Dietary tributyrin, an HDAC inhibitor, promotes muscle growth through enhanced terminal differentiation of satellite cells
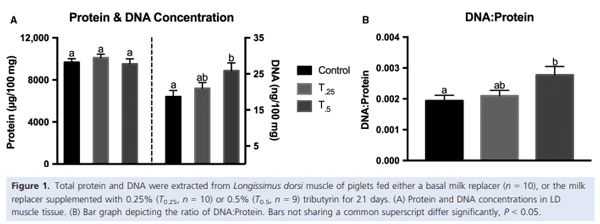
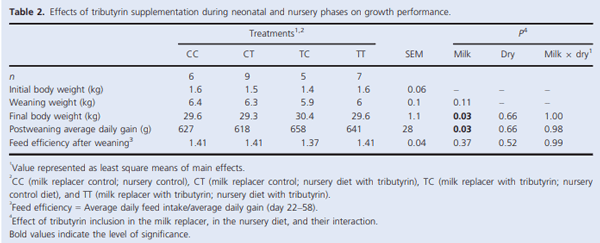
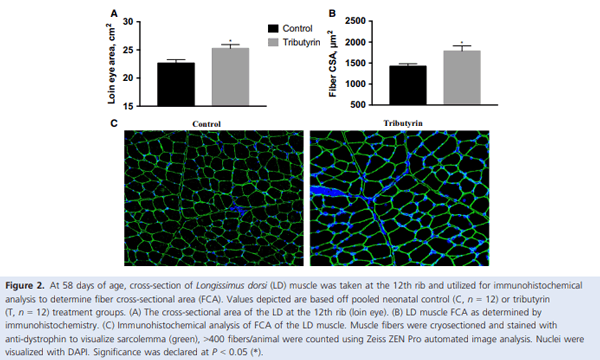
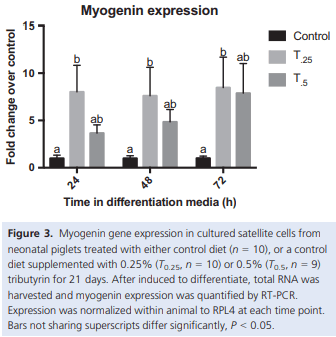
Muscle growth and repair rely on two main mechanisms – myonuclear accretion and subsequent protein accumulation. Altering the ability of muscle resident stem cells (satellite cells) to progress through their myogenic lineage can have a profound effect on lifetime muscle growth and repair. The use of the histone deacetylase (HDAC) inhibitor, butyrate, has had positive outcomes on the in vitro promotion of satellite cell myogenesis. In animal models, the use of butyrate has had promising results in treating myopathic conditions as well as improving growth efficiency, but the impact of dietary butyrate on satellite cells and muscle growth has not been elucidated. We investigated the impact of tributyrin, a butyrate prodrug, on satellite cell activity and muscle growth in a piglet model. Satellite cells from tributyrin-treated piglets had altered myogenic potential, and piglets receiving tributyrin had a ~40% increase in DNA:protein ratio after 21 days, indicating the potential for enhanced muscle growth. To assess muscle growth potential, piglets were supplemented tributyrin (0.5%) during either the neonatal phase (d1–d21) and/or the nursery phase (d21–d58) in a 2 9 2 factorial design. Piglets who received tributyrin during the neonatal phase had improved growth performance at the end of the study and had a ~10% larger loin eye area and muscle fiber cross-sectional area. Tributyrin treatment in the nursery phase alone did not have a significant effect on muscle growth or feed efficiency. These findings suggest that tributyrin is a potent promoter of muscle growth via altered satellite cell myogenesis.

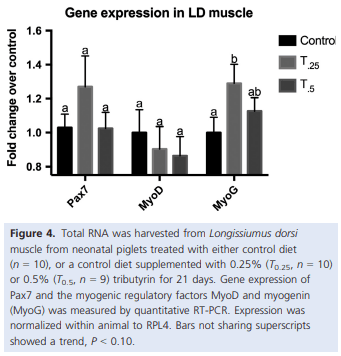
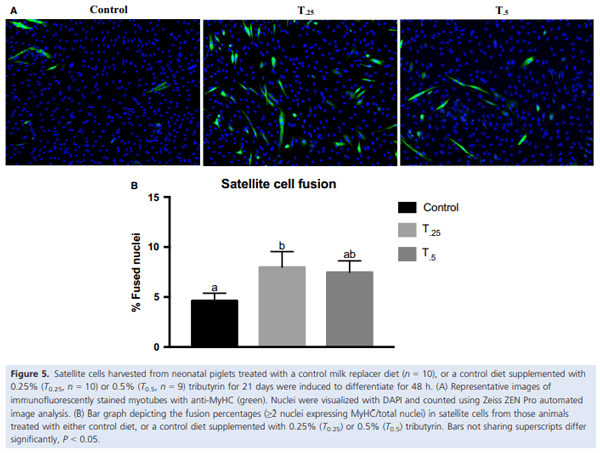
Alexander, L. S., B. S. Seabolt, R. P. Rhoads, and C. H. Stahl. 2012. Neonatal phosphate nutrition alters in vivo and in vitro satellite cell activity in pigs. Nutrients 4:436–448.
Allbrook, D. B., M. F. Han, and A. E. Hellmuth. 1971. Population of muscle satellite cells in relation to age and mitotic activity. Pathology 3:223–243.
Allen, R. E., C. J. Temm-Grove, S. M. Sheehan, and G. Rice. 1997. Skeletal muscle satellite cell cultures. Methods Cell Biol. 52:155–176.
Bedford, A., H. Yu, E. J. Squires, S. Leeson, and J. Gong. 2017. Effects of supplementation level and feeding schedule of butyrate glycerides on the growth performance and carcass composition of broiler chickens. Poult. Sci. 96:3221–3228.
Bentzinger, C. F., Y. X. Wang, and M. A. Rudnicki. 2012. Building muscle: molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 4:p. a008342.
Blais, A., M. Tsikitis, D. Acosta-Alvear, R. Sharan, Y. Kluger, and B. D. Dynlacht. 2005. An initial blueprint for myogenic differentiation. Genes Dev. 19:553–569.
Bodine, S. C., T. N. Stitt, M. Gonzalez, W. O. Kline, G. L. Stover, R. Bauerlein, et al. 2001. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 3:1014–1019.
Briggs, D., and J. E. Morgan. 2013. Recent progress in satellite cell/myoblast engraftment – relevance for therapy. FEBS J. 280:4281–4293.
Brown, S. C., and N. C. Stickland. 1994. Muscle at birth in mice selected for large and small body size. J. Anat. 184(Pt 2):371–380.
Campion, D. R., R. L. Richardson, J. O. Reagan, and R. R. Kraeling. 1981. Changes in the satellite cell population during postnatal growth of pig skeletal muscle. J. Anim. Sci. 52:1014–1018.
Candido, E. P., R. Reeves, and J. R. Davie. 1978. Sodium butyrate inhibits histone deacetylation in cultured cells. Cell 14:105–113.
Davie, J. R. 2003. Inhibition of histone deacetylase activity by butyrate. J. Nutr. 133:2485S–2493S.
Davis, T. A., and M. L. Fiorotto. 2009. Regulation of muscle growth in neonates. Curr. Opin. Clin. Nutr. Metab. Care 12:78–85.
Dong, L., X. Zhong, J. He, L. Zhang, K. Bai, W. Xu, et al. 2016. Supplementation of tributyrin improves the growth and intestinal digestive and barrier functions in intrauterine growth-restricted piglets. Clin. Nutr. 35:399–407.
Doumit, M. E., and R. A. Merkel. 1992. Conditions for isolation and culture of porcine myogenic satellite cells. Tissue Cell 24:253–262.
Dumont, N. A., C. F. Bentzinger, M. C. Sincennes, and M. A. Rudnicki. 2015. Satellite cells and skeletal muscle regeneration. Compr. Physiol. 5:1027–1059.
Duquet, A., A. Polesskaya, S. Cuvellier, S. Ait-Si-Ali, P. Hery, L. L. Pritchard, et al. 2006. Acetylation is important for MyoD function in adult mice. EMBO Rep. 7:1140–1146.
Edwards, J. D., and M. E. Butchbach. 2016. Effect of the butyrate prodrug pivaloyloxymethyl butyrate (AN9) on a mouse model for spinal muscular atrophy. J. Neuromuscul. Dis. 3:511–515.
Egorin, M. J., Z. M. Yuan, D. L. Sentz, K. Plaisance, and J. L. Eiseman. 1999. Plasma pharmacokinetics of butyrate after intravenous administration of sodium butyrate or oral administration of tributyrin or sodium butyrate to mice and rats. Cancer Chemother. Pharmacol. 43:445–453.
Fiszman, M. Y., D. Montarras, W. Wright, and F. Gros. 1980. Expression of myogenic differentiation and myotube formation by chick embryo myoblasts in the presence of sodium butyrate. Exp. Cell Res. 126:31–37.
Galfi, P., and J. Bokori. 1990. Feeding trial in pigs with a diet containing sodium n-butyrate. Acta Vet. Hung. 38:3–17.
Hasty, P., A. Bradley, J. H. Morris, D. G. Edmondson, J. M. Venuti, E. N. Olson, et al. 1993. Muscle deficiency and neonatal death in mice with a targeted mutation in the myogenin gene. Nature 364:501–506.
He, J., L. Dong, W. Xu, K. Bai, C. Lu, Y. Wu, et al. 2015. Dietary tributyrin supplementation attenuates insulin resistance and abnormal lipid metabolism in suckling piglets with intrauterine growth retardation. PLoS ONE 10: e0136848.
Hou, Y., L. Wang, D. Yi, B. Ding, X. Chen, Q. Wang, et al. 2014. Dietary supplementation with tributyrin alleviates intestinal injury in piglets challenged with intrarectal administration of acetic acid. Br. J. Nutr. 111:1748–1758.
Huang, C., P. Song, P. Fan, C. Hou, P. Thacker, and X. Ma. 2015. Dietary sodium butyrate decreases postweaning diarrhea by modulating intestinal permeability and changing the bacterial communities in weaned piglets. J. Nutr. 145:2774–2780.
Iezzi, S., G. Cossu, C. Nervi, V. Sartorelli, and P. L. Puri. 2002. Stage-specific modulation of skeletal myogenesis by inhibitors of nuclear deacetylases. Proc. Natl Acad. Sci. USA 99:7757–7762.
Iezzi, S., M. Di Padova, C. Serra, G. Caretti, C. Simone, E. Maklan, et al. 2004. Deacetylase inhibitors increase muscle cell size by promoting myoblast recruitment and fusion through induction of follistatin. Dev. Cell 6:673–684.
Johnston, L. A., S. J. Tapscott, and H. Eisen. 1992. Sodium butyrate inhibits myogenesis by interfering with the transcriptional activation function of MyoD and myogenin. Mol. Cell. Biol. 12:5123–5130.
Kotunia, A., J. Wolinski, D. Laubitz, M. Jurkowska, V. Rome, P. Guilloteau, et al. 2004. Effect of sodium butyrate on the small intestine development in neonatal piglets fed [correction of feed] by artificial sow. J. Physiol. Pharmacol. 55(Suppl 2):59–68.
Le Gall, M., M. Gallois, B. Seve, I. Louveau, J. J. Holst, I. P. Oswald, et al. 2009. Comparative effect of orally administered sodium butyrate before or after weaning on growth and several indices of gastrointestinal biology of piglets. Br. J. Nutr. 102:1285–1296.
Leeson, S., H. Namkung, M. Antongiovanni, and E. H. Lee. 2005. Effect of butyric acid on the performance and carcass yield of broiler chickens. Poult. Sci. 84:1418–1422.
Leibovitch, M. P., S. A. Leibovitch, M. Raymondjean, and J. Kruh. 1984. Effect of sodium butyrate on gene expression in a rat myogenic cell line. Biochem. Biophys. Res. Commun. 125:1129–1136.
Lu, H., S. Su, and K. M. Ajuwon. 2012. Butyrate supplementation to gestating sows and piglets induces muscle and adipose tissue oxidative genes and improves growth performance. J. Anim. Sci. 90(Suppl 4):430–432.
Ma, K., J. K. Chan, G. Zhu, and Z. Wu. 2005. Myocyte enhancer factor 2 acetylation by p300 enhances its DNA binding activity, transcriptional activity, and myogenic differentiation. Mol. Cell. Biol. 25:3575–3582.
Mal, A., M. Sturniolo, R. L. Schiltz, M. K. Ghosh, and M. L. Harter. 2001. A role for histone deacetylase HDAC1 in modulating the transcriptional activity of MyoD: inhibition of the myogenic program. EMBO J. 20:1739–1753.
Marks, P. A., V. M. Richon, and R. A. Rifkind. 2000. Histone deacetylase inhibitors: inducers of differentiation or apoptosis of transformed cells. J. Natl Cancer Inst. 92:1210– 1216.
McKinsey, T. A., C. L. Zhang, and E. N. Olson. 2001. Control of muscle development by dueling HATs and HDACs. Curr. Opin. Genet. Dev. 11:497–504.
Megeney, L. A., B. Kablar, K. Garrett, J. E. Anderson, and M. A. Rudnicki. 1996. MyoD is required for myogenic stem cell function in adult skeletal muscle. Genes Dev. 10:1173–1183.
Miller, A. A., E. Kurschel, R. Osieka, and C. G. Schmidt. 1987. Clinical pharmacology of sodium butyrate in patients with acute leukemia. Eur. J. Cancer Clin. Oncol. 23:1283–1287.
Minetti, G. C., C. Colussi, R. Adami, C. Serra, C. Mozzetta, V. Parente, et al. 2006. Functional and morphological recovery of dystrophic muscles in mice treated with deacetylase inhibitors. Nat. Med. 12:1147–1150.
Moresi, V., N. Marroncelli, and S. Adamo. 2015. New insights into the epigenetic control of satellite cells. World J. Stem Cells 7:945–955.
Olguin, H. C., and B. B. Olwin. 2004. Pax-7 up-regulation inhibits myogenesis and cell cycle progression in satellite cells: a potential mechanism for self-renewal. Dev. Biol. 275:375–388.
Olguin, H. C., Z. Yang, S. J. Tapscott, and B. B. Olwin. 2007. Reciprocal inhibition between Pax7 and muscle regulatory factors modulates myogenic cell fate determination. J. Cell Biol. 177:769–779.
Oustanina, S., G. Hause, and T. Braun. 2004. Pax7 directs postnatal renewal and propagation of myogenic satellite cells but not their specification. EMBO J. 23:3430–3439.
Piva, A., A. Prandini, L. Fiorentini, M. Morlacchini, F. Galvano, and J. B. Luchansky. 2002. Tributyrin and lactitol synergistically enhanced the trophic status of the intestinal mucosa and reduced histamine levels in the gut of nursery pigs. J. Anim. Sci. 80:670–680.
Piva, A., E. Grilli, L. Fabbri, V. Pizzamiglio, P. P. Gatta, F. Galvano, et al. 2008. Intestinal metabolism of weaned piglets fed a typical United States or European diet with or without supplementation of tributyrin and lactitol. J. Anim. Sci. 86:2952–2961.
Piva, A., M. Morlacchini, G. Casadei, P. P. Gatta, G. Biagi, and A. Prandini. 2016. Sodium butyrate improves growth performance of weaned piglets during the first period after weaning. Ital. J. Anim. Sci. 1:35–41.
Prasad, K. N. 1980. Butyric acid: a small fatty acid with diverse biological functions. Life Sci. 27:1351–1358.
Puchalska, P., and P. A. Crawford. 2017. Multi-dimensional roles of ketone bodies in fuel metabolism, signaling, and therapeutics. Cell Metab. 25:262–284.
Rehfeldt, C., I. Fiedler, G. Dietl, and K. Ender. 2000. Myogenesis and postnatal skeletal muscle cell growth as influenced by selection. Livest. Prod. Sci. 66:177–188.
Rudnicki, M. A., F. Le Grand, I. McKinnell, and S. Kuang. 2008. The molecular regulation of muscle stem cell function. Cold Spring Harb. Symp. Quant. Biol. 73:323–331.
Schultz, E. 1996. Satellite cell proliferative compartments in growing skeletal muscles. Dev. Biol. 175:84–94.
Seale, P., L. A. Sabourin, A. Girgis-Gabardo, A. Mansouri, P. Gruss, and M. A. Rudnicki. 2000. Pax7 is required for the specification of myogenic satellite cells. Cell 102:777–786.
Sincennes, M. C., C. E. Brun, and M. A. Rudnicki. 2016. Concise review: epigenetic regulation of myogenesis in health and disease. Stem Cells Transl. Med. 5:282–290.
Sun, Y., Y. Ge, J. Drnevich, Y. Zhao, M. Band, and J. Chen. 2010. Mammalian target of rapamycin regulates miRNA-1 and follistatin in skeletal myogenesis. J. Cell Biol. 189:1157– 1169.
Venuti, J. M., J. H. Morris, J. L. Vivian, E. N. Olson, and W. H. Klein. 1995. Myogenin is required for late but not early aspects of myogenesis during mouse development. J. Cell Biol. 128:563–576.
Walsh, M. E., A. Bhattacharya, Y. Liu, and H. Van Remmen. 2015a. Butyrate prevents muscle atrophy after sciatic nerve crush. Muscle Nerve 52:859–868.
Walsh, M. E., A. Bhattacharya, K. Sataranatarajan, R. Qaisar, L. Sloane, M. M. Rahman, et al. 2015b. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell 14:957– 970.
Wang, Y. X., and M. A. Rudnicki. 2012. Satellite cells, the engines of muscle repair. Nat. Rev. Mol. Cell Biol. 13:127– 133.
Yan, H., and K. M. Ajuwon. 2017. Butyrate modifies intestinal barrier function in IPEC-J2 cells through a selective upregulation of tight junction proteins and activation of the Akt signaling pathway. PLoS ONE 12:e0179586.
Yin, F., H. Yu, D. Lepp, X. Shi, X. Yang, J. Hu, et al. 2016. Transcriptome analysis reveals regulation of gene expression for lipid catabolism in young broilers by butyrate glycerides. PLoS ONE 11:e0160751.
Zammit, P. S., F. Relaix, Y. Nagata, A. P. Ruiz, C. A. Collins, T. A. Partridge, et al. 2006. Pax7 and myogenic progression in skeletal muscle satellite cells. J. Cell Sci. 119:1824–1832.


Dr. Chad, great article. With reference to their information, it can be deduced that the microbiota, due to the production of volatile fatty acids, including butyric, directly influences the health and development of pigs. Thus, it is evident that one of the desired characteristics in the strains of microorganisms that are used as probiotics is their ability to promote increased butyric acid production.






