathy following passive administration of vomitoxin-induced IgA monoclonal antibodies. Food Chem. Toxicol. 36:1095-1106.
Yang, G.H. and J.J. Pestka. 2002. Vomitoxin (deoxynivalenol)-mediated inhibition of nuclear protein binding to NRE-A, an IL-2 promoter negative regulatory element, in EL-4 cells. Toxicology 172:169-179.
Author: RAGHUBIR P. SHARMA
Department of Physiology and Pharmacology, The University of Georgia, Athens, Georgia, USA
Related topics:
Recommend
Comment
Share

5 de junio de 2008
Although mold fungus are considered as poor immune provoker, still they suppress immune system in long time. If the immune system of animals is stimulated animals can withstand the effect of mycotoxin.
Recommend
Reply

Would you like to discuss another topic? Create a new post to engage with experts in the community.





Featured users in Mycotoxins

