Introduction
The conventional management in the swine industry is for weaning to start the sow’s next reproductive cycle. However, breeding sows during lactation has been proposed as an alternative that may increase sow productivity. If sows conceive while lactating, farrowing interval and nonproductive days may be decreased, increasing the litters per sow per year. Early attempts to breed sows during lactation yielded inconsistent results (Crighton, 19701 ; Stevenson and Davis, 19842 ; Newton et al., 1987a3 ), and the longer wean-to-estrus interval (WEI) in sow genotypes at the time may have contributed to the limited success. Nevertheless, these studies showed that a consistent feature of successful lactational ovulation is the reduction of the suckling stimulus to alleviate suckling-induced suppression of luteinizing hormone (LH). This allows follicular development similar to that normally occurring after the litter is weaned. Reduced suckling can be achieved by temporary separation from the litter (intermittent suckling, IS), or permanent removal of a portion of the litter (split-weaning, SW).
Recent attempts to induce lactational estrus have been more successful than those reported previously (Gerritsen et al., 20094 ; Terry et al., 20135 ; Terry et al., 20146 ). More than 90% of sows have been observed in estrus during lactation when suckling reduction is combined with daily boar exposure. Importantly, no decrease in sow fertility or litter growth parameters has been observed. An altered suckling method (ALT), combining elements of IS and SW, also yielded positive results in a recent study (Frobose et al., 20137 ), and may aid in reducing variation in pig BW. However, data directly comparing these suckling reduction methods are limited, and questions remain regarding the most practical method of application on farms.
Our objectives were to compare different suckling reduction strategies in boar-exposed lactating sows as a means to induce lactational estrus and to assess the impact on litter growth to market weight.
Procedures
Animals and Housing
The Kansas State University Institutional Animal Care and Use Committee approved this study. All experimental procedures were conducted at the Kansas State University Swine Teaching and Research Center in Manhattan, KS, from February through August 2013. The farrowing, gestation, and nursery barns used were totally enclosed, environmentally controlled, and mechanically ventilated. A total of 135 sows (PIC 1050; Hendersonville, TN) and their litters were used in 5 farrowing groups (35 d between groups). Parity ranged from 1 to 5 and averaged 2.6 ± 1.4. Each group included up to 29 sows that were moved into a single farrowing room on d 110 of gestation. Farrowing crates (7.0 × 2.0 ft for the sow and an additional 2.13 × 3.1 ft for the litter) were arranged in 2 parallel rows. Sows not farrowing by d 115 of gestation were injected IM with dinoprost tromethamine (Lutalyse®; 10 mg; Zoetis Animal Health, Florham Park, NJ) to induce parturition. Litter size at birth varied from 6 to 18 live pigs and was equalized within 2 d after farrowing by cross-fostering pigs, resulting in an average litter size of 11.5 ± 1.1 pigs. Pigs were individually weighed, ear-notched, and given 2 mL iron dextran and 1 mL of ceftiofur sodium (Naxcel®; Zoetis Animal Health, Florham Park, NJ) IM within 24 h post-farrowing. Male pigs were castrated approximately 7 d after birth. The day on which most of the litters were born was considered d 0 of lactation for the group, and all treatment procedures were performed on the same calendar day for all litters in the farrowing group. Litters were born between 4 d before to 2 d after d 0.
Feeding
Sows were fed a lactation diet (1,466 kcal/lb, 19.8% CP, and 1.11% total Lys) based on corn and soybean meal and fed in meal form. Lactation feed was provided ad libitum beginning the day after farrowing by individual Gestal Solo (JYGA Technologies, St-Nicolas, Quebec, Canada) electronic sow feeders. According to a predetermined feed curve, each sow was allowed to request up to 120% of the daily target before the next daily feeding period. Ad libitum water access was provided to sows and litters via cup waterers at floor level. Temperature in the farrowing house was maintained at a minimum of 68°F, and supplemental heat was provided to piglets with heat lamps. To standardize photoperiod, the artificial lights remained on for 24 h/d throughout lactation and post-weaning until ovulation was confirmed in all experimental animals.
From d 14 until weaning, a commercial Phase 1 nursery diet was offered in a rotary creep feeder (Rotechna Mini Hopper Pan, Rotechna SA, Agramount, Spain). The creep diet was fed in pellet form (2 mm pellets), and sufficient feed was maintained in the hopper to ensure that feed was always available. The creep feeder was available for all pigs including while temporarily separated from sows. Pigs also had access to a supplemental water source during litter separation.
All sows were tested during daily boar exposure by applying back pressure. At weaning, sows were moved into group pens (6 to 8) and daily estrus detection continued. Sows from 2 farrowing groups (n = 53; 9 to 12 sows per treatment) were examined daily by transrectal ultrasound for ovarian structures from d 17 until ovulation. After weaning, sows were temporarily moved into individual gestation stalls each day for ultrasound. Sows detected in estrus were moved to individual gestation stalls (7.0 × 2.0 ft) and fed 5 lb/d of a corn and soybean meal-based gestation diet (1,470 kcal/lb, 14.1% CP, and 0.56% Lys).
Treatments
On d 18 of lactation, sows were allotted to 1 of 5 treatments (n = 26 to 28) with parity, d 18 litter size (average 11.3 ± 1.2 pigs) and day of farrowing equalized as nearly as possible. Treatments were: 1) control; 2) altered suckling (ALT); 3) litter separation (SEP); 4) split-weaning (SW); and 5) 24 h litter separation (24HR). Control sows were managed according to standard farm practice, and their litters were continuously free to nurse until weaning on d 21. For sows in the 4 reduced suckling treatments, treatment commenced on d 18 and continued until weaning on d 25. The ALT sows were placed in adjacent pairs within the farrowing room such that 2 litters could be combined and rotated between sows by temporarily lifting the pen divider. On d 18, all but the 5 lightest-weight pigs from each ALT litter were SW and moved to the nursery. The remaining 10 lightweight pigs on paired ALT litters were combined to form a new litter of 10 pigs. These combined litters were rotationally-suckled (RS) between paired sows at 12 h intervals (0600 and 1800 h), and pigs had access to a sow 24 h/d, but each ALT sow was only nursed 12 h/d. Sows in the SEP treatment were also placed in adjacent pairs so that 2 complete litters could be combined during the daily 12-h (0600 to 1800 h) separation period in a common area (7.0 × 3.1 ft) created by removing the original crate divider and attaching new dividers to the sides of each sow’s individual crate. From 1800 to 0600 h, dividers were removed, and all pigs could move freely between and nurse either of the paired sows. For SW sows, all but the 5 lightest-weight pigs were weaned and moved to the nursery. The remaining pigs were allowed continuous nursing until weaning. Sows in the 24HR treatment were paired as in ALT and SEP treatments so pigs from the two adjacent 24HR litters could be combined in a common area between two crates as in the SEP treatment. On d 18, 24HR pigs were separated from the sow for a single 24 h period, after which 24HR pigs were returned to their original sow and allowed to continuously nurse until weaning.
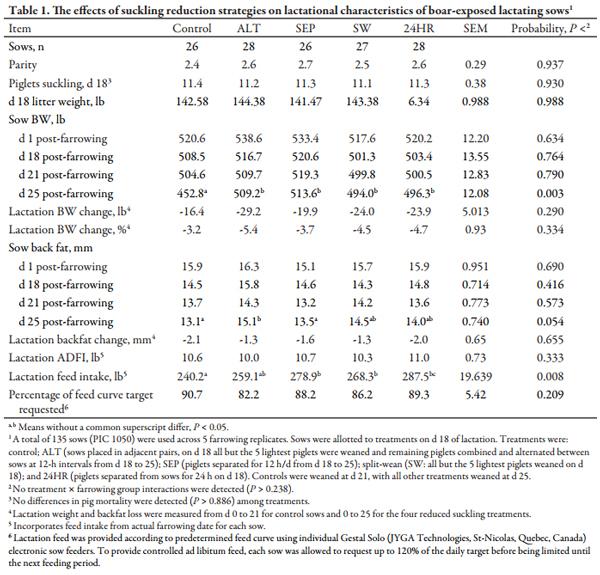
Beginning on d 18, all sows were provided daily boar exposure by moving a mature boar into the center aisle of the farrowing room between the 2 rows of center-facing farrowing crates. The boar was harnessed to a remote-controlled boar cart (BoarBot; Swine Robotics Inc., Leola, SD) and positioned between farrowing crates to deliver olfactory, audio, visual, and tactile (nose to nose) contact with sows. Each sow received approximately 5 min of contact. Two mature boars were used on alternating days. Boar exposure was provided from d 18 until ovulation or weaning. Sow BW and backfat thickness were measured at entry to the farrowing crate, post-farrowing, and on d 18, 21, and 25 post-farrowing. Daily lactation feed intake was also recorded.
Reproduction
Standing estrus was confirmed using a back-pressure test in the presence of a boar. Sows were artificially inseminated at first observed estrus and again 24 h later. Lactational and post-weaning inseminations were performed in the crate using post-cervical artificial insemination delivered during the refractory period immediately following standing estrus. Each insemination contained approximately 70 mL of extended semen (<5 d old) purchased from a commercial boar stud (Zoltenko Farms Inc., Courtland, KS). Pregnancy diagnosis was performed by transabdominal ultrasound (Hitachi-Aloka USA, Wallingford, CT) at 28 to 35 d after insemination. After pregnancy determination, sows were either culled or allocated to another experiment and, due to confounding treatment effects, subsequent reproductive performance was not collected.
Follicular Measurements
For all sows in 2 farrowing groups (replicates 2 and 4), transrectal ultrasound was performed once daily using an Aloka 500V ultrasound with a 5.0-MHz linear transducer (Hitachi-Aloka USA, Wallingford, CT) from d 17 until ovulation. Ovulation was considered to have occurred at 12 h prior to the ultrasound exam when less than four intact preovulatory follicles (usually 8 to 12 mm diameter) were found. At each scan, the number of follicles per ovary and the average diameter of the three largest follicles on each ovary were recorded. A sow was considered to have cystic follicles when multiple large structures (1 to 3 cm diameter) with anechoic interiors remained present for at least 5 d after estrus onset. Single large cystic follicles were detected occasionally, and these were noted and but not included in the follicle count. Single cysts are commonly observed in sows and apparently do not affect fertility. Therefore these latter sows were not considered cystic.
Hormone Analysis
Blood was collected from all sows on d 17, 21, and 25 and assayed for estradiol (E2). After estrus two additional samples were collected, one between 8 and 12 d and another between 18 and 21 d post-estrus to verify ovulation and confirm pregnancy recognition by extended elevated progesterone (P4), respectively. Jugular vein blood was collected using 38-mm × 20-gauge needles and 10 mL blood collection tubes without additive (Covidien Ltd., Mansfield, MA). After clotting for 6 h, the serum was separated by centrifugation (1,600 × g for 25 min at 39°F) and stored (-4°F) until analysis by RIA. Serum estradiol-17β (E2; MP Biomedical, Solon, OH) and P4 (Coat-A-Count, Siemens Medical, Los Angeles, CA) were analyzed in duplicate using commercial RIA kits. Assay sensitivity was 0.6 pg/mL for E2 and 0.06 ng/mL for P4. Intra-assay CV was 6.45 and 5.92%, for E2 and P4, respectively.
Piglet Measurements
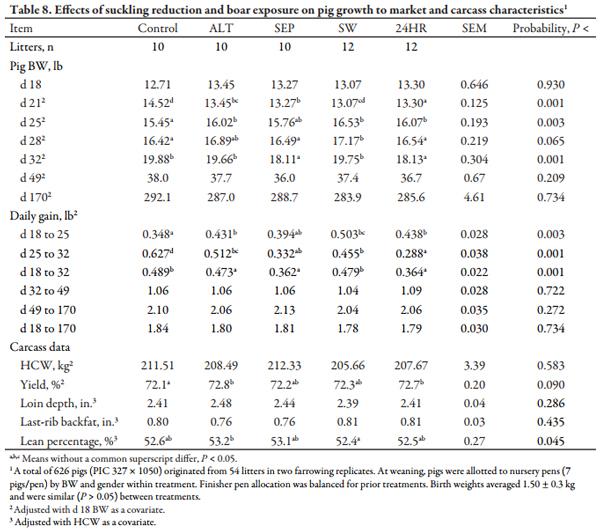
Pig growth to market was measured for all litters from 2 of the 5 farrowing groups (54 litters, 626 pigs). Weaned pigs were allotted to pens within treatment by BW and gender, with 7 pigs per pen. Nursery pens (4.0 × 5.0 ft) had woven wire flooring, a 3-hole, dry self-feeder, and a nipple waterer to allow ad libitum access to feed and water. Regardless of weaning age and sow treatment, pigs were fed according to the same feed budget consisting of 3 lb/pig of a commercial Phase 1 pelleted diet followed by 12 lb/ pig of Phase 2 diet and then Phase 3 until the end of the nursery phase (d 49). After exiting the nursery phase, pigs were moved to an on-site grower facility for 21 d prior to beginning the finishing phase. In both the grower and finisher facilities, pigs from each treatment were distributed as evenly as possible among pens. Pig BW was recorded at birth and at d 18, 21, 25, 28, 32, 49, and 170.
On d 170, pigs were weighed immediately prior to transport (approximately 122 mi) to a commercial abattoir (Triumph Foods Inc., St. Joseph, MO). Pigs were individually tattooed according to pen number to allow for data retrieval by pen and carcass data collection at the abattoir. Standard carcass criteria of percentage carcass yield, HCW, back fat depth, loin depth, and percentage lean were measured. Hot carcass weights were measured immediately after evisceration, and percentage yield was calculated by dividing HCW by live BW obtained at the farm prior to transport.
Data Analysis and Statistics
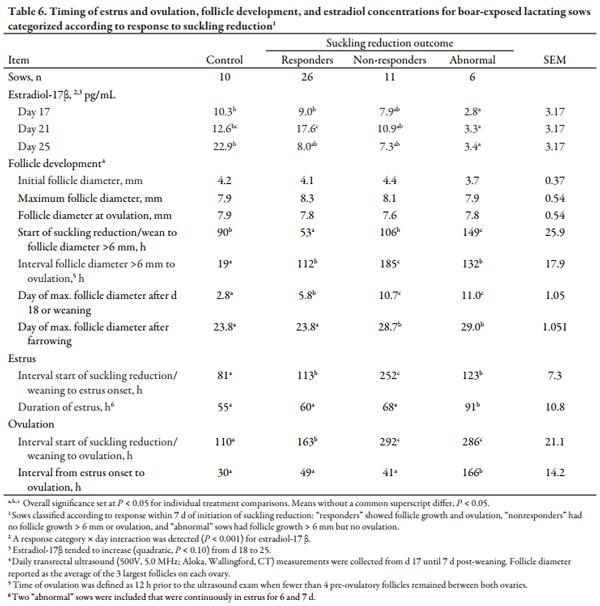
Progesterone concentrations were used to determine whether ovulation had occurred prior to d 18 post-farrowing and to confirm ovulation after visual estrus detection as well as the establishment of pregnancy. Ovulation was assumed to have taken place when P4 exceeded 4.0 ng/mL. Data in tables and figures are presented as least squares means ± maximal SEM. Normally distributed data were analyzed using the MIXED procedure of SAS, version 9.4 (SAS Institute, Inc., Cary, NC). Treatment means were compared using LSMEANS. For sow performance, analysis included the factors treatment and farrowing group and their interactions if significant (P < 0.05). Sow was the experimental unit and farrowing group was included in the model as a random effect. Conception rate and occurrence of lactational estrus were evaluated by χ2 analysis using the GLIMMIX procedure of SAS; however, controls were excluded from the lactational estrus analysis due to lack of variance. For serum E2 analysis, the statistical structure was the same except day of bleeding and treatment × d of bleeding served as fixed effects in addition to treatment. Day of bleeding also served as the repeated measure with sow as the subject. Post hoc testing was used to categorize the response of ultrasounded sows to suckling reduction treatments (n = 53). Sows were classified as: “responders” if follicle growth > 6 mm was observed with ovulation in < 7 d after initiating suckling reduction, “non-responders” if follicle growth > 6 mm did not occur within 7 d, and as “abnormal” if follicle growth > 6 mm progressed within 7 d, but the sow failed to ovulate.
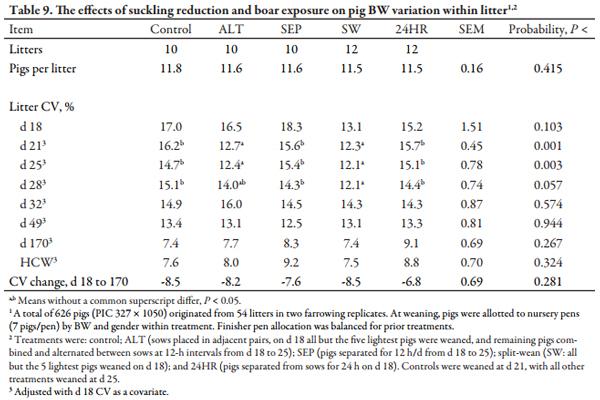
For pig growth and BW variation within litter (CV), d 18 pig BW was used as a covariate. For carcass performance, HCW was used as a covariate for fat depth, loin depth, and lean percentage. Although litter was considered to be the experimental unit, data were expressed on a per pig basis. Although control, SEP, and 24HR litters remained intact until weaning, to evaluate the effects of suckling reduction on heavy- and lightweight pigs, pig were retrospectively sorted into “heavy” and “light” categories using d 18 BW and the criteria applied to ALT and SW litters. For these comparisons, pig was the experimental unit, with sow, nursery pen, and farrowing group included as random effects. Least squares means were calculated for each independent variable, and treatment means were separated using PDIFFS in SAS. For all statistical tests, overall significance was set at P ≤ 0.10 and differences among individual treatments were considered significant at P ≤ 0.05 and trends if P ≤ 0.10.
Results and Discussion
The use of mature boars trained to work with the remote-controlled boar cart was an effective means to provide 5 min of boar contact daily. The remote control allowed one handler to maneuver the boar efficiently while simultaneously checking sows for estrus response. Although boar exposure for 3 d prior to weaning did not cause any control sows to show estrus during lactation, previous research has shown that boar exposure during late lactation can shorten the WEI and reduce the number of anestrus sows (Walton, 19868).
No treatment × farrowing group interactions were observed for sow BW, feed intake, or BF through d 25 post-farrowing (Table 1). Sows in the four reduced suckling treatments were heavier (P < 0.05) than controls at d 25 post-farrowing, and ALT sows had greater (P < 0.05) fat depth at d 25 versus control and SEP sows, with SW and 24HR sows intermediate. The SEP, SW, and 24HR sows had 4 d longer lactations and consumed more (P < 0.05) total lactation feed than controls. However, lactation ADFI was similar across treatments, and no differences were observed for backfat or BW changes during lactation. No differences in piglet mortality were observed during the 7 d of suckling reduction for any of the treatments.
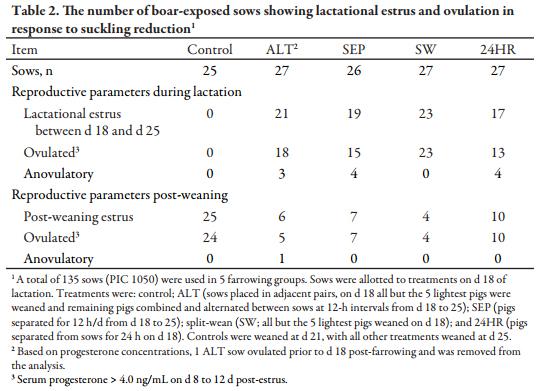
Despite receiving boar exposure from d 18 to 21, no control sows were in estrus or ovulated during lactation (Table 2). Based on P4 concentrations >4.0 ng/mL, 24 of 25 controls ovulated within 7 d post-weaning. According to P4 analysis, 1 ALT sow ovulated prior to d 18 post-farrowing, and she was removed from the dataset. A total of 21 of 27 ALT sows were in estrus during lactation, with 18 of those sows ovulating based on P4. Of the 3 ALT sows with lactational estrus but failing to ovulate, 2 continued to show estrus behavior for 6 and 7 d consecutively, and the third sow returned to estrus 4 d after weaning. Of 26 SEP sows, 19 exhibited estrus behavior during lactation and P4 concentrations indicate that 15 of those sows ovulated. Two of the 4 anovulatory SEP sows based on P4 were detected in estrus within 7 d after weaning. According to P4 analysis, all 23 of the SW sows exhibiting lactational estrus also ovulated. The 24HR treatment yielded 17 of 27 sows with lactational estrus, but 4 of those sows failed to ovulate during lactation and exhibited estrus behavior again 3 to 4 d after the initial estrus was observed. All 27 of the sows not responding with lactational estrus in the 4 reduced suckling treatments showed estrus within 7 d of weaning, and 26 of those sows ovulated based on serum P4 levels.
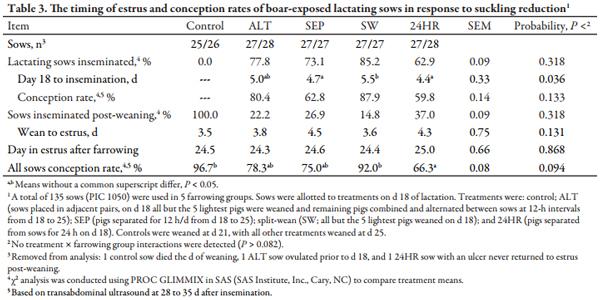
No treatment × farrowing group interactions were present regarding onset of estrus and conception rates (Table 3). Three sows were removed from the analysis due to death, ovulation prior to allotment, and an ulcer. For the remaining sows, the incidence of lactational estrus was not significantly different between reduced suckling treatments, ranging from 63 to 85%, with similar conception rates between those sows lactationally mated. Of sows in estrus during lactation, SEP and 24HR sows responded with lactational estrus more rapidly (P < 0.05) after d 18 compared to SW sows, with ALT sows intermediate. For sows exhibiting estrus post-weaning, the WEI was similar regardless of treatment. However, the overall conception rate for 24HR sows was lower (P < 0.05) than controls or SW sows, with ALT and SEP sows intermediate.
Sows within farrowing groups 2 and 4 (9 to 12 sows/trt) were examined by ultrasound daily from d 17 to ovulation (Table 4). Among those sows, all 10 control sows developed preovulatory-size follicles (> 6 mm) within 7 d of weaning, but collectively, 6 of the 41 sows in reduced suckling treatments did not respond to suckling reduction with follicles > 6 mm in the first 7 d. More than half of sows (26 of 41) in the four reduced suckling treatments responded to treatment initiation on d 18, with follicles in excess of 6 mm and ovulation within 7 d. While no control or SW sows showed “abnormal” follicle growth, 2 ALT sows, 5 SEP sows, and four 24HR sows developed preovulatory follicles within 7 d of treatment initiation but failed to ovulate. The 2 ALT sows remained in estrus for an extended period (6 to 7 d) and then ovulated. Three SEP and two 24HR sows fitting this categorization developed follicles > 6 mm, but these follicles regressed without ovulating. An additional 2 sows in both SEP and 24HR treatments appeared to ovulate based on ultrasound, however they then showed estrus within 7 d post-weaning and ovulated after the post-weaning estrus.
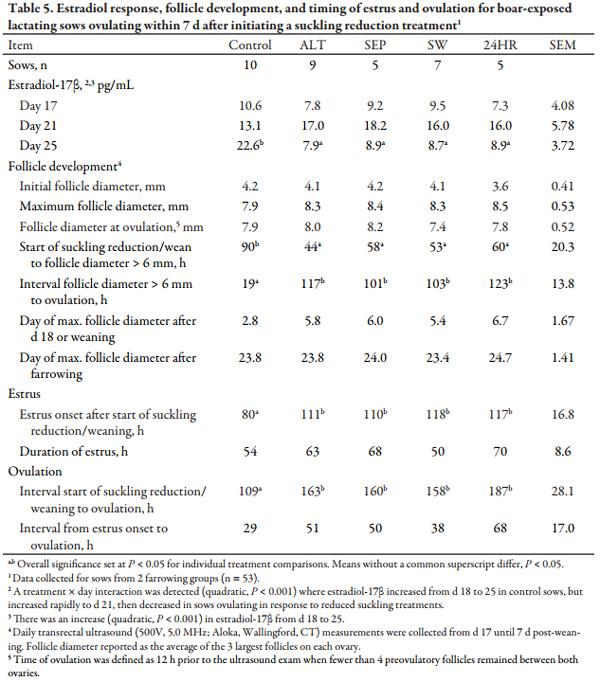
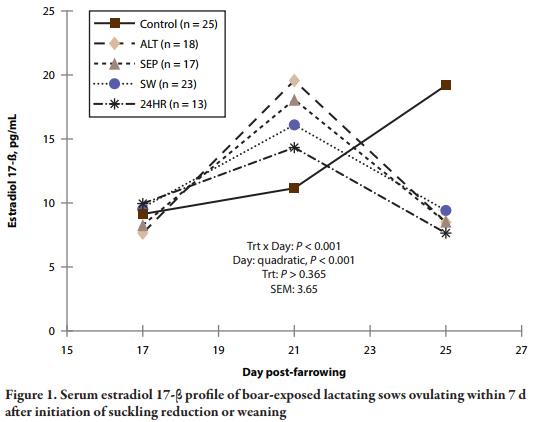
For sows ovulating within 7 d of suckling reduction or weaning (Table 5), a treatment × day interaction was detected in ultrasound (quadratic, P < 0.001) for E2 concentrations. For controls, E2 increased from d 18 to 25, but for reduced-suckling sows E2 increased to d 21 and then decreased (quadratic, P < 0.001) in sows ovulating in response to reduced suckling treatments. This coincides with the more rapid (P < 0.05) follicle growth to > 6 mm for sows in the four reduced suckling treatments. While other follicular characteristics were similar regardless of treatment, control sows ovulated more quickly (P < 0.05) after developing preovulatory-sized follicles than sows in reduced suckling treatments. Moreover, the interval from weaning to estrus in controls was shorter (P < 0.05) than the interval from initial suckling reduction to estrus in sows responding to suckling reduction.
The outcomes described in Table 4 as “responders,” “nonresponders,” and “abnormal” in response to suckling reduction treatment were compared in Table 6. A response category × day interaction was present (P < 0.001) for E2, as E2 increased from d 18 to 25 for controls, increased to d 21 then decreased in responders and in nonresponders to a lesser extent, while E2 was lowest (P < 0.05) in abnormal sows, regardless of the time point. Figure 1 presents the E2 responses for sows in estrus by 7 d after weaning or initial litter separation. The E2 profiles were generally similar. Control sows ovulated the fastest (P < 0.05) after follicles > 6 mm were present, and accordingly, reached maximum follicle diameter the fastest (P < 0.05) relative to weaning or treatment initiation; whereas nonresponders and abnormal sows reached maximum follicle diameter approximately 5 d later (P < 0.05) post-farrowing compared to controls and responders.
The nonresponders were commonly primiparous sows more likely to be in a negative energy balance due to the concurrent demands of growth and lactation (Hoving et al., 20119 ), and consequently less likely to exhibit lactational estrus (Stevenson and Davis, 1984a). Moreover, this experiment took place during the summer months, and due to limited body reserves at farrowing, primiparous sows are the most susceptible to the negative effects of high ambient temperature on lactation feed intake.
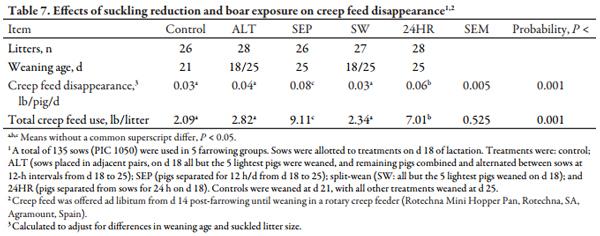
Table 7 shows that SEP litters had the greatest (P < 0.05) creep feed disappearance both on a litter basis and when adjusted and reported as lb/pig/d. While not to the magnitude of SEP litters, pigs in the 24HR treatment also had greater (P < 0.05) creep feed use than control, ALT, or SW litters. Day 21 pig weights depicted in Table 8 indicate increased (P < 0.05) pig BW for controls, reflecting the post-weaning growth check experienced by pigs weaned on d 18 in ALT and SW treatments, and the negative effect of decreased nursing time in SEP and 24HR pigs. Conversely, control pigs were lighter BW (P < 0.05) at d 25, the period after controls were weaned. Pigs from SW litters were heaviest (P < 0.01) on d 28 and, similar to control and ALT pigs, were heavier (P < 0.05) than SEP and 24HR pigs at d 32. Accordingly, d 18 to 32 ADG was lowest (P < 0.05) in SEP and 24HR pigs. Nonetheless, no differences in BW were found at the end of the nursery phase (d 49) or at marketing (d 170), congruent with the lack of a difference in ADG beyond d 32. Unexpectedly, carcass yield was decreased (P < 0.05) in control pigs compared to ALT and 24HR pigs, with SEP and SW pigs intermediary. Moreover, the greatest (P < 0.05) lean percentage was in ALT pigs and lowest (P < 0.05) in SW pigs, with other treatments similar.
Although the lactational estrus responses were poorer in SEP and 24HR treatments, they increased creep feed intake in the litters. They also decreased pig BW at weaning (d 25) due to limited nursing and potentially decreased milk yield, but SEP and 24HR pigs experienced a less marked post-weaning growth check. Other IS treatments have been reported to increase creep intake during late lactation and reduce the post-weaning growth check (Kuller et al., 200710; Berkeveld et al., 200711). They may help maintain gut health and function during the peri-weaning period. However, most reports agree that this benefit is temporary and not maintained to market weight (Matte et al., 199212). Based on these data, the additional creep intake and less severe post-weaning growth check induced by litter separation for 12 h/d or a single 24 h period is not sufficient to overcome the decreased pig weaning weight. However, one may consider short-term (1 to 3 d) applications of similar separation and commingling techniques as an alternative.
In a previous report (Frobose et al., 2013), the ALT treatment reduced litter BW variation compared to controls from d 18 to 32. In the present study, litters to which the ALT and SW treatments were applied had decreased (P < 0.01) BW variation on d 21 and d 25, and SW litters continued to have less (P < 0.05) within-litter BW variation versus controls, SEP, and 24HR litters until d 32 (Table 9). Nonetheless, BW variation within litter was similar at the end of the nursery and at marketing (d 170), with no differences between treatments for the change in CV from d 18 to d 170.
Taken together, the results of this study indicate that the suckling reduction strategies used vary in their ability to induce lactational estrus; with SW and ALT treatments responding similarly to sows conventionally mated post-weaning. Furthermore, 5 min of fence-line boar contact in front of the farrowing crate seems to be sufficient boar stimulus. Fertility of SW and ALT sows bred during lactation warrants additional investigation, as do the abnormal patterns of follicular development and estrus behavior observed in some sows. For pig performance, SEP and 24HR treatments stimulate creep feed intake, reduced the severity of the post-weaning growth check, and lightweight pigs in ALT and SW treatments benefited from the additional nursing access. However, effects on pig growth were transitory and were not maintained to market weight.
This article was originally published in Kansas Agricultural Experiment Station Research Reports: Vol. 1: Iss. 7. https://doi.org/10.4148/2378-5977.1146. This is an Open Access article licensed under a Creative Commons Attribution 4.0 License.