Oxidative stress in ruminants: enhancing productivity through antioxidant supplementation
Published: December 29, 2014
By: Pietro Celi 1,2, Chauhan SS2, Cottrell JJ2, F.R. Dunshea 2, Lean IJ 1,3, Leury BJ2, Liu F2 (1Faculty of Veterinary Science, The University of Sydney, MC Franklin Lab C04, Private Bag 4003, Narellan, NSW 2567, Australia. 2Faculty of Veterinary and Agricultural Sciences, The University of Melbourne, Parkville, VIC 3010, Australia, 3SBScibus, Camden, NSW 2570, Australia)
As the demand for animal protein continue to increase, global animal production faces several challenges in order to meet these demands because of environmental challenges (global warming and climate change). Furthermore, the intensification of animal production systems might compromise animal health and welfare and consequently increase the incidence of the metabolic diseases. Ruminant health and production is crucial for a sustainable animal production system, and this area of research is now attracting international interest, especially the mechanisms by which antioxidant supplementation may influence metabolism and health.
Oxidants play a central role in normal cellular function, providing an important feedback loop between metabolic activity and regulation of cellular functions. Oxidative stress (OS) arises due to an imbalance between pro-oxidants and antioxidants, which can occur under circumstances of increased antioxidant utilisation, impaired buffering capacity or immune function. In ruminant health and production the study of redox homeostasis is contributing to the understanding of important pathways involved in metabolic disorders. Indeed, OS seems to play a central role in the regulation of the metabolic activity of some organs and productivity in farm animals (Celi, 2011a). For example, during the last trimester of pregnancy and at the beginning of lactation the rapid foetal growth and the production of large amounts of colostrum and milk, an increase in both maternal and foetal metabolism result in increased reactive oxygen species production of during late gestation and early lactation and increased requirements for micronutrients, including antioxidants (Pedernera et al., 2010). Therefore manipulation of micronutrients and antioxidants has the potential to control the effects of oxidative damage, such as occurs during the transition period (Lean at al., 2014). This period is characterised by extremely high levels nutrient utilisation due to the growing foetus and then lactation diverts micronutrients and antioxidants from the cow to the mammary gland. The relationship between OS and disease and metabolic disorders is further demonstrated by lowered antioxidant status during mastitis, metritis, retained placenta, acidosis, ketosis and milk fever (Celi, 2011a).
It is important to consider that reducing equivalents, provided by NADPH, are consumed by both antioxidant defence and reactions catalysed by steroidogenic enzymes. Excessive utilisation of reducing equivalents during OS results in NADPH depletion and increase NADP concentrations despite elevated activity of the monophosphate shunt, which generates the reduced form (NADPH). The OS-induced depletion of reducing equivalents can reduce the supply of NADPH available for other important physiological processes and the consequent induction of the monophosphate shunt would divert glucose from other pathways such as milk production, resulting in decreased productivity (Figure 1).
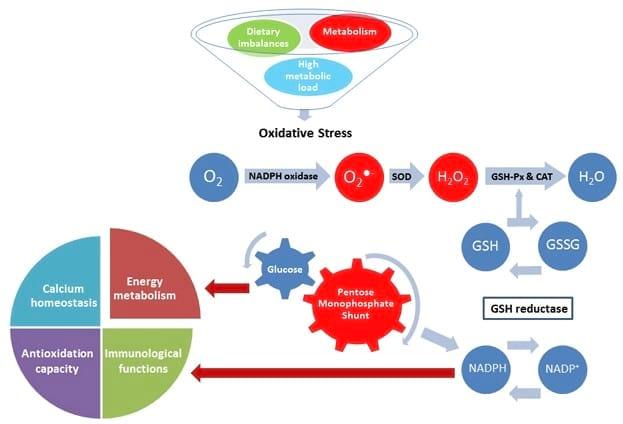
Oxidative stress
Figure 1.Superoxide (O2?-) is generated during normal metabolism by the activity of the NADPH oxidase. Dietary imbalances and high metabolic load also stimulate the activity of the NADPH oxidase leading to oxidative stress if not removed by the antioxidant system. In normal conditions, O2?- is converted in hydrogen peroxide (H2O2) by superoxide dismutase (SOD); H2O2 is then converted in water (H2O) by the activity of glutathione peroxidase (GSH-Px) and catalase (CAT). Reduction of peroxides is accompanied by oxidation of reduced glutathione (GSH) which can be regenerated from glutathione disulfide (GSSG) by reducing equivalents from NADPH, which is generated by the pentose monophosphate shunt. The resulting destruction of GSH increases consumption of reducing equivalents, diverting glucose from important physiological pathways and competing with NADPH dependent metabolic pathways such as energy metabolism, immunological functions, antioxidation capacity and calcium homeostasis.
An impairment of the antioxidant systems can often be the consequence of damage from ROS through deficiencies in the antioxidant vitamins and minerals and therefore a decrease in antioxidant status can be the direct consequence of an increase in ROS. Indeed, the strong link between the incidence of disease and OS has generated a strong research interest on antioxidants supplementation to improve animal health. Therefore, mineral and antioxidants dietary supplementation may provide a solution to boost ruminants’ health and productive performance (Leanet al., 2013).
While it is well recognised that adequate antioxidant concentrations including selenium and vitamin E are important for the maintenance of health and performance of ruminants (Chauhan et al., 2014a), responses to antioxidant supplementation have resulted in conflicting reports. It seems that productive and reproductive outcomes measured after the administration of these two antioxidants depend on the doses and on the different forms administered. Moreover, these apparent contradictory observations could be ascribed to the extensive interactions between antioxidants and the pro-oxidative stressors of the metabolism of ruminants per se and exerted through the environment. Indeed, there are significant interactions between antioxidants other dietary substrates (energy, protein) and environmental conditions such as heat stress.
Heat stress impairs animal performance and it has been implicated in promoting OS either through excessive ROS production or decreased antioxidant defences (Chauhan et al., 2014a). Therefore, a robust antioxidant network capable of preventing oxidative damage of biological molecules holds promise for improving the health and performance of animals during heat stress. Supplementation of sheep diets with supranutritional (greater than normally recommended) levels of vitamin E and Se ameliorates the negative effects of heat stress on feed intake, respiratory physiology, rectal temperature, acid base balance and OS (Chauhan et al., 2014a and 2014b)suggesting that dietary vitamin E and Se not only act as antioxidants, but also influence body homeostasis (namely thermoregulation) when administered at supranutritional levels.
Considering the complexity of numerous interactions between antioxidants and body systems (genome, proteome, and metabolome), it is conceivable that a thorough analysis of antioxidants-animal interactions is necessary to achieve a deeper understanding of the effects of antioxidant supplementation in ruminant diets. Future research should not only focus on the establishment of a reference panel of biomarker of OS, but should also address the issue of standardization of techniques and methodologies to study oxidative stress (Celi, 2011b). Clarity of understanding of the pathophysiology of OS in ruminants will allow designing specific antioxidant interventions. Selenium and Vitamin E represent ideal antioxidant supplements to improve ruminant health, production and reproduction, however, while the practical value of the use antioxidants in animal nutrition is obvious and clear, a thorough scientific approach is needed to explain their success and effectiveness. Finally, as OS has detrimental impacts on immune function and associated health disorders, future research need to focus on the maintenance of redox homeostasis especially during the transition period. More work is necessary to characterize the interactions between antioxidants for metabolic use during transition period or heat stress, and to investigate the homeorhetic mechanism which controls the fluxes of dietary antioxidants during metabolic and environmental challenges. Furthermore, the use of plants and plant parts such as Yerba Mate and moringa leaves that are rich in antioxidants is a new goal in livestock nutrition (Celi, 2013; Makkar et al. 2007; Rochfort et al. 2008). Exploitation of these novel resources in animal diets needs to be explored.
References
Celi, P (2011a) Oxidative Stress in Ruminants. In 'Studies on Veterinary Medicine.' (Eds L Mandelker, P Vajdovich.) Vol. 5 pp. 191-231. (Humana Press: New York). doi: 10.1007/978-1-61779-071-3_13
Celi, P (2011a) Oxidative Stress in Ruminants. In 'Studies on Veterinary Medicine.' (Eds L Mandelker, P Vajdovich.) Vol. 5 pp. 191-231. (Humana Press: New York). doi: 10.1007/978-1-61779-071-3_13
Celi, P (2011b) Biomarkers of oxidative stress in ruminants. Immunopharmacology & Immunotoxicology, 33(2): 233-240.
Celi, P. (2013). Yerba Mate (Ilex paraguariensis) as strategic supplement for dairy cows. In: Enhancing animal welfare and farmer income through strategic animal feeding - Some case studies. pp 11-18. Edited by Harinder P.S. Makkar. FAO Animal Production and Health Paper No. 175. Rome, Italy.
Chauhan, SS, Celi, P, Ponnampalam, EN, Leury, BJ, Liu, F, Dunshea, FR (2014a) Antioxidant dynamics in the live animal and implications for ruminant health and product (meat/milk) quality: role of vitamin E and Selenium. Animal Production Science, 54: 1525-1536.
Chauhan, SS, Celi, P, Leury, BJ, Clarke, IJ, Dunshea, FR (2014b) Dietary antioxidants at supranutritional doses improve oxidative status and reduce the negative effects of heat stress in sheep. Journal of Animal Science, 92: 3364-3374.
Lean, IJ, VanSaun, R, DeGaris, PJ (2013) Mineral and antioxidant management of transition dairy cows. Veterinary Clinics of North America: Food Animal Practice 29: 367-386.
Lean, IJ, DeGaris, PJ, Celi, P, McNeill, DM, Rodney, RM, Fraser, DR (2014) Influencing the future: interactions of skeleton, energy, protein and calcium during late gestation and early lactation. Animal Production Science, 54: 1177–1189.
Makkar, HPS, Francis, G, Becker, K (2007) Bioactivity of phytochemicals in some lesser-known plants and their effects and potential applications in livestock and aquaculture production systems. Animal 1: 1371-1391.
Pedernera, M, Celi, P, García, SC, Salvin, HE, Barchia, I, Fulkerson, WJ (2010) Effect of diet, energy balance and milk production on oxidative stress in early-lactating dairy cows grazing pasture. The Veterinary Journal 186: 352-357.
Rochfort, S, Parker, AJ, Dunshea, FR (2008) Plant bioactives for ruminant health and productivity. Phytochemistry 69: 299-322
This article was first published on Feedipedia - Animal Feed Resources Information System - INRA CIRAD AFZ and FAO. www.feedipedia.org
Related topics:
Authors:

The University of Sydney

University of Melbourne
Recommend
Comment
Share

Orffa Excentials
17 de abril de 2015
Good article. Congratulations and thank you for the figure, they are of great usefulness to fully understand physiological processes.
Regards,
Alfredo.
Recommend
Reply

5 de mayo de 2015
I found the article to be most informative and most definitely something our local nutritionists should be paying more attention to when formulating rations. Stress in any form is most debilitating and we do not pay enough attention to the underlying causes in our different animal production systems.
Thank you
Kind regards
Ralph
Recommend
Reply

Would you like to discuss another topic? Create a new post to engage with experts in the community.




You may be interested in
.jpg&w=3840&q=75)
Balchem Animal Nutrition