mycotoxins on the body’s antioxidant defence
Impact of mycotoxins on the body’s antioxidant defence
Among all mycotoxins, those from Fusarium species are considered to be important contaminants of poultry feed. Trichothecenes, zearalenone, fumonisins, moniliformin and fusaric acid are the major Fusarium mycotoxins occurring on a worldwide basis in cereal grains, animal feeds and forages (D’Mello et al., 1999; Figure 1).
Furthermore, the trichothecene mycotoxins themselves comprise a vast group of over 100 fungal compounds with the same basic structure (Leeson et al., 1995).
Acute mycotoxicosis outbreaks are rare events in modern poultry production. However, low levels of mycotoxin contamination, which very often are not detected, are responsible for reduced efficiency of production and increased susceptibility to infectious disease. These sometimes undetectable or unseen compounds can cost poultry producers a fortune, unless a radical solution for the problem is found. The problem is further complicated since in many cases molecular mechanisms of their action have not been fully elucidated.

Figure 1. Diverse structure of some common mycotoxins.
Biochemical changes in mycotoxicosis vary greatly and lipid peroxidation is regarded as one of the most important consequences of mycotoxicosis (Mezes et al., 1999). Aflatoxin B1 (AFB1), T-2 toxin, ochratoxin A (OA), fumonisin B1 (FB1), zearalenone (ZEN) and deoxynivalenol (DON) are the most notorious and extensively studied mycotoxins (Galvano et al., 2001) and all of them are involved in lipid peroxidation.
Compromised antioxidant defences due to mycotoxicoses
A delicate balance between antioxidants and prooxidants in the body in general and specifically in the cell is responsible for regulation of various metabolic pathways leading to maintenance of immunocompetence, growth and development and protection against stress conditions associated with commercial poultry production (Surai and Dvorska, 2001). This balance can be regulated by dietary antioxidants, including vitamin E (Surai et al., 1999), carotenoids (Surai and Speake, 1998; Surai et al., 2001) and selenium (Se) (Surai, 2000). On the other hand, nutritional stress factors have a negative impact on this antioxidant/pro-oxidant balance. In this respect, mycotoxins can be considered among the most important feed-borne stress factors.
It is not clear at present whether mycotoxins stimulate lipid peroxidation directly by enhancing free radical production or if the increased tissue susceptibility to lipid peroxidation is a result of compromising antioxidant defence. It seems likely that both processes are involved. In most cases lipid peroxidation in tissues caused by mycotoxins was associated with decreased concentrations of natural antioxidants. For example, in an experiment with quail, levels of the primary liver antioxidants (α- tocopherol, γ-tocopherol, carotenoids and ascorbic acid) significantly decreased as a result of T-2 toxin consumption (Dvorska and Surai, 2001; Figure 2).
Similarly, the presence of T-2 toxin in the diet decreased liver concentration of α-tocopherol in chickens (Hoehler and Marquardt, 1996). T-2 toxin consistently depressed concentrations of vitamin E in chicken plasma (Coffin and Combs, 1981).
Similarly, AFB1 in the feed interfered with the accumulation of carotenoids in chicken tissues (Schaeffer et al., 1988), inducing pale bird syndrome. In fact, AFB1 caused a significant depression of lutein in the toe web, liver, serum and mucosa (Schaeffer et al., 1988). Pigment restoration was accomplished by feeding the same diet supplemented with lutein (70 mg/kg). In young chickens AFB1 reduced the lutein content of the jejunal mucosa up to 35% and serum lutein was reduced up to 70% (Tyczkowski and Hamilton, 1987a), suggesting that AFB1 interfered with the absorbtion, transport and deposition of carotenoids.
More precisely, AFB1 impaired lutein absorption in chickens (Tyczkowski and Hamilton, 1987b). In similar fashion, OA was shown to affect carotenoid assimilation in chickens. Again, depression in uptake of carotenoids by intestinal mucosa and depressed transport in serum were considered important mechanisms of AFB1 action on carotenoid metabolism (Schaeffer et al., 1987; Huff and Hamilton, 1975).

Figure 2. Effect of T-2 toxin and absorbents on antioxidants concentrations in quail liver (from Dvorska and Surai, 2001).

Figure 3. Effect of T-2 toxin and sorbents on vitamin A (retinol and retinol esters) concentrations in quail liver (from Dvorska and Surai, 2001).
In general, malabsorption syndrome is considered a common result of mycotoxicoses. For example, aflatoxicosis, ochratoxicosis and T-2 toxicosis were produced by feeding diets containing graded concentrations of the appropriate toxin to broiler chickens from hatching until 3 weeks of age (Osborne et al., 1982). In this experiment AFB1, levels lower than those needed for growth inhibition produced a malabsorption syndrome characterized by steatorrhea, hypocarotenoidemia, and decreased concentrations of bile salts and pancreatic enzymes.
T-2 toxin also produced malabsorption, but at concentrations higher than required to inhibit growth. Ochratoxicosis produced mainly hypocarotenoidemia (Osborne et al., 1982). It is postulated that the decreased level of vitamin A in the quail liver as a result of T-2 toxin consumption (Dvorska and Surai, 2001; Figure 3) is also a reflection of the decreased intestinal absorption of fat soluble nutrients.
Molecular mechanisms of malabsorption development due to mycotoxins need further investigation. However, it can be suggested that mycotoxins may stimulate lipid peroxidation in enterocytes leading to damage that could substantially contribute to malabsorption development.
The presence of ochratoxin A in the diet significantly decreased the concentration of α- tocopherol in the chicken liver (Hoehler and Marquardt, 1996). Furthermore, aflatoxin-treated barrows had decreased serum tocopherol and retinol concentrations compared with control and pre-test values, and decreased tocopherol concentration in cardiac tissue (Harvey et al., 1994).
A pro-oxidant effect of mycotoxins in many cases could be mediated via changes in reduced glutathione concentration. For example, Rizzo et al. (1994) demonstrated that T-2 toxin decreased GSH content in rat liver. Treatment of fasted mice with a single dose of T-2 toxin (1.8 or 2.8 mg/kg body weight) by oral gavage led to marked decrease in hepatic GSH levels (Atroshi et al., 1997). In male broiler chicks, hepatic GSH concentration decreased after 7 days of treatment (1.5 mg T-2 toxin/kg body weight/day) (Leal et al., 1999). Acute exposure of mice to T-2 toxin (4 mg/kg,) resulted in a progressive decrease in hepatic glutathione content, reaching a minimum 6-8 hrs after toxin administration (Fricke and Jorge, 1991).
Intraperitoneal administration of AFB1 to rats (2 mg/kg) was also associated with decreased GSH in the liver. In contrast, in 3-week-old male chickens daily aflatoxin gavage (2 mg/kg body wt, in corn oil) for 5 and 10 days elevated plasma hepatic GSH; and renal GSH was elevated after 10 days of aflatoxin treatment (Beers et al., 1992a). Similarly, hepatic GSH increased 2 and 8 hrs following a single AFB1 dose and continued to increase through five daily doses of AFB1 (Beers et al., 1992b).
There was GSH depletion in cultured rat hepatocytes as a result of AFB1 toxicosis (Liu et al., 1999). Similarly, consumption of OA for two weeks was associated with a depletion of GSH from the mouse liver (Atroshi et al., 2000). The mycotoxin patulin also decreased GSH concentration in rat hepatocytes (Busbee et al., 1999). Since glutathione is responsible for the maintenance of cellular redox status (Sies, 1999) and therefore participates in regulation of gene expression (Arrigo, 1999), changes in GSH status could be detrimental.
One of the most important mycotoxin actions is their effect on antioxidant enzymes. Depending on experimental conditions (species, dose, route and duration of exposure, concentrations of other antioxidants etc.), antioxidant enzyme activities can increase in response to oxidative stress or decrease by direct or indirect action of mycotoxins. For example, treatment of pig kidney cells with 50 μM FB1 for 24 hrs significantly decreased cellular GSH and increased the activity of glutathione reductase (Kang and Alexander, 1996). The activities of GSHPx, catalase and Cu/Zn-SOD were not changed by this treatment. Oral administration of T-2 toxin to rats (1.25 mg/kg) for five days decreased activity of liver glutathione-S-transferase (Ahmed and Ram, 1986). In contrast, feeding a single dose of T-2 toxin (2 mg/kg body weight) to rats was associated with increased activities of GSH-shuttle enzymes including GSH-Px, glutathione reductase and glucose-6-phosphate dehydrogenase (Suneja et al., 1989), probably reflecting an adaptive response to oxidative stress. On the other hand, when male rats were fed a diet deficient in vitamins C and E and Se were orally administered a single dose of DON or T-2 toxin, there was a significant decrease in activities of GSH-Px, catalase, SOD and glutathione reductase (Rizzo et al., 1994). Activity of GSH-Px in rat blood was decreased due to consumption of AFB1 (Choi et al., 1995). Administration of AFB1 to rats (2 mg/kg intraperitoneally) decreased the activities of SOD, catalase, GSH-Px, glutathione- S-transferase and glutathione reductase in liver (Rastogi et al., 2001a). A significant increase in liver SOD activity occurred following AFB1 exposure of ducks (Barraud et al., 2001).
One of the most important targets for mycotoxins is embryonic development. Since chicken embryo tissues contain high levels of PUFA, they are vulnerable to peroxidation, and oxidative stress caused by mycotoxins could be lethal. For example, aurofusarin increased late mortality of quail embryos (Dvorska et al., 2001). Furthermore, contamination of the diet with T-2 toxin markedly decreased egg production and impaired hatchability (Tobias et al., 1992). Confirmation of a possible association of this effect with oxidative stress came from data indicating that increased dietary vitamin E concentration during the first week of the experiment significantly decreased the number of infertile eggs and significantly improved the hatching percentage (Tobias et al., 1992).
Increased lipid peroxidation as a consequence of mycotoxicoses
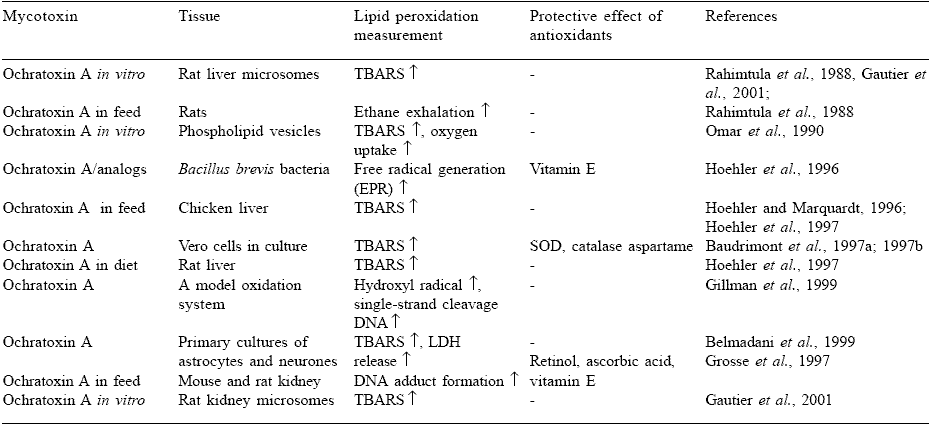
As illustrated in Table 1, OA has a stimulating effect on lipid peroxidation. In most cases, thiobarbituric acid reactive substances (TBARS) accumulation was used as a measurement of lipid peroxidation. Furthermore, ethane exhalation, EPR-registered free radicals, hydroxyl radical formation, singlestrand cleavage of DNA, DNA adduct formation as well as LDH release were also used to confirm pro-oxidant properties of OA. Various in vitro and in vivo systems were also used including liver microsomes, phospholipid vesicles, primary cell cultures, whole organs and whole body.
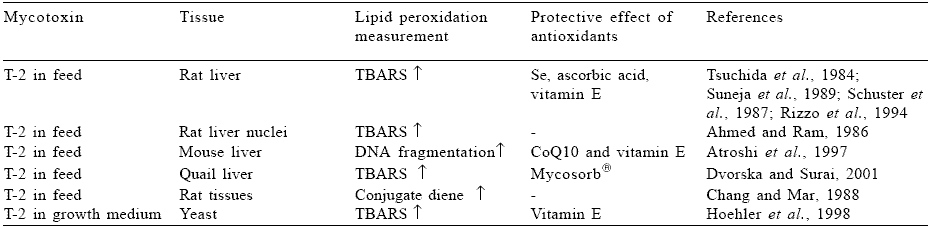
T-2 toxin was also shown to have pro-oxidant properties (Table 2). Those properties were confirmed with rat, mouse and quail liver tissue and yeast. TBARS accumulation was a method of choice for most of the studies, however, data on conjugate diene formation and DNA fragmentation also showed those effects. Effect of AFB1 on lipid peroxidation has been studied in rat liver and kidney as well as in cultured hepatocytes and in an in vitro model system (Table 3). Similar to the examples above, TBARS accumulation was substantially increased as well as conjugate diene production. At the same time GSH concentration and activities of antioxidant enzymes significantly declined as a result of AFB1 action.
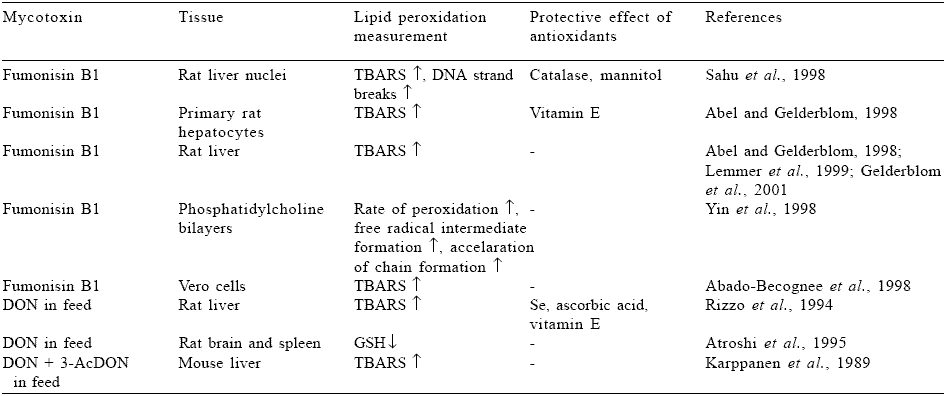
FB1 also stimulated lipid peroxidation in rat liver, rat liver nuclei fraction, primary rat hepatocytes, Vero cells in culture and PC bilayers. In those systems TBARS accumulation and DNA strand breaks were increased (Table 4). DON increased TBARS formation in rat and mice liver and decreased GSH in rat brain and spleen. There are also data available indicating pro-oxidant properties of zearalenone (Karagezyan et al., 1995; Ghedira- Chekir et al., 1999) and citrinin (Ribeiro et al., 1997).
Table 1. Stimulation of lipid peroxidation by ochratoxin A.
To enlarge the image, click here
Table 2. Effect of T-2 toxin on lipid peroxidation.
To enlarge the image, click here
It is clear from these data that mycotoxins strongly promote lipid peroxidation in various in vitro and in vivo systems. This effect was obvious no matter which measurement was used to assess the process of lipid peroxidation.
Protection against pro-oxidant effects of mycotoxins
The range of mycotoxins that can contaminate poultry feed and their different chemical structures make protection against mycotoxin-related toxicity a difficult task. There are various approaches to control or combat mycotoxin problems. The simplest strategy is based on the prevention of the formation of mycotoxins in feeds by special management programmes including storage at low moisture levels and prevention of grain damage during processing (Dawson, 2001). However, modern agronomic technology is not able to eliminate pre-harvest infection of susceptible crops by fungi (Wood, 1992). Therefore this strategy can only partially be effective; and in countries with warm and humid conditions, this strategy could be quite costly.
Other strategies based on microbial or thermal inactivation of toxins, physical separation of contaminated feedstuffs, irradiation, ammoniation and ozone degradation have not proved practical because they are either time-consuming or comparatively expensive (Dawson, 2001). In recent years, nutritional manipulation has been actively used to improve animal self-defence against mycotoxins or to decrease detrimental consequences of mycotoxin consumption.
Table 3. Stimulation of lipid peroxidation in rats by Aflatoxin B1.

To enlarge the image, click here
*IP = intraperitoneally
Since lipid peroxidation plays an important role in mycotoxin toxicity, a protective effect of antioxidants is expected (Galvano et al., 2001). Indeed, as can be seen from data presented in Tables 1-4, protective effects against lipid peroxidation caused by mycotoxins were attributed to various antioxidant compounds including vitamins A and E, ascorbic acid, CoQ10, selenium, antioxidant enzymes as well as various plant extracts.
In spite of positive effects of natural antioxidants on animals fed mycotoxin-contaminated diets, the most promising and practical approach has been the addition of adsorbents to contaminated feed (Ledoux and Rottinghaus, 2000). Mycotoxins can be bound to the adsorbent and pass harmlessly through the digestive tract. Many compounds have been tested for adsorbent effects, however comparatively few have proven successful and still fewer (mainly bentonites, zeolites and alumosilicates) are used commercially (Devegowda et al., 1998).
The extent to which various compounds bind specific toxins varies considerably. Many products only bind aflatoxin, leaving such mycotoxins as T-2 in the intestinal tract without alteration. In addition to the various clays and zeolites, a yeast cell wallderived glucomannan (Mycosorb ®) has been shown to be effective against a wide range of mycotoxins (Devegowda et al., 1998).
Mycotoxin binders can substantially improve the status of antioxidant systems in animals. This effect depends on the mycotoxin-binding activity of adsorbents. For example, inclusion of zeolite in the quail diet at 3% had a minor protective effect on the antioxidants in the quail liver and changes were not statistically significant (Figure 2; Dvorska and Surai, 2001a). Only the concentration of retinyllinoleate in the liver of quail exposed to T-2 toxin simultaneously with zeolite was significantly higher compared with birds fed the diet containing T-2 toxin alone. These data indicate that zeolites alone were not effective in prevention of T-2 toxicity. These data are in agreement with observations of Kubena et al. (1990; 1998) indicating absence of protective effects of aluminosilicate sorbents against T-2 toxicosis. Superactivated charcoal (Edrington et al., 1997) and inorganic sorbents (Bailey et al., 1998) were also ineffective against T-2 toxicosis. Therefore, zeolite was probably unable to bind a substantial amount of T-2 toxin in the digestive tract; and as a result did not interfere with pro-oxidant properties of this mycotoxin.
In marked contrast, inclusion of yeast
glucomannans (Mycosorb®) in T-2 toxin-containing diets fed quail significantly slowed the depletion of natural antioxidants and vitamin A in the liver (Figure 3). This protective effect can be attributed to the high adsorbent capability that esterified glucomannans have for T-2 (Dawson, 2001). It could well be that mycotoxin binding by Mycosorb® also prevents T-2 toxin participation in development of oxidative stress in the intestine. As a result, damage to the enterocytes is prevented thereby maintaining effective antioxidant absorption, assimilation and delivery to the target tissues.
Table 4. Effect of Fumonisin B1 and DON on lipid peroxidation.
To enlarge the image, click here
Due to antioxidant depletion in the liver, susceptibility to lipid peroxidation increased more than 2-fold (Figure 4). Inclusion of zeolites in the diet did not prevent antioxidant depletion; and therefore susceptibility to lipid peroxidation in the liver was increased, showing no significant difference from the group fed the T-2 toxin treatment without an adsorbent additive. On the other hand, inclusion of Mycosorb® in the T-2-contaminated diet significantly decreased tissue susceptibility to lipid peroxidation in comparison to diets containing toxin only, although the inclusion of the Mycosorb® adsorbent material was unable to completely mitigate the powerful stimulating effect of T-2 toxin on lipid peroxidation.

Figure 4. Lipid peroxidation in the quail liver. (Adapted from Dvorska and Surai, 2001).
Inclusion of Mycosorb® in the quail diet was unable to completely prevent the adverse effects of T-2 toxin on the antioxidant systems of the liver of the growing quail; indicating that not all T-2 toxin was bound and released from the intestine. Therefore, a combination of mycotoxin binders with natural antioxidants, in particular with Se and vitamin E, could be the next step in preventing damaging effects of mycotoxins.
Conclusions
Stimulation of lipid peroxidation and consequent apoptosis are important mechanisms of the toxicity of various mycotoxins (Surai, 2002). Therefore, various natural antioxidants have been successfully tested as protective agents. However, using antioxidants alone it is impossible to prevent detrimental effects of mycotoxins on various metabolic processes in the body. Mycotoxin binders show promise in decreasing mycotoxin toxicity as well as in preventing damage to antioxidant systems. It is most likely that a combination of mycotoxin binders such as Mycosorb® with natural antioxidants such as organic selenium or vitamin E would be an effective approach to combat mycotoxicoses in future.
References
Authors: PETER F. SURAI1, JULIA E. DVORSKA2, NICK H.C. SPARKS1 and KATE A. JACQUES3Abado-Becognee, K., T.A. Mobio, R. Ennamany, F. Fleurat-Lessard, W.T. Shier, F. Badria and E.E. Creppy. 1998. Cytotoxicity of fumonisin B1: implication of lipid peroxidation and inhibition of protein and DNA syntheses. Archives of Toxic. 72:233-236.
Abel, S. and W.C. Gelderblom. 1998. Oxidative damage and fumonisin B1-induced toxicity in primary rat hepatocytes and rat liver in vivo. Toxicology 131:121-131.
Ahmed, N. and G.C. Ram. 1986. Nuclear lipid peroxidation induced in rat liver by T-2 mycotoxin. Toxicol. 24:947-949.
Arrigo, A.P. 1999. Gene expression and the thiol redox state. Free Radical Bio. and Med. 27:936- 944.
Atroshi F., A. Rizzo, I. Biese, M. Salonen, L.A. Lindberg and H. Saloniemi. 1995. Effects of feeding T-2 toxin and deoxynivalenol on DNA and GSH contents of brain and spleen of rats supplemented with vitamin E and C and selenium combination. J. Anim. Physiol. Anim. Nutr. 74:157-164.
Atroshi, F., A. Rizzo, I. Biese, P. Veijalainen, E. Antila and T. Westermarck. 1997. T-2 toxininduced DNA damage in mouse livers: the effect of pretreatment with coenzyme Q10 and alphatocopherol. Molecular Aspects of Med. 18 Suppl:S255-S2588.
Atroshi, F., I. Biese, H. Saloniemi, T. Ali-Vehmas, S. Saari, A. Rizzo and P. Veijalainen. 2000. Significance of apoptosis and its relationship to antioxidants after ochratoxin A administration in mice. J. of Phar. and Pharm. Sci. 3:281-291.
Barraud, L., T. Douki, S. Guerret, M. Chevallier, C. Jamard, C. Trepo, C.P. Wild, J. Cadet and L. Cova. 2001. The role of duck hepatitis B virus and aflatoxin B1 in the induction of oxidative stress in the liver. Cancer Det. and Prev. 25:192-201.
Bailey, R.H., L.F. Kubena, R.B. Harvey, S.A. Buckley and G.E. Rottinghaus. 1998. Efficacy of various inorganic sorbents to reduce the toxicity of aflatoxin and T-2 toxin in broiler chickens. Poultry Sci. 77:1623-1630.
Baudrimont, I., R. Ahouandjivo and E.E. Creppy. 1997a. Prevention of lipid peroxidation induced by ochratoxin A in Vero cells in culture by several agents. Chemico-Bio. Inter. 104:29-40.
Baudrimont, I., A.M. Betbeder and E.E. Creppy. 1997b. Reduction of the ochratoxin A-induced cytotoxicity in Vero cells by aspartame. Archives of Toxicol. 71:290-298.
Beers, K.W., H. Nejad and W.G. Bottje. 1992a. Aflatoxin and glutathione in domestic fowl (Gallus domesticus). I. Glutathione elevation and attenuation by high dietary methionine. Comp. Bio. and Phy. 101C:239-244.
Beers, K.W., R.P. Glahn, W.G. Bottje and W.E. Huff. 1992b. Aflatoxin and glutathione in domestic fowl (Gallus domesticus). II. Effects on hepatic blood flow. Comp. Bio. and Phy. 101C:463-467.
Belmadani, A., P.S. Steyn, G. Tramu, A.M. Betbeder, I. Baudrimont and E.E. Creppy. 1999. Selective toxicity of ochratoxin A in primary cultures from different brain regions. Archives of Toxicol. 73:108-114.
Busbee, D., R. Barhoumi, R.C. Burghardt, C. Gauntt, B. McAnalley and H.R. McDaniel. 1999. Protection from glutathione depletion by a glyconutritional mixture of saccharides. Age 22:159-165.
Chang, I.M. and W.C. Mar. 1988. Effect of T-2 toxin on lipid peroxidation in rats: elevation of conjugated diene formation. Toxic. Letters 40:275-280.
Choi, Y.K., K.K. Jung, K.Y. Chae, I. Jang, B.D. Lee and K.H. Nahm. 1995. Effects of vitamin E and selenium supplementation to diets containing aflatoxin B 1 on the contents of liver lipids and various blood parameters in rats. Asian-Austr. J. of Anim. Sci. 8:379-385.
Coffin, J.L. and G.F. Combs, Jr. 1981. Impaired vitamin E status of chicks fed T-2 toxin. Poultry Sci. 60:385-392.
Dawson, K.A. 2001. Understanding the absorption characteristics of yeast cell wall preparations associated with mycotoxin binding. In: Biotechnology in the Feed Industry: Proceedings of Alltech’s 17th Annual Symposium (T.P. Lyons and K.A. Jacques, eds), Nottingham University Press, Nottingham, UK, pp. 169-181.
Devegowda, G., M.V.L.N. Raju and H.V.L.N. Swamy. 1998. Mycotoxins: novel solutions for their counteraction. Feedstuffs 70:12-15, December 7.
D’Mello, J.P.F., C.M. Placinta and A.M.C. MacDonald. 1999. Fusarium mycotoxins: a review of global implications for animal health, welfare and productivity. Anim. Feed Sci. and Tech. 80:183-205.
Dvorska, J.E. and P.F. Surai. 2001a. Effects of T-2 toxin, zeolite and Mycosorb on antioxidant systems of growing quail. Asian-Aust. J. of Anim. Sci. 14:1-6.
Dvorska, J.E. and P.F. Surai. 2001b. Stimulating effect of aflatoxin B1 on lipid peroxidation in the in vitro model systems. Scientific programme and abstracts of 6th International Symposium on Poisonous Plants, Glasgow, UK, p. 21.
Dvorska, J.E., P.F. Surai and N.H.C. Sparks. 2001. Mycotoxin aurofusarin: effect on quail. Scientific programme and abstracts of 6th International Symposium on Poisonous plants, Glasgow, UK, p. 86.
Edrington, T.S., L.F. Kubena, R.B. Harve and G.E. Rottinghaus. 1997. Influence of a superactivated charcoal on the toxic effects of aflatoxin or T-2 toxin in growing broilers. Poultry Sci. 76:1205- 1211.
Fink-Gremmels, J. 1999. Mycotoxins: their implications for human and animal health. Vet. Quatery 21:115-120.
Fricke, R.F. and J. Jorge. 1991. Methylthiazolidine- 4-carboxylate for treatment of acute T-2 toxin exposure. J. of Applied Toxic. 11:135-140.
Galvano, F., A. Piva, A. Ritieni and G. Galvano. 2001. Dietary strategies to counteract the effects of mycotoxins: a review. J. of Food Prot. 64:120- 131.
Gautier, J.C., D. Holzhaeuser, J. Markovic, E. Gremaud, B. Schilter and R.J. Turesky. 2001. Oxidative damage and stress response from ochratoxin A exposure in rats. Free Radical Bio. and Med. 30:1089-1098.
Gelderblom W.C., D. Galendo, S. Abel, S. Swanevelder, W.F. Marasas, C.P. Wild. 2001. Cancer initiation by fumonisin B(1) in rat liverrole of cell proliferation. Cancer Letter 169:127-137.
Ghedira-Chekir, L., K. Maaroufi, E.E. Creppy and H. Bacha. 1999. Cytotoxic and genotoxic effects of zearalenone: Prevention by vitamin E. J. of Toxicology –Toxin reviews 18:355-368.
Gillman, I.G., T.N. Clark and R.A. Manderville. 1999. Oxidation of ochratoxin A by an Feporphyrin system: model for enzymatic activation and DNA cleavage. Chem. Res. in Toxic. 12:1066- 1076.
Grosse, Y., L. Chekir-Ghedira, A. Huc, S. Obrecht- Pflumio, G. Dirheimer, H. Bacha and A. Pfohl-Leszkowicz. 1997. Retinol, ascorbic acid and alpha-tocopherol prevent DNA adduct formation in mice treated with the mycotoxins ochratoxin A and zearalenone. Cancer Letters 114:225-229.
Ha T.G., N.D. Kim. 1998. 2-(allylthio)pyrazine inhibition of aflatoxin B1-induced hepatotoxicity in rats: inhibition of cytochrome P450 2B- and 3A2-mediated bioactivation. Res Commun Mol Pathol Pharmacol. 102:69-78.
Harvey, R.B., L.F. Kubena and M.H. Elissalde. 1994. Influence of vitamin E on aflatoxicosis in growing swine. Amer. J. of Vet. Res. 55:572-577.
Hoehler, D. and R.R. Marquardt. 1996. Influence of vitamins E and C on the toxic effects of ochratoxin A and T-2 toxin in chicks. Poultry Sci. 75:1508-1515.
Hoehler, D., R.R. Marquardt, A.R. McIntosh and M. Srinivasa. 1998. Free radical mediated lipid peroxidation induced by T-2 toxin in yeast (kluyveromyces marxianus). J. Nutr. Biochem. 9:370-379.
Hoehler, D., R.R. Marquardt, A.R. McIntosh and H. Xiao. 1996. Free radical generation as induced by ochratoxin A and its analogs in bacteria (Bacillus brevis ). J. of Bio. Chem. 271:27388- 27394.
Hoehler, D., R.R. Marquardt and A.A. Frohlich. 1997. Lipid peroxidation as one mode of action in ochratoxin A toxicity in rats and chicks. Canadian J. of Anim. Sci. 77:287-292.
Huff, W.E. and P.B Hamilton. 1975. Decreased plasma carotenoids during ochratoxicosis. Poultry Sci. 54:1308-1310.
Kang, Y.J. and J.M. Alexander. 1996. Alterations of the glutathione redox cycle status in fumonisin B1-treated pig kidney cells. J. of Bio. Toxic. 11:121-126.
Karagezyan, M.K., L.M. Ovsepyan, S.S. Ovakimyan, A.S. Boyadzhyan, L.L. Osipyan and K.G. Karagezyan. 1995. Mycotoxin zearalenone is an activator of the free-radical lipid peroxidation process in the mitochondrial and microsomal fractions of white rat liver. Doklady, Biochemistry 341:45-47.
Karppanen, E., A. Rizzo, L. Saari, S. Berg and H. Boström. 1989. Investigation on trichothecenestimulated lipid peroxidation and toxic effects of trichothecenes in animals. Acta Vet. Scandinavica 30:391-399.
Kubena, L.F., R.B. Harvey, W.E. Huff, D.E. Corrier, T.D. Philips and G.E. Rottinghaus. 1989. Influence of ochratoxin A and T-2 toxin singly and in combination on broiler chickens. Poultry Sci. 68:867-872.
Kubena, L.F., R.B. Harvey, W.E. Huff, D.E. Corrier, T.D. Philips and G.E. Rottinghaus. 1990. Efficacy of a hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2 toxin. Poultry Sci. 69:1078-1086.
Kubena, L.F., R.B. Harvey, T.H. Bailey, S.A. Buckley and G.E. Rottinghaus. 1998. Effects of a hydrated sodium calcium aluminosilicate (TBind) on mycotoxicosis in young broiler chickens. Poultry Sci. 77:1502-1509.
Leal, M., A. Shimada, F. Ruiz, E. Gonzalez de Mejia. 1999. Effect of lycopene on lipid peroxidation and glutathione-dependent enzymes induced by T-2 toxin in vivo. Toxicology Letters 109:1-10.
Ledoux, D.R. and G.E. Rottinghaus. 2000. Animal model for testing adsorbents to detoxify mycotoxins. Feed Mix 8:18-20.
Leeson, S., G. Diaz and J.D. Summers. 1995. Poultry metabolic disorders and mycotoxins. University Books. Guelph, Canada.
Lemmer, E.R., W.C.A. Gelderblom, E.G. Shephard, S. Abel, B.L. Seymour, J.P. Cruse, R.E. Kirsch, W.F.O. Marasas and P. de la M. Hall. 1999. The effects of dietary iron overload on fumonisin B1- induced cancer promotion in the rat liver. Cancer Letters 146:207-215.
Liu, J., C.F. Yang, B.L. Lee, H.M. Shen, S.G. Ang and C.N. Ong. 1999. Effect of Salvia miltiorrhiza on aflatoxin B1-induced oxidative stress in cultured rat hepatocytes. Free Radical Res. 31:559-568.
Meki A.R., S.K. Abdel-Ghaffar and I. El-Gibaly. 2001. Aflatoxin B1 induces apoptosis in rat liver: protective effect of melatonin. Neuroendocrinol. Letter 22:417-426.
Mezes, M., M. Barta and G. Nagy. 1999. Comparative investigation on the effect of T-2 mycotoxin on lipid peroxidation and antioxidant status in different poultry species. Res. in Vet. Sci. 66:19-23.
Omar, R.F., B.B. Hasinoff, F. Mejilla and A.D. Rahimtula. 1990. Mechanism of ochratoxin A stimulated lipid peroxidation. Bio. Pharm. 40:1183- 1191.
Osborne, D.J., W.E. Huff, P.B. Hamilton and H.R. Burmeister. 1982. Comparison of ochratoxin, aflatoxin, and T-2 toxin for their effects on selected parameters related to digestion and evidence for specific metabolism of carotenoids in chickens. Poultry Sci. 61:1646-1652.
Premalatha, B., V. Sujatha and P. Sachdanandam. 1997. Modulating effect of Semecarpus anacardium Linn. nut extract on glucose metabolizing enzymes in aflatoxin B1-induced experimental hepatocellular carcinoma. Pharmacol Res. 36:187-192.
Rahimtula, A.D., J.C. Bereziat, V. Bussacchini-Griot and H. Bartsch. 1988. Lipid peroxidation as a possible cause of ochratoxin A toxicity. Bio. Pharm. 37:4469-4477.
Rastogi, R., A.K. Srivastava and A.K. Rastogi. 2001a. Long term effect of aflatoxin B(1) on lipid peroxidation in rat liver and kidney: effect of picroliv and silymarin. Phytotherapy Res. 15:307- 310.
Rastogi, R., A.K. Srivastava and A.K. Rastogi. 2001b. Biochemical changes induced in liver and serum of aflatoxin B1-treated male wistar rats: preventive effect of picroliv. Pharm. and Toxic. 88:53-58.
Ribeiro, S.M.R., G.M. Chagas, A.P. Campello and M.L.W. Kluppel. 1997. Mechanism of citrinininduced dysfunction of mitochondria .5. Effect on the homeostasis of the reactive oxygen species. Cell Bio. and Function 15:203-209.
Rizzo, A.F., F. Atroshi, M. Ahotupa, S. Sankari and E. Elovaara. 1994. Protective effect of antioxidants against free radical-mediated lipid peroxidation induced by DON or T-2 toxin. J. of Vet. Med. A. 41:81-90.
Sahu, S.C., R.M. Eppley, S.W. Page, G.C. Gray, C.N. Barton and M.W. O’Donnell. 1998. Peroxidation of membrane lipids and oxidative DNA damage by fumonisin B1 in isolated rat liver nuclei. Cancer Letters 125:117-121.
Schaeffer, J.L., J.K. Tyczkowski and P.B. Hamilton. 1987. Alterations in carotenoid metabolism during ochratoxicosis in young broiler chickens. Poultry Sci. 66:318-324.
Schaeffer, J.L., J.K. Tyczkowski and P.B. Hamilton. 1988. Depletion of oxycarotenoid pigments in chickens and the failure of aflatoxin to alter it. Poultry Sci. 67:1080-1088.
Schuster, A., G. Hunder, B. Fichtl and W. Forth. 1987. Role of lipid peroxidation in the toxicity of T-2 toxin. Toxicon 25:1321-1328.
Shen, H.M., C.Y. Shi, H.P. Lee and C.N. Ong. 1994. Aflatoxin B1-induced lipid peroxidation in rat liver. Toxic. and Appl. Pharm. 127:145-150.
Shen, H.M., C.N. Ong and C.Y. Shi. 1995. Involvement of reactive oxygen species in aflatoxin B1-induced cell injury in cultured rat hepatocytes. Toxicology 99:115-123.
Sies, H. 1999. Glutathione and its role in cellular functions. Free Radical Bio. and Med. 27:916- 921.
Souza, M.F., A.R. Tome and V.S. Rao. 1999. Inhibition by the bioflavonoid ternatin of aflatoxin B1-induced lipid peroxidation in rat liver. J. of Pharm. and Pharm. 51:125-129.
Suneja, S.K., D.S. Wagle. and G.C. Ram. 1989. Effect of oral administration of T-2 toxin on glutathione shuttle enzymes, microsomal reductases and lipid peroxidation in rat liver. Toxicol. 27:995-1001.
Surai, P.F. 2000. Effect of the selenium and vitamin E content of the maternal diet on the antioxidant system of the yolk and the developing chick. British Poultry Sci. 41:235-243.
Surai P.F. 2002. Natural antioxidants in avian nutrition and reproduction. Nottingham University Press.
Surai, P.F. and J.E. Dvorska. 2001. Is organic selenium better for animals than inorganic sources? Two different scenarios in stress conditions. Feed Mix 9:8-10.
Surai, P.F. and B.K. Speake. 1998. Distribution of carotenoids from the yolk to the tissues of the chick embryo. J. of Nutr. Bio. 9:645-651.
Surai, P.F., R.C. Noble and B.K. Speake. 1999. Relationship between vitamin E content and susceptibility to lipid peroxidation in tissues of the newly hatched chick. British Poultry Sci. 40:406- 410.
Surai, P.F., B.K. Speake and N.H.C. Sparks. 2001. Carotenoids in avian nutrition and embryonic development. 2. Antioxidant properties and discrimination in embryonic tissues. J. of Poultry Sci. 38:117-145.
Tobias, S., I. Rajic and A. Vanyi. 1992. Effect of T- 2 toxin on egg production and hatchability in laying hens. Acta Veterinaria Hungarica 40:47-54.
Tsuchida, M., T. Miura, T. Shimizu and K. Aibara. 1984. Elevation of thiobarbituric acid values in the rat liver intoxicated by T-2 toxin. Biochemical Medicine 31:147-166.
Tyczkowski, J.K. and P.B. Hamilton. 1987a. Altered metabolism of carotenoids during aflatoxicosis in young chickens. Poultry Sci. 66:1184-1188.
Tyczkowski, J.K. and P.B. Hamilton. 1987b. Metabolism of lutein diester during aflatoxicosis in young chickens. Poultry Sci. 66:2011-2016.
Wood, G.E. 1992. Mycotoxins in foods and feeds in the United States. J. of Anim. Sci. 70:3941-3949.
Yang, C.F., J. Liu, H.M. Shen and C.N. Ong. 2000. Protective effect of ebselen on aflatoxin B1- induced cytotoxicity in primary rat hepatocytes. Pharm. and Toxic. 86:156-161.
Yin, J.J., M.J. Smith, R.M. Eppley, S.W. Page and J.A. Sphon. 1998. Effects of fumonisin B1 on lipid peroxidation in membranes. Biochimica et Biophysica Acta. 1371:134-142.
1 Avian Science Research Centre, SAC, Auchincruive, Ayr, Scotland
2 Sumy State Agrarian University, Ukraine
3 Alltech North American Biosciences Center, Nicholasville, Kentucky, USA

An article which I was searching for a solution of effect on mycotoxin in causing SDS in broiler chicken. Very helpful and innovative.







