avian influenza (bird flu) in fowl
A fowl fear: is avian influenza (bird flu) on the leading edge of a global pandemic?
Avian influenza A (AI) H5N1, also known as Asian bird flu, is at the forefront of nearly every daily news report around the world. The reason for this intense interest in AI H5N1 is due to the growing fear that a global pandemic might occur if exchange of genetic material between H5N1 and a human influenza virus results in a virus that can be transmitted from person to person.
Avian influenza A viruses pose the threat of initiating new pandemics in humans because the human population is serologically naïve to most influenza hemagglutinin (HA) and neuraminidase (NA) subtypes that partially characterize each influenza virus. Until recently, it was thought that pigs were required as an intermediate host for transmission of AI viruses to humans (Gammelin et al., 1989; Yasuda et al., 1991), and it was believed that AI viruses, in general, did not replicate efficiently or cause disease in humans (Beare and Webster, 1991).
However, the AI H5N1 virus is the first avian influenza virus to have been documented as being capable of directly causing respiratory distress and death in humans (Yuen et al., 1998). To date 83 people have died after becoming infected by an AI H5N1 virus strain.
Most of those deaths have occurred in Asian countries. Ordinarily, the loss of 83 lives, spread sporadically among diverse populations and among different ages of people, would not generate such intense interest in the worldwide community, but the human mortality rate resulting from AI H5N1 infection is more than 50%, signaling to the world that AI H5N1 may be poised to cause the next human pandemic.
More than 75% of those human victims of AI H5N1 infection had recently been confirmed as being exposed to live or recently dead infected chickens or other birds. In October 2005, the appearance of the Asian bird flu virus in both wild birds and domestic fowl in Romania, Croatia, Turkey and Eastern European countries far from Southeast Asian countries known to have AI H5N1 virus drove global health officials to increase planning activities to prepare for the possibility that the H5N1 virus will infect humans worldwide.
There is ample evidence to show that periodically avian influenza A viruses do mutate to novel antigenic subtypes that cause disease and high rates of mortality in humans. It is believed that the 1918 influenza A (H1N1) pandemic, which killed 40 to 50 million people worldwide, was the result of such a mutation in an avian influenza A virus (Reid et al., 1999; Gamblin et al., 2004; Stevens et al., 2004). Similarly, emergence of human influenza A pandemics in 1957 (H2N2) and 1968 (H3N2) appeared to have had origins in avian/swine influenza A viruses (de Jong and Hien, 2006).
The 1918 Spanish flu virus (H1N1) is believed to have been due to the adaptation of a purely avian strain to infect humans directly, while in 1957 (H2N2) and 1968 (H3N2) the avian influenza A viruses adapted for human infection via genetic reassortment among avian, swine, and human virus strains (Webster et al., 1992; Murphy and Webster, 1996).
It has been a common belief among virologists that restriction of the avian influenza A viruses to birds has precluded its potential for efficient bird-tohuman transmission. The only other reports of natural infections of humans by highly pathogenic AI viruses were cases of conjunctivitis associated with avian influenza H7N7 viruses, transmitted either directly from birds to humans or by seals (Lang et al., 1981; Banks et al., 1998). Fouchier et al. (2004) and Koopmans et al. (2004) reported that even H7N7 was also the cause of one human death.
At present, AI H5N1 remains foremost an animal health problem, and human infection by AI H5N1 is not a common occurrence even though people have become infected, which oftentimes has been lethal. Fortunately, transmission of AI H5N1 from person to person has not been a problem; and in Vietnam, healthcare workers who had direct contact with AI H5N1-infected patients in 2003 and 2004 did not seroconvert to H5N1 (Liem and Lim, 2005).
In Table 1 are data reported to the World Health Organization (WHO), which show cumulative numbers of avian influenza A H5N1 cases in humans and mortality resulting from those infections. When compared with reported outbreaks in birds (Figure 1) there appears to be a parallel between outbreaks of AI H5N1 in poultry and in humans, but these reported outbreaks are inconsistent and probably reflect a far lower than actual incidence.
Table 1. Cumulative number of confirmed human cases of avian influenza H5N1 reported to the World Health Organization (January 16, 2006).
aIn 1997, Hong Kong had the first outbreak of avian influenza A H5N1 human infections with 18 reported cases and six known deaths. In early 2003, H5N1 caused an additional two infections with one death in a Hong Kong family with a recent travel history to southern mainland China.
Figure 1. Avian outbreaks of AI H5N1 through 6 January 2006 (WHO, 2006).
Avian influenza virus infection
The causative agent for Asian bird flu, AI H5N1, is classified as a member of the family Orthomyxoviridae genus Influenzavirus A (Swayne and Halvorson, 2003; Fouchier et al., 2005). Both the HA and the NA antigens on the virus contribute to its pathogenicity and are used in classifying it. There are 16 HA (H1-H16) subtypes and 9 NA (N1-N9) subtypes of surface glycoproteins that are recognized as being associated with AI viruses.
The H16 subtype was discovered only recently on influenza A viruses isolated from black-headed gulls from Sweden and Norway (Fouchier et al., 2005). HA is an antigenic glycoprotein found on the surface of all influenza viruses and is responsible for binding the virus to the cell that is being infected. The highly pathogenic H5N1 subtype appears to infect humans at a very low rate, but a single amino acid change in the H5N1 strain’s subtype H5 antigen has been found in human patients, and is believed to alter significantly the receptor specificity of avian H5N1 viruses, providing them with an ability to bind to receptors optimal for human influenza viruses (Gambaryan et al., 2006; Suzuki, 2005).
This finding seems to explain how an H5N1 virus that normally does not infect humans can adapt to efficiently infect human cells. Although H1, H2, and H3 subtypes have been associated with human influenza pandemics of the past, it is possible that any of the 16 different HA antigens can be involved in human influenza outbreaks, and therefore, H5 from H5N1 is indeed a potential candidate for involvement in a new human influenza outbreak.
The HA of the H5N1 virus has been associated with the high pathogenicity of this flu virus strain, apparently due to its ease of conversion to an active form by proteolysis (Senne et al., 1996; Hatta et al., 2001). HA has two primary functions: 1) the recognition of target vertebrate cells, accomplished through the binding of sialic acid-containing receptors (Weis et al., 1990), and 2) the fusion of host and viral endosomal membranes (White et al., 1997), accomplished through the recruitment of HA molecules to the fusion site where some undergo conformational alterations to destabilize the cell membrane lipid bilayer thereby forming a fusion intermediate which anneals the two bilayers between host and virus.
NA is an antigenic glycoprotein enzyme (EC 3.2.1.18) found on the surface of influenza viruses. Nine NA subtypes are known, and many occur only in various species of duck and chicken. Subtypes N1 and N2 have been positively linked to epidemics in man. Neuraminidase has functions that aid in the efficiency of virus release from cells.
Neuraminidase cleaves terminal sialic acid residues from carbohydrate moieties on the surfaces of infected cells. This promotes the release of progeny viruses from infected cells. Neuraminidase also cleaves sialic acid residues from viral proteins, preventing aggregation of viruses. Administration of chemical inhibitors of neuraminidase is a treatment that limits the severity and spread of viral infections. Neuraminidase inhibitor drugs currently in use for combating virus infections in humans are zanamivir and oseltamivir.
Prevalence of avian influenza (H5N1)
Countries with confirmed cases of H5N1 infections in poultry or wild birds are shown in Table 2 (CDC, 2006). The fact that AI H5N1 in domestic poultry and wild birds has been reported in Eastern Europe, throughout Asia, several unconfirmed cases in Western Europe, and in Africa suggests that there are large populations that have been exposed, but very few cases to date in humans.
Research in the United States has not revealed populations of wild birds or domestic fowl with AI H5N1 infections (Hanson et al., 2003; 2005), but H5, H7 and H9 avian influenza virus subtypes, which are associated with high pathogenicity in poultry, were detected in higher than expected levels (Hanson et al., 2003).
In an earlier review, Stallknecht and Shane (1988) demonstrated that AI viruses could be detected in 12 orders and 88 species of free living birds. Most of the isolates were from ducks, geese, swans and shore birds such as gulls and terns, which apparently serve as a reservoir for AI viruses similar to the findings of epidemiologists and virologists working on the current H5N1 problem in Asia (Murphy and Webster, 1996; Webster et al., 1992; 2006).
Table 2. Countries with confirmed avian influenza A (H5N1) infections in domestic poultry, wild birds, or mammals.
aPoultry
bWild birds
cMammals
Human infections with avian influenza H5N1 virus strains
Beigel et al. (2005) reported that most of the H5N1-infected individuals have been previously healthy children or older adults. The four who died in Turkey were also previously healthy children. In Asia, age of infected persons has varied from 1 to 60 years with median ages ranging from 9.5 in the 1997 Hong Kong outbreak to 22 in the 2005 Cambodian outbreak. There has been no indication that the gender of victims of the infection is a factor for concern. People who have developed the disease showed signs within two to eight days after exposure. Recently, incubation time has increased before showing signs of infection. Incubation times currently can range as long as eight to 17 days post exposure.
Among infected persons in the countries with an AI problem in humans, 70 to 100% had exposure to H5N1-infected live or recently dead chickens. While this observation seems compelling evidence of a direct movement of H5N1 to humans, there is the possibility of predisposing factors that might make the victims susceptible to H5N1 infections.
Most infected humans had a history of direct contact with poultry, although not those who were involved in mass culling. Plucking and preparing of diseased birds; handling fighting cocks; playing with poultry, particularly asymptomatic infected ducks; and consumption of raw duck blood or possibly undercooked poultry have all been implicated in the transmission of AI H5N1 to humans.
Transmission to felids has been observed by feeding raw infected chicken meat to tigers and leopards in zoos in Thailand (Keawcharoen et al., 2004; Thanawongnuwech et al., 2005) and to domestic cats under experimental conditions (Kuiken et al., 2004). Some infections may be initiated by pharyngeal or gastrointestinal inoculation of virus. In each possible case of avian to mammal transmission of the virus, body fluids bearing the virus had to have been involved.
Among these factors, there looms the possibility of genetic susceptibility of some victims (Perdue and Swayne, 2005) as illustrated by infection in some clusters in Turkey, Vietnam, Cambodia, and Thailand. An intercurrent disease that might have been immunosuppressive allowing the virus to be more infective, consumption of uncooked H5N1-infected poultry meat with direct delivery of the virus to receptive sites in the gastrointestinal tract (Mase et al., 2005a), or even nutritional deficiencies all might make the expression of the virus more pathogenically potent. It is important to note that family clusters of victims from H5N1 infections do not automatically indicate a mutation of the AI H5N1 virus making it more efficient in person-to-person transmission (Olsen et al., 2005).
However, conversion of the AI H5N1 virus to a virulent form or exposure to large numbers of infective organisms, i.e., dose response, that can infect humans seem to be the most probable causes of human outbreaks.
Regardless of the immense media interest and coverage of the AI H5N1 problem in Asia and now Eastern Europe, Perdue and Swayne (2005) rightly emphasize that the current Asian and Eastern European AI H5N1 outbreaks are primarily animal health problems. Though human infections are relatively few and sporadic this is not meant to diminish the potential human risk associated with exposure to the AI H5N1 virus. Perdue and Swayne (2005) draw attention to the fact that there is a major human risk, albeit a low risk, associated with the potential that a seasonally recurring influenza H1N1 or H3N2 strain will infect an H5N1-infected human and a reassortant virus containing the correct combination of genes for efficient human-to-human transmission will emerge.
Another issue is the unknown transmission efficiencies to other mammals and the likelihood of a co-infection of a mammal, such as a pig, with an avian and human influenza A virus strain. Currently circulating H1N2 and H3N2 swine influenza strains with genes from birds, human and swine influenza viruses can be isolated in both the United States and Asia (Olsen, 2002; Richt et al., 2003; Jung and Chae, 2004 ).
Evolution of influenza viruses
The AI H5N1 viruses involved in current outbreaks apparently are still evolving (Beigel et al., 2005). There have been changes in antigenicity and internal gene constellations (Horimoto et al., 2004; Li et al., 2005), expanded host range in avian species (Perkins and Swayne, 2002), ability to infect cats (Kuiken et al., 2004; Tiensin, 2005; Rimmelzwaan et al., 2006), enhanced pathogenicity in experimentally infected mice and ferrets (Zitzow et al., 2002), and increased environmental stability (Beigel et al., 2005). The fact that cats might be susceptible to AI H5N1 is a very important observation that provides evidence about the potential spread of the disease to and by a mammalian species within and between mammalian hosts.
Beare and Webster (1991) and, more recently, Gambaryan et al. (2004) state that avian influenza viruses generally do not replicate efficiently in human hosts. Gambaryan et al. (2004 and 2006) have reported that Asian AI H5N1 strains that infected humans in 2003 and 2004 had changed their receptor binding affinity. All but two of the isolates had preserved high affinity for Sia2-3Gal (avian-like) (3′SLN) receptors, but in 2003 those two exceptional isolates had decreased affinity to Sia2-3Gal and increased affinity to Sia2-6Gal (human-like) (6′SLN) receptors.
These receptors provide a mechanism for influenza virus to bind to host cells via their hemagglutinin (HA) surface proteins to sialylglycoconjugates exposed on the cell surface. Avian viruses preferentially bind to Sia2-3Gal-terminated sugar chains while mammalian viruses prefer to bind to Sia2- 6Gal-terminated structures (Conner et al., 1994; Gambaryan et al., 1997). Additionally these two exceptional AI H5N1 isolates had a unique Ser227-Asn change in the HA surface protein, which provided the AI H5N1 isolates the ability to bind to receptors that are optimal for human influenza viruses.
Gambaryan et al. (2006) showed that the 2003-2004 Asian H5N1 isolates from both chickens and humans had increased affinity for the sulfated trisaccharide Neu5Acα2-3Galß1-4(6-HSO3)GlcNAcß (Su-3′SLN) receptor differing from the 1997 Hong Kong isolates, which had increased affinity for the fucosylated Su-3′SLN receptor. American poultry H5 viruses also have increased affinity for the Su-3′SLN receptor. Gambaryan et al. (2006) concluded that genetic evolution of the avian influenza A H5N1 viruses is accompanied, during adaptation to poultry, by the evolution of their receptor specificity.
Virus evolution is driven by recombination of nucleic acids in the virus. Suarez et al. (2004) reported that a low pathogenic H7N3 AI virus had mutated in chickens to become a highly pathogenic H7N3 AI virus. That mutation was due to recombination between the hemagglutinin (HA) gene and the nucleoprotein genes. This recombination resulted in a 10 amino acid insert in the HA0 cleavage site, which made the H7N3 virus atypical among highly pathogenic viruses.
Among all of the virulent influenza A viruses, one virulence factor is correlated with the HA0 cleavage site. The surface HA glycoprotein is produced as a precursor HA0 and requires posttranslational cleavage by host proteases before it becomes functional, allowing the virus to become infectious (Rott, 1992).
All highly pathogenic AI viruses have multiple basic amino acids (arginine and lysine) at the HA0 cleavage site, and low pathogenic AI viruses have only two basic amino acids at the HA0 cleavage site (positions 1 and 4 for H5 viruses and positions 1 and 3 for H7 viruses) (Wood et al., 1993). The difference in HA0 cleavage sites between low pathogenic AI and highly pathogenic AI signals the difference in virulence.
The low pathogenic form is limited to cleavage by proteases at sites where trypsin-like enzymes are present such as in the respiratory and intestinal tracts while the highly pathogenic forms of AI with multiple cleavage sites can be cleaved by ubiquitous host proteases (Steineke et al., 1992) allowing them to replicate systemically, damaging vital organs and tissues leading to more severe disease and death. Uiprasertkul et al. (2005) reported from a single post-mortem finding that H5N1 virus could be found in the spleen of an AI H5N1-infected person, but did not find evidence of viral replication beyond the respiratory and intestinal tracts.
The highly pathogenic AI H5N1 virus that affected people in Hong Kong appears to have been the product of recombination between two other AI viruses (Chen et al., 1999; Chin et al., 2002; Claas et al., 1998; Peiris et al., 2004; Subbarao et al., 1998; Xu et al., 1999).
Claas reported that in May 1997, a 3-year-old boy in Hong Kong was admitted to the hospital and subsequently died from pneumonia, acute respiratory distress syndrome, Reye’s syndrome, multiorgan failure, and disseminated intravascular coagulation. An influenza A H5N1 virus was isolated from a tracheal aspirate. Preceding this incident, avian influenza outbreaks of high mortality were reported from three chicken farms in Hong Kong, and the virus involved was also found to be of the H5 subtype.
An antigenic and molecular comparison of the influenza A H5N1 virus isolated from the boy with one of the viruses isolated from outbreaks of avian influenza by hemagglutination-inhibition and neuraminidase-inhibition and nucleotide sequence analysis revealed differences in the antigenic reactivity of the viruses. However, nucleotide sequence analysis of all gene segments revealed that the human virus A/Hong Kong/ 156/97 was genetically closely related to the avian A/chicken/Hong Kong/258/97. Although direct contact between the sick child and affected chickens could not be established, they concluded that transmission of the virus from infected chickens to the child without another intermediate mammalian host acting as a ‘mixing vessel’ had occurred.
In 1996, a highly pathogenic AI H5N1 influenza virus, A/goose/Guangdong/1/96 (GSGD/96) was isolated from a sick goose in Southern China, but there was little interest in that virus at that time (Chen et al., 1999; Xu et al., 1999). In 1997 the highly pathogenic AI H5N1 virus, which apparently had derived its HA gene from a GSGD/96 like-virus and its other seven genes from the H6N1 subtype A/teal/Hong Kong/W312/ 97-like virus, was detected and caused disease outbreaks in chickens in Hong Kong live (wet) markets (Chin et al., 2002).
The virus was transmitted to humans causing fatal disease (Subbarao et al., 1998). Birds involved in this outbreak of H5N1 also died in large numbers, over a period of 48 hr. Even with only these two human cases resulting from this outbreak, it seemed clear that recombination among AI viruses can be a major cause of the evolution of potentially lethal viruses that can infect humans. This conclusion is supported by observations that most of the H5N1 viruses isolated before 2001 were unable to replicate in the mouse mammalian model, but viruses isolated in 2001 and 2002 replicated systemically and were highly lethal in the mouse model (Li et al., 2005).
Signs and pathology of avian influenza H5N1 infection
IN HUMANS
Signs of AI H5N1 infection are complex, and infected persons can be asymptomatic for some signs while showing many of the other signs (Beigel et al., 2005). Clinical signs tend to be somewhat variable among patients and include headache, myalgia, diarrhea, abdominal pain, vomiting, coughing, excessive sputum, sore throat, nasal drip, and shortness of breath (Beigel et al., 2005; de Jong and Hien, 2006). Some of the pathological developments associated with H5N1 infection include pulmonary infiltrates, lymphopenia, thrombocytopenia, elevated aminotransferase activity, cytokine and chemokine elevation, CD4+/CD8+ ratio inversion, hemophagocytosis, hepatic lobular necrosis, renal tubular necrosis, and lymphoid depletion (de Jong and Hien, 2006).
Many of the clinical signs can be related to immune-mediated pathology in the pathogenesis of the H5N1 infection (de Jong and Hien, 2006), which might lead to immunosuppression in patients. Indeed, it has been reported that humans with H5N1 infection have multiorgan failure despite the fact that viral replication is restricted to the lungs and gastrointestinal tract (Uiprasertkul et al., 2005). It was hypothesized that cytokine dysregulation might promote the organ failure in the pathogenesis of severe H5N1 disease (Chan et al., 2005; Cheung et al., 2002).
PATHOLOGY OF H5N1 INFECTION IN CHICKENS
Swayne and Halvorson (2003) have described the pathology associated with both low and highly pathogenic forms of avian influenza in poultry species. Lesions may be absent in cases of sudden death of chickens with highly pathogenic AI H5N1 infection because the virus acts so quickly (Nakatani et al., 2005).
In cases where mortality was slower to develop, observations at necropsy revealed congestion of the musculature, dehydration, subcutaneous edema of the head and neck areas, nasal and oral cavity discharge, severe congestion of conjunctiva sometimes with petechiae, excessive mucous exudate in the lumen of the trachea, severe hemorrhagic tracheitis, petechiae on the inside of the sternum, petechiae on the serosa and abdominal fat, petechiae on serosal surfaces and in the body cavity, severe renal congestion often with urate deposits in the tubules, hemorrhage and degeneration of the ovary, hemorrhages on the mucosal surface of the proventriculus especially at the junction with the ventriculus, hemorrhage and/or erosion of the ventricular lining and hemorrhagic foci on the lymphoid tissues in the intestinal mucosa.
Ducks shedding the highly pathogenic viruses may not show any of the clinical signs and lesions associated with highly pathogenic AI in chickens and turkeys.
All avian influenza virus strains are not highly pathogenic
AI H5N1 is a type of influenza virus that is hosted by birds but can infect several species of mammals. Wild waterfowl, primarily ducks, geese, swans and gulls are considered the natural reservoir of all influenza A viruses (Stallknecht and Shane, 1988; Webster et al., 1992; Murphy and Webster, 1996; Sturm-Ramirez et al., 2005). These wild birds have probably carried low pathogenicity forms of these influenza viruses, with no apparent harm, for long periods of time.
Virulent strains can emerge either by genetic mutation or by reassortment of less virulent strains. It is believed that AI H5N1 has been circulating in wild bird populations in China for some time before the discovery of the A/goose/ Guangdong/1/96 (GSGD/96) virus and its subsequent transfer to Hong Kong in sick chickens in 1997.
Domestic poultry are particularly susceptible to the more virulent forms (H5N1, H5N2, H7N2, H7N3, H7N7, and H9N2) (CDC, 2005; Mase et al., 2001; 2005b; Munster et al., 2005; Nakatani et al., 2005). They spread rapidly through poultry flocks, cause disease affecting multiple internal organs, and have a mortality that can approach 100%, often within 48 hr. Once avian flu is established in domestic poultry, it is a highly contagious disease and wild birds are no longer an essential ingredient for spread. Infected birds excrete virus in high concentration in feces and in nasal and ocular discharges.
The disease generally spreads rapidly in a flock by direct contact, but occasionally the spread is erratic. Pathogenic strains can emerge and cause disease in domestic poultry in any country at any time without warning.
In poultry flocks, AI exists in two main forms and is distinguished by low and high pathogenicity extremes of virulence (Swayne and Halvorson, 2003; Suarez et al., 2004; Perdue and Swayne, 2005). The highly pathogenic form causes systemic disease with rapid death in domestic poultry, but the low pathogenicity form, which causes localized infection, often causes little or no disease unless exacerbated by intercurrent disease or environmental stressors that may induce immunosuppression.
Low pathogenic AI viruses are found in wild birds around the world (Stallknecht and Shane, 1988; Murphy and Webster, 1996; Webster et al., 1992; Munster et al., 2005), and highly pathogenic H5 and H7 AI virus subtypes have been isolated from wild birds in Europe and Asia where the current outbreaks of H5N1 have occurred (Murphy and Webster, 1996; Webster et al., 1992; Munster et al., 2005). Highly pathogenic AI infections in poultry are rare and should not be confused with viruses of low pathogenicity, which can also be of H5 and H7 subtypes (Suarez et al., 2004; Perdue and Swayne, 2005).
Each virus has one HA and one NA protein which can be, potentially, in any combination. All of the HA and NA subtype combinations have been isolated from wild avian species (Swayne and Halvorson, 2003; SCWDS Fact Sheet, 2005). All highly pathogenic isolates have been influenza A viruses of subtypes H5 and H7, but not all H5 and H7 virus subtypes cause highly pathogenic influenza (Swayne and Halvorson, 2003; Suarez et al., 2004).
It has been demonstrated that avirulent H7 viruses can pass through chickens and mutate via antigenic shift into the highly pathogenic form (Li et al., 1991; Ito et al., 2001; Suarez et al., 2004). This potential for conversion of avirulent to virulent status via passage through a common host is why it is so important to monitor for AI and to depopulate flocks if AI is detected, even if few or no signs of diseases are present, in order to prevent highly pathogenic viruses from developing.
Antigenic shift vs. antigenic drift as a means for virus evolution
Influenza strains are named after their types of HA and NA surface proteins for example, H3N2 for type-3 HA and type-2 NA. If two different strains of influenza infect the same cell simultaneously, their protein capsids and lipid envelopes are removed, exposing their RNA, which is then transcribed to mRNA. The host cell then forms new viruses that combine antigens, for example, H3N2 and H5N1 can form H5N2 or H3N1 this way. Because the human immune system has difficulty recognizing the new influenza strain, it might become a highly infectious lethal virus responsible for global pandemics.
One increasingly worrying situation is the possible antigenic shift between avian influenza and human influenza. This antigenic shift could cause the formation of a highly virulent virus as is believed to have happened with the highly pathogenic AI H5N1 virus (Perdue and Swayne, 2005).
Recombination among low pathogenic AI viruses to form highly pathogenic AI viruses appears to be the manner in which the lethal forms evolve (Chen et al., 1999; Chin et al., 2002; Claas et al., 1998; Li et al., 2005; Peiris et al., 2004; Subbarao et al., 1998; Xu et al., 1999).
Therefore, it is important to understand how these transformations can take place. Antigenic shift is the process by which two different strains of influenza viruses combine and exchange genetic information to form a new subtype having a mixture of the surface antigens (HA and NA) of the two original strains. The term ‘antigenic shift’ is specific to the influenza literature (Figure 2) and in other viral systems, the same process is called reassortment or viral shift.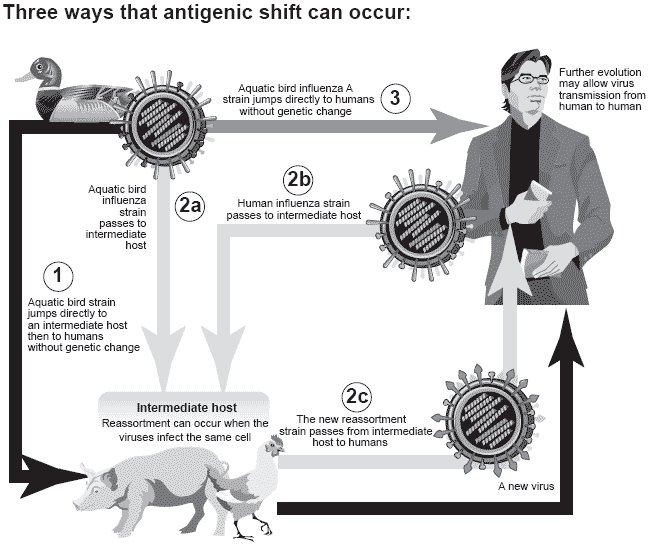
To enlarge the image click here
Figure 2. Illustration of the concept of antigenic shift in influenza viruses (NIH- National Institute of Allergy and Infectious Diseases, 2005).
Antigenic shift can happen in three ways:
Antigenic Shift 1
• Without undergoing genetic change, a bird strain of influenza A can jump directly from a duck or other aquatic bird to an intermediate animal host and then to humans.
• The new strain may further evolve to spread from person to person. If so, a flu pandemic could arise.
Antigenic shift 2
• A duck or other aquatic bird passes a bird strain of influenza A to an intermediate host such as a chicken or pig.
• A person passes a human strain of influenza A to the same chicken or pig.
• When the viruses infect the same cell, the genes from the bird strain mix with genes from the human strain to yield a new strain.
• The new strain can spread from the intermediate host to humans. Antigenic Shift 3
• Without undergoing genetic change, a bird strain of influenza A can jump directly from a duck or other aquatic bird to humans.
Antigenic shift is contrasted with antigenic drift (Figure 3), which is the natural mutation over time of known strains of influenza viruses to evade the immune system. Antigenic drift occurs in all types of influenza including influenza A, B and C. Antigenic shift, however, occurs only in influenza A because it infects more than just humans. Affected species include other mammals and birds, giving influenza A the opportunity for a major reorganization of surface antigens. Influenza B and C only infect humans, drastically minimizing the chance to mutate.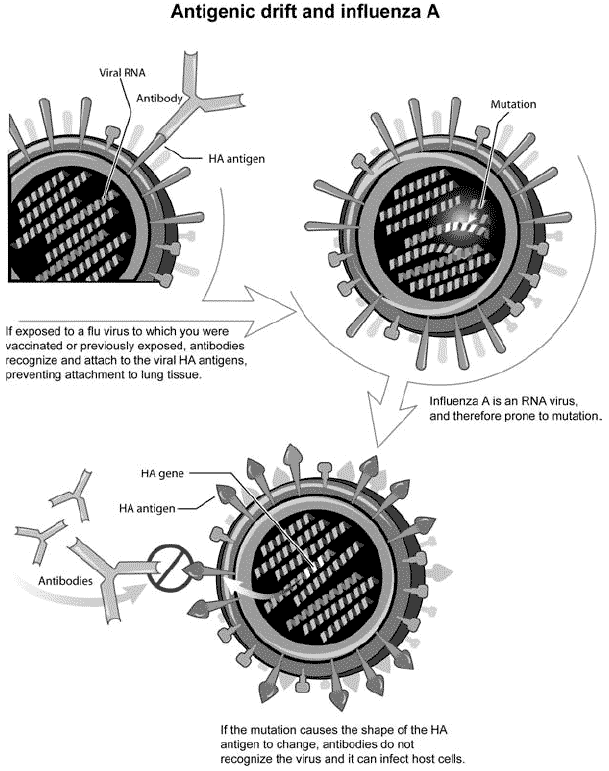
To enlarge the image click here
Figure 3. Illustration of the concept of antigenic drift in influenza viruses (NIH- National Institute of Allergy and Infectious Diseases, 2005).
Each year’s flu vaccine contains three flu strains - two A strains and one B strain, which change from year to year. After vaccination, the body produces infection-fighting antibodies against the three flu strains in the vaccine. If exposure occurs to any of the three flu strains during the flu season, the antibodies will bind to the virus’s HA antigens, preventing the flu virus from attaching to and infecting healthy cells.
Influenza virus genes, made of RNA, are more prone to mutations than genes made of DNA, and if there is a nucleotide alteration, the HA gene can change, causing the surface glycoprotein antigen to change conformation. If the HA antigen on the surface of the virus is altered, antibodies that normally would bind the viral antigen can no longer do so, allowing the newly mutated virus to infect the body’s cells. This type of genetic mutation is called ‘antigenic drift’.
Control and prevention of H5N1 infection in poultry and human populations
Influenza A viruses are found in many different animals, including ducks, chickens, pigs, whales, horses, and seals (Webster et al., 1992), but wild birds, which often are asymptomatic or show only mild infection, are the primary natural reservoir for all subtypes of influenza A viruses.
However, the range of symptoms in birds varies greatly, depending on the strain of virus. Infection with certain avian influenza A viruses (for example, some strains of H5 and H7 viruses) can cause widespread disease and death among some species of wild and especially domestic birds such as chickens and turkeys (Swayne and Halvorson, 2003). The potential for antigenic shift in influenza viruses with development of a new virus capable of person-to-person transmission is a constant problem that virologists have long feared.
Suarez et al. (2004) and Webster et al. (2006) pointed out that the avian influenza virus might undergo an antigenic shift with the human flu virus and cause a global influenza pandemic like the one in 1918. The human health implications have now gained importance, both for illness and fatalities that have occurred following natural infection with avian viruses, and for the potential of generating a reassortment virus that could give rise to a pandemic strain (Capua and Alexander, 2004). Thus, it is important that strategies for the control and prevention of highly pathogenic influenza A viruses be developed.
In the modern poultry industry, it is possible to contain H5N1 if coordinated programs involving depopulation, vaccination and strict biosecurity are practiced, but there is no known treatment. Recovered flocks still shed virus on an intermittent basis, requiring intense surveillance and subsequent removal. After an infected flock has been removed, it is possible to use commonly available detergents and disinfectants with heating and drying to inactivate AI H5N1 virus.
Nevertheless, any remaining organic material, such as feces, will continue to protect the virus, and active virus can be isolated from that material for at least 105 days. Thus, it is imperative that after an infected flock has been removed, all facilities and equipment must be cleaned and disinfected; and composting of litter is also recommended as a means to destroy residual virus (AVMA, 2005).
In Southeast Asia, more than 200 million chickens, ducks and other kinds of poultry have been culled in an effort to eliminate the virus. Swayne (2004) pointed out that all AI control programs should have three primary goals: 1) to prevent, 2) to manage, or 3) to eradicate AI.
To achieve these three goals, management procedures to prevent introduction or escape of the pathogen at the farm level (biosecurity enhancements) must be in place. Adequate diagnostic and surveillance detection of the pathogen or infections caused by it must be available. Infected animals must be destroyed and marketing of animals that have recovered or do not show signs of disease must be prevented. Host susceptibility must be decreased via vaccination programs or genetic selection pressure on animal breeds/strains that are resistant to the pathogen; and educational programs for people who produce poultry must be expanded to provide a better understanding of AI and to teach how to prevent or minimize transmission.
It has been reported that in Southeast Asia the spread of highly pathogenic AI H5N1 has occurred in small villages and hamlets, signifying that the spread is primarily among domestic fowl and ducks. These animals are raised in open pens/fields and can have contact with wild or migratory birds, which serve as a reservoir for H5N1.
Direct contact between humans and infected domestic fowl appears to be a mechanism for vectoring the virus to humans, and this has been demonstrated by the movement of the infected birds to wet markets in urban areas. The constant contact of the domestic fowl with infected wild birds/ducks will then provide the circumstance that allows recurring infections in the domestic fowl and further transfer to humans. This management practice for poultry production is problematic in the control of the spread of H5N1. Since these domestic fowl represent a significant portion of the livelihood of the rural people, the people will be unwilling to voluntarily destroy the birds without just compensation.
Therefore, it will be difficult to eliminate potential vectors of H5N1. Nevertheless, if just compensation were to be made for the eradication of the potentially infected poultry, control and management of its presence would be made easier. However, part of the control, management and eradication program is adequate diagnostic, surveillance, and detection of the pathogen or infections by the pathogen.
Even with the intense interest in H5N1, the lack of sufficiently trained healthcare workers in the rural and small urban areas of the affected countries makes it difficult to keep a potentially pandemic virus under control. Thus, developing countries, such as Vietnam, Cambodia, Laos, and Indonesia, where the H5N1 virus is endemic in birds, must become the battleground on which the virus must be contained to prevent transmission from birds to humans. Thailand has adopted a policy of eradication of infected flocks with compensation to their owners.
This approach will be much less costly to contain the virus before it becomes a pandemic causing virus.
Impact of avian influenza H5N1 on local poultry industries
It has been estimated by the World Bank that direct costs of the AI H5N1 outbreaks in the poultry industry in Vietnam can range from $30 million to more than $105 million. After the first outbreak in 2003-2004, there was an approximate 15% decline in poultry production that amounted to a revenue loss of $45 million. Nearly 50 million birds were slaughtered. Additionally, there was a decline in egg production of almost 913 million eggs representing an additional $46 million in lost revenue. Compensation amounted to about $2 million. Currently, there is an AI vaccination program planned for Vietnam costing an estimated $22 million. In Vietnam, after the 2003-2004 outbreaks, the swine industry raised its production by approximately 12% partially off-setting the economic impact of such large losses to poultry production.
In Turkey after the initial outbreak of H5N1 in October 2005, there was an almost 95% decline in poultry production and an almost total loss of egg sales. People were frightened to consume the poultry and eggs. Temporarily, nearly 100,000 people who worked in the poultry industry were directly affected by the H5N1 outbreaks.
In other countries where outbreaks of highly pathogenic H5N1 have occurred, nearly 200 million birds have been slaughtered in an attempt to contain the virus. The reported economic impact of H5N1 infection in the poultry industries in China, Turkey, Vietnam, Indonesia and Malaysia pales in comparison to the impact in Thailand. In Thailand and much of Southeast Asia, the poultry industry has been characterized as being divided into four separate but interactive sectors (FAO, 2005): commercial with high biosecurity (sector 1), commercial with moderate to high biosecurity (sector 2), local commercial with low biosecurity (sector 3), and local [backyard] with no biosecurity (sector 4).
Sector 1 is represented by an industrial integrated system, sector 2 is represented by a semivertical integrated system, sector 3 is represented by small, locally operated farms on which there may be caged layers and birds with access to the outdoors or even free roaming birds, and sector 4 is characterized by village or backyard flocks which include fighting cocks. Among these sectors, the vast majority of the AI H5N1 positive flocks have been associated with sector 4 birds (83%), and ducks were determined to be the silent carriers of the virus (Tiensin et al., 2005). To date Thailand has suffered more than $1.06 billion in lost revenue as the result of H5N1 infections in the poultry industry.
Most of the lost revenue was due to loss of export markets and there was a roughly 5% contraction in the poultry industry as a result of this loss. After the resurfacing of H5N1 in 2005, Thailand is now the leader in Southeast Asia in its attempts to eradicate the highly pathogenic virus and is aggressively combating the problem.
Thailand believes that one of the most critical problems associated with the spread of H5N1 in the country is movement of birds. As a means to control the spread of H5N1, movement of birds from sector 4 production is strictly regulated, more slaughter of infected birds, registration and regulation of fighting cocks is mandatory, mobile checkpoints were set up in the provinces most affected to enhance scrutiny of such movements, and surveillance teams have been deployed throughout the country.
At this time, Thailand is not vaccinating against avian influenza, but Thailand has promoted regional cooperation on containing the flu, proposing an Association of Southeast Asian Nations (ASEAN) animal hygienic fund and pledging substantial funds to start the project. The resulting center would enhance cross-border surveillance and control measures, as well as serve as an information distribution center for all ASEAN countries on the spread of H5N1. These measures are absolutely required, and any effort that is not coordinated among the Southeast Asian regional governments is doomed to failure.
While there is no known vaccine to protect humans against H5N1 at this time, massive vaccination programs have been designed to protect poultry from other forms of AI. In a report concerning the Hong Kong outbreaks, vaccination of chickens with a commercially available killed H5N2 vaccine was evaluated as an additional tool to enhance biosecurity measures and intensive surveillance for control of highly pathogenic AI H5N1 (Ellis et al., 2004).
In December 2002 to January 2003, there were outbreaks of H5N1 disease in waterfowl in two recreational parks, wild water birds, several poultry markets and five chicken farms.
In addition to quarantine, depopulation of the affected sheds and increased biosecurity, vaccination of the unaffected sheds and surrounding unvaccinated farms was undertaken on three farms. On at least two farms, infection spread to recently vaccinated sheds with low rates of H5N1 mortality when the chickens were between 9 and 18 days post-vaccination, but after 18 days post-vaccination no more deaths from H5N1 avian influenza occurred and intensive monitoring by virus culture on those farms showed no evidence of asymptomatic shedding of the virus. These observations suggested that an appropriate H5 vaccine might interrupt virus transmission in a field setting.
China now plans to vaccinate more than 5 billion chickens, there are over 150 million doses of AI vaccines scheduled for Vietnam, and other countries are considering the use of massive vaccination programs, as well.
Swayne and colleagues also have shown that it is possible to protect poultry species from highly pathogenic AI via vaccination similar to earlier reports (Taylor et al., 1988; Beard et al., 1991; Webster et al., 1991). Using a recombinant fowlpox-AI hemagglutinin subtype H5 gene insert vaccine, Swayne et al. (2000a) demonstrated that it was possible to protect broiler breeders and leghorn pullets from highly pathogenic H5 AI, but if the birds were given a secondary recombinant vaccination after a primary fowlpox vaccination, they found inconsistent protection.
Nevertheless, these observations supported the concept that highly pathogenic AI could be controlled via vaccination with an appropriate vaccine. Swayne and colleagues (2000b) further demonstrated that by using a recombinant fowlpox vaccine with the H5 gene insert, it was possible to protect poultry from several highly pathogenic H5 AI viruses. Even more important was the observation that the recombinant fowlpox vaccine with the H5 gene insert could protect against changes in field viruses if the H5 gene insert was similar to the challenge virus, and in this case natural antigenic drift did not interfere with general protection as has been reported for human influenza viruses.
The use of an AI vaccination program will control some of the pathogenic forms of AI, but not all forms (Capua and Marangon, 2004). It is known that vaccination for both low pathogenic and highly pathogenic forms of AI protects against clinical signs and mortality, reduces virus shedding, and increases resistance to infection (Capua et al., 2004). Capua and Marangon (2004) point out that the use of vaccination by itself is not going to eradicate AI. Much more is involved, because one must be able to differentiate vaccinated from infected animals.
In 2003 a method, Differentiating Infected from Vaccinated Animals (DIVA), was reported in Italy and was used in the regional effort to eradicate highly pathogenic AI (Capua et al., 2003). In Italy, where DIVA is widely credited for the eradication of AI, not only was vaccination used, but a territorial strategy was used in which biosecurity was upgraded, increased monitoring of vaccine efficacy was instituted, increased laboratory testing, more vigorous slaughter/controlled marketing of infected flocks, and financial compensation to farmers were instituted as well.
Capua and Marangon (2004) emphasize that in Asian countries where it is possible to implement an AI vaccination/DIVA plan such as that used in Italy, it might be possible to eradicate AI H5N1 before it becomes even more widespread. Some of the affected countries in Asia have not been able to develop such a plan and implement it, which suggests that the weakened plan would not be effective, leading to an endemic condition with highly pathogenic AI in poultry flocks in those countries. This then leaves upgrading biosecurity, use of an aggressive vaccination program coupled with depopulation of infected farms followed by disinfection and some down time.
The use of AI vaccination without DIVA is a problem because the vaccine virus in products might preclude their export to many other countries. Since it is possible to differentiate between vaccine virus and infective virus (Capua et al., 2003), the prospect of continued export trade becomes a distinct possibility, but all trading partners must reach a political/veterinary health agreement on the export/import of poultry that have been vaccinated. This lessens the prospect for lost revenue due to AI infections in poultry if trading partners have equivalent AI management and control programs.
A nutritional deficiency to exacerbate the avian influenza problem?
There are more than 13,000 scientific studies demonstrating increased susceptibility to infection in association with malnourishment in both lower vertebrate animals and humans, and a thorough review would be too extensive and complex for this paper. Most of those studies have focused on poor diets and how the host immune system had been negatively affected. In comparison, little is known about how the malnourished host may affect a virus and how that virus then interacts with the host. There is one ultramicro trace element, selenium, that seems to play a major role in the infections due to RNA viruses (Combs, 2001; Field et al., 2002; Lyons et al., 2003; Beck et al., 2004).
Research into the influence of malnutrition on host-viral interactions was initiated by Melinda A. Beck and colleagues at the University of North Carolina when she discovered the emergence of new viral variants in a Se-deficient model (Beck et al., 1994; 1995; 1998; 2003). Their observations revealed that selenium deficiency also affected the activity of infectious microorganisms invading the malnourished individuals.
In Beck’s research, the scientists found that mice deficient in selenium were more susceptible to coxsackievirus B3. Those mice infected with a normally harmless strain of coxsackievirus developed myocarditis because in the Se-deficient mice, the avirulent virus had mutated to a virulent form. The re-isolated virus was then shown to be virulent by inducing myocarditis in normal mice. GSH-Px knockout mice also developed myocarditis after being infected with a normally harmless viral strain, indicating that the antioxidant selenoenzyme GSH-Px-1 was critical in the normal, successful battle against viral infection.
The researchers sequenced viral samples isolated from Se-deficient mice and found mutations in the viral genome of the coxsackievirus that indicated increased virulence. The nutritional status of an individual is important not only for ensuring an optimal response of the immune system to an invading virus, but also in preventing viral mutations that can lead to increased pathogenicity of the virus itself.
Following Beck’s pioneering work, numerous other studies have revealed similar relationships involving host selenium deficiency and virulence of RNA viruses.
Selenium is very important in the maintenance of health status in HIV-infected patients (Foster, 2003). It has been reported that selenium inhibits HIV replication (Look et al., 1997) and reactivation by hydrogen peroxide (Sappey et al., 1994). Selenium also plays a protective role in patients with hepatitis B and C infections by preventing progression to cirrhosis and liver cancer (Yu et al., 1997; 1999). Measles virus-infected children given a 12 month nutritional supplement with selenium had significantly lower incidences of diarrhea, fever, and acute lower respiratory infection (Juyal et al., 2004).
Additionally, Broome et al. (2004) reported that patients given a live attenuated polio vaccine along with selenium supplementation had improved immune function and stopped shedding the polio vaccine virus more quickly, and higher rates of selenium supplementation (100 μg/day) was better than a lower rate (50 μg/day) of supplementation.
While the viruses mentioned above are important in the health status of people around the world, they do not necessarily represent risks for global pandemic infections in humans and animals alike. However, the work done with those viruses, which show the importance of selenium status in the host and how that status can affect virulence of the virus, is extremely important to the problem of influenza, especially today with AI H5N1 threatening to expand worldwide. Again, Beck and colleagues have demonstrated that even influenza virus is influenced by host selenium status (Beck et al., 2001; Nelson et al., 2001; Beck et al., 2004). Using the low pathogenic influenza A/Bangkok/1/79 (H3N2) virus strain that produces a mild pneumatitis in mice, Beck et al. (2001) found much more severe pathology in Se-deficient mice than in mice fed a Se-adequate diet.
Part of the increased level of pathology was due to increased proinflammatory cytokine and proinflammatory chemokine production in lungs of Se-deficient mice. Additional studies by Beck et al. (2001) revealed that influenza virus re-isolated from Se-deficient mice, when passed back to Se-adequate mice, caused those mice to develop much more severe pathology than in the Se-adequate mice challenged with virus re-isolated from selenium adequate mice.
The fact that Se-deficient mice developed more severe pathology than Se-adequate mice prompted investigation as to whether genomic change in the influenza virus in the Se-deficient mice. Beck and colleagues sequenced influenza A/Bangkok/1/79 RNA segments encoding for hemagglutinin (HA), neuraminidase (NA), and the matrix protein gene (M1), which are known virulence factors, from virus isolated from both Se-adequate and Se-deficient mice (Nelson et al., 2001). The increased virulence in the Se-deficient mice was due to mutations in the influenza virus genome, which resulted in a more virulent genotype. Most of the mutations occurred in the gene for the M1 protein.
A total of 29 different nucleotide changes were observed, and these changes were identical in three separate isolates taken from three separate Se-deficient mice. One to three mutations were seen in the genes encoding HA and NA in the Se-deficient mice. Once these mutations occur, selenium adequate hosts become susceptible to the new virulent variants of the influenza virus. Thus, poor selenium status might contribute to the emergence of new virus strains such as H5N1.
| Conclusion The veterinary and human medical workers involved in the worldwide efforts to prevent an AI H5N1 pandemic are doing a very difficult job with multitudinous obstacles before them. The media has portrayed AI H5N1 as a killer of immense proportions and has created fear that has had far-reaching consequences. Even though more than 50% of the people infected with highly pathogenic AI H5N1 have died, potential exposures to the virus have yielded less than 200 human cases. It is apparent that H5N1 must have certain requirements met before it is capable of infecting and causing illness in humans. Based on reports from around the world, it would appear that AI H5N1 is not the best candidate for creating the next global influenza pandemic. However, involvement of AI H5N1 in the generation of another virus, via antigenic shift, that can easily pass from birds to humans and then from human to human is highly probable. Therefore, it is imperative that control and management programs are put into place and that every flock known to be infected with highly pathogenic AI H5N1 be destroyed. Vaccination strategies against AI must be developed and development of DIVA programs for monitoring the progress of the vaccination/disease outbreaks must be made mandatory for all domestic poultry species in affected areas. Without these types of programs that ultimately lead to eradication of highly pathogenic AI virus in domestic poultry, the probability for a global influenza pandemic will continue to increase. Control and eradication of AI H5N1 and other highly pathogenic influenza viruses is not the problem of a single country, but must become the mandate of all countries, especially in Southeast Asia. Highly coordinated control and eradication measures and even vaccination programs for AI in poultry must be implemented. More importantly, control of movement of infected birds within and between countries that are affected by the AI H5N1 virus must be strictly regulated. |
References
American Veterinary Medical Association. 2005. Avian Influenza Backgrounder. Updated January 6, 2005.
Banks, J., E. Speidel and D.J. Alexander. 1998. Characterisation of an avian influenza A virus isolated from a human- is an intermediate host necessary for the emergence of pandemic influenza viruses? Arch. Virol. 143:781-787.
Beard, C.W., W.M. Schnitzlein and D.N. Tripathy. 1991. Protection of chickens against highly pathogenic avian influenza virus (H5N2) by recombinant fowlpox viruses. Avian Dis. 35:356-359.
Beare, A.S. and R.G. Webster. 1991. Replication of avian influenza viruses in humans. Arch. Virol. 119:37-42.
Beck, M.A., R.S. Ellsworthy, Y.S. Ho and F.F. Chu. 1998. Glutathione peroxidase protects mice from viral-induced myocarditis. FASEB 12:1143-1149.
Beck, M.A., J. Handy and O.A. Levander. 2004. Host nutritional status: The neglected virulence factor. TRENDS Microbiol. 12:417-423.
Beck, M.A., P.C. Kolbeck, L.H. Rohr, Q. Shi, V.C. Morris and O.A. Levander. 1994. Benign human enterovirus becomes virulent in selenium-deficient mice. J. Med. Virol. 43:166-170.
Beck, M.A., O.A. Levander and J. Handy. 2003. Selenium deficiency and viral infection. J. Nutri. 133:1463S-1467S.
Beck, M.A, H.K. Nelson, Q. Shi, P. Van Dael, E.J. Schiffrin, S. Blum, D. Barclay and O.A. Levander. 2001. Selenium deficiency increases the pathology of an influenza virus infection. FASEB 15:1481-1483.
Beck, M.A., Q. Shi, V.C. Morris and O.A. Levander. 1995. Rapid genomic evolution of a non-virulent Coxsackievirus B3 in selenium-deficient mice results in selection of identical virulent isolates. Nature Med. 1:433-436.
Beigel, J.H., J. Farrar, A.M. Han, F.G. Hayden, R. Hyer, M.D. de Jong, S. Lochindarat, N.T.K. Tien, N.T. Hien, T.T. Hien, A. Nicoll, S. Touch and N-Y. Yuen. 2005. Avian influenza A (H5N1) infection in humans. New England J. Med. 353:1374-1385.
Broome, C.S., F. McArdle, J.A. M. Kyle, F. Andrews, N.M. Lowe, C.A. Hart, J.A. Arthur and M.J. Jackson. 2004. An increase in selenium intake improves immune function and poliovirus handling in adults with marginal selenium status. Am. J. Clin. Nutri. 80:154-162.
Capua, I. and D. Alexander. 2004. Avian influenza: Recent developments. Avian Pathol. 33:393-404.
Capua, I. and S. Marangon. 2004. Vaccination for avian influenza in Asia. Vaccine 22:4137-4138.
Capua, I., C. Terregino, G. Cattoli and A. Toffan. 2004. Increased resistance of vaccinated turkeys to experimental infection with an H7N3 low pathogenicity avian influenza virus. Avian Pathol. 33:158-163.
Capua, I., C. Terregino, G. Cattoli, F. Mutinelli and J. F. Rodriguez. 2003. Development of a DIVA (Differentiating Infected from Vaccinated Animals) strategy using a vaccine containing a heterologous neuraminidase for the control of avian influenza. Avian Pathol. 32:47-55.
Centers for Disease Control and Prevention. 2005. Avian influenza infection in humans. http://www.cdc.gov/flu/avian/gen-info/avian-flu-humans.htm.
Centers for Disease Control and Prevention. 2006. Avian influenza (bird flu): recent avian influenza outbreaks in Asia and Europe. http://www.cdc.gov/flu/avian/outbreaks/ asia.htm.
Chan, M.C.W., C.Y. Cheung, W.H. Chui, S.W. Tsai, J.M. Nicholls, Y.O. Chan, R.W.Y. Chan, H.T. Long, L.L. M. Poon, Y. Guan and J.S.M. Peiris. 2005. Proinflammatory cytokine responses induced by influenza A (H5N1) viruses in primary human alveolar and bronchial epithelial cells. Resp. Res. 6:135-147.
Chen, H., K. Yu and Z. Bu. 1999. Molecular analysis of hemagglutinin gene of goose origin highly pathogenic avian influenza virus. Chin. Agric. Sci. 32:87-92.
Cheung, C.Y., L.L. Poon, A.S. Lau, W. Luk, Y.L. Lau, K.F. Shortridge, S. Gordon, Y. Guan and J.S. Peiris. 2002. Induction of proinflammatory cytokines in human macrophages by influenza A (H5N1) viruses: a mechanism for the unusual severity of human disease? Lancet 360:1831-1837.
Chin, P.S., E. Hoffmann, R. Webby, R.G. Webster, Y. Guan, M. Peiris and K.F. Shortridge. 2002. Molecular evolution of H6 influenza viruses from poultry in Southeastern China: prevalence of H6N1 influenza viruses possessing seven A/Hong Kong/156/97 (H5N1)- like genes in poultry. J. Virol. 76:507-516.
Claas, E.C., A.D.M.E. Osterhaus, R. vanBeek, J.C. de Jong, G.F. Rimmelzwaan, D.A. Senne, S. Krauss, K.F. Shortridge and R.G. Webster. 1998. Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet 351:472-477.
Combs, Jr., G.F. 2001. Selenium in global food systems. Brit. J. Nutri. 85:517-547.
Conner, R.J., Y. Kawaoka, R.G. Webster and J.C. Paulson. 1994. Receptor specificity in human, avian, and equine H2 and H3 influenza virus isolates. Virol. 205:17-23.
de Jong, M.D. and T.T. Hein. 2006. Avian influenza A (H5N1). J. Clin. Virol. 35:2-13.
Ellis, T., C.Y. Leung, M.K. Chow, L.A. Bissett, W. Wong, Y. Guan and J.S. Peiris. 2004. Vaccination of chickens against H5N1 avian influenza in the face of an outbreak interrupts virus transmission. Avian Pathol. 33:405-412.
Field, C.J., I.R. Johnson and P.D. Schley. 2002. Nutrients and their role in host resistance to infection. J. Leuk. Biol. 71:16-32.
Food and Agriculture Organization. 2005. FAO recommendations on the prevention, control and eradication of highly pathogenic avian influenza (HPAI) in Asia. http:// www.fao.org/ag/againfo/subjects/en/health/diseases-cards/27septrecomm.pdf.
Foster, H.D. 2003. Why HIV-1 has diffused so much more rapidly in Sub-Saharan Africa than in North America. Med. Hypoth. 60:611-614.
Fouchier, R.A.M., P.M. Schneeberger, F.W. Rozendaal, J.M. Broekman, S.A.G. Kemink, V. Munster, T. Kuiken, G.F. Rimmelzwaan, M. Schutten, G.J.J. van Doornum, G. Koch, A. Bosman, M. Koopmans and A.D.M.E. Osterhaus. 2004. Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proc. Natl. Acad. Sci. USA 101:1356-1361.
Fouchier, R.A.M., V. Munster, A. Wallensten, T. M. Bestebroer, S. Herfst, D. Smith, G.F. Rimmelzwaan, B. Olsen and A.D. Osterhaus. 2005. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J. Virol. 79:2814-2822.
Gambaryan, A.S., A.B. Tuzikov, V.E. Piskarev, S.S. Yamnikova, D.K. Lvov, J.S. Robertson, N.V. Bovin and M.N. Matrosovich. 1997. Specification of receptor binding phenotypes of influenza virus isolates from different hosts using synthetic sialylglycopolymers: non-egg-adapted human H1 and H3 influenza B viruses share a common binding affinity for 6’-sialyl(N-acetyllactosamine). Virol. 232:3455-350.
Gambaryan, A.S., A.B. Tuzikov, G.V. Pazyninia, R.G. Webster, M.N. Matrosovich, and N.V. Bovin. 2004. H5N1 chicken influenza viruses display a high binding affinity for Neu5Aca2-3Galb1-4(6-HSO3)GlcNAcb-containing receptors. Virol. 326:310-316.
Gambaryan, A., A. Tuzikov, G. Pazyninia, N. Bovin, A. Balish and A. Klimov. 2006. Evolution of the binding phenotype of influenza A (H5) viruses. Virol. 344:432-438.
Gamblin, S.J., L.F. Haire, R.J. Russell, D.J. Stevens, B. Xiao, Y. Ha, N. Vasisht, D.A. Steinhauer, R.S. Daniels, A. Elliot, D.C. Wiley and J.J. Skehel. 2004. The structure and receptor binding properties of the 1918 influenza hemagglutinin. Science 303:1838- 1842.
Gammelin, M., J. Mandler and C. Scholtissek. 1989. Two subtypes of nucleoproteins (NP) of influenza A viruses. Virol. 170:71–80.
Hanson, B.A., D.E. Stallknecht, D.E. Swayne, L.A. Lewis and D.A. Senne. 2003. Avian influenza viruses in Minnesota ducks during 1998-2000. Avian Dis. 47:867- 871.
Hanson, B.A., D.E. Swayne, D.A. Senne, D.S. Lobpries, J. Hurst and D.E. Stallknecht. 2005. Avian influenza viruses and paramyxoviruses in wintering and resident ducks in Texas. J. Wildlife Dis. 41:624-628.
Hatta, M., P. Gao, P. Halfmann and Y. Kawaoka. 2001. Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses. Science 293:1840-1842.
Horimoto, T., N. Fukuda, K. Iwatsuki-Horimoto, Y. Guan, W. Lim, M. Peiris, S. Sugii, T. Odagiri, M. Tashiro and Y. Kawaoka. 2004. Antigenic differences, between H5N1 human influenza viruses isolated in 1997 and 2003. J. Vet. Med. Sci. 66:303-305.
Ito, T., H. Goto, E. Yamamoto, H. Tanaka, M. Takeuchi, M. Kuwayama, Y. Kawaoka and K. Otsuki. 2001. Generation of a highly pathogenic avian influenza A virus from an avirulent field isolate by passaging in chickens. J. Virol. 75:4439-4443.
Jung, K. and C. Chae. 2004. Phylogenetic analysis of an H1N2 influenza A virus isolated from a pig in Korea. Arch. Virol. 149:1415-1422.
Juyal, R., M. Osmamy, R.E. Black, U. Dhingra, A. Sarkar, P. Dhingra, P. Verma, D. Marwah, R. Saxsena, V.P. Menon and S. Sazawal. 2004. Efficacy of micronutrient fortification of milk on morbidity in pre-school children and growth- a double blind randomized controlled trial. Asia Pac. J. Clin. Nutri. 13:44S.
Keawcharoen, J., K. Oraveerakul, T. Kuiken, R.A. Fouchier, A. Amonsin, S. Payungporn, S. Noppornpanth, S. Wattanodorn, A. Theambooniers, R. Tantilertcharoen, R. Pattanarangsan, N. Arya, P. Ratanakorn, D.M. Osterhaus and Y. Poovorawan. 2004. Avian influenza H5N1 in tigers and leopards. Emerg Infect Dis. 10:2189-91.
Koopmans, M.B. Wilbrink, M. Conyn. G. Natrop, H van der Nat, H. Vennema, A. Meijer, J. van Steenbergen, R. Fouchier, A. Osterhaus and A Bosman. 2004.
Transmission of H7N7 avian influenza A virus to human beings during a large outbreak in commercial poultry farms in the Netherlands. Lancet 363:587-593.
Kuiken, T., G. Rimmelzwaan, D. van Riel, G. van Amerongen, M. Baars, R. Fouchier and A. Osterhaus. 2004. Avian influenza in cats. Science 306:241.
Lang, G., A. Gagnon and J.R. Geraci. 1981. Isolation of an influenza A virus from seals. Arch. Virol. 68:189-195.
Li, S., M. Orlich and R. Rott. 1991. Generation of seal influenza virus variants pathogenic for chickens because of hemagglutinin cleavage site changes. J. Virol. 64:3297-3303.
Li, Z., H. Chen, P. Jiao, G. Deng, G. Tian, Y. Li, E. Hoffmann, R.G. Webster, Y. Matsuoka and K. Yu. 2005. Molecular basis of replication of duck H5N1 influenza viruses in a mammalian mouse model. J. Virol. 79:12058-12064.
Liem, N.T. and W. Lim. 2005. Lack of H5N1 avian influenza transmission to hospital employees, Hanoi, 2004. Emerg. Infect. Dis. 11:210-215.
Look, M.P., J.K. Rockstroh, G.S. Rao, K.A. Kreuzer, U. Spengler and T. Sauerbruch. 1997. Serum selenium versus lymphocyte subsets and markers of disease progression and inflammatory response in human immunodeficiency virus-infection. Biol. Trace Elem. Res. 56:31-41.
Lyons, G., J. Stangoulis and R. Graham. 2003. Nutriprevention of disease with high selenium wheat. J. Australasian Coll. Nutri. Environ. Med. 22:3-9.
Mase, M., T. Imada, Y. Sanda, M. Etoh, N. Sanda, K. Tsukamoto, Y. Kawaoka and S. Yamaguchi. 2001. Imported parakeets harbor H9N2 influenza A viruses that are genetically closely related to those transmitted to humans in Hong Kong. J. Virol. 75:3490-3494.
Mase, M., M. Etoh, Tanimura, K. Imai, K. Tsuamoto, T. Horimoto, Y. Kawaoka and S. Yamaguchi. 2005a. Isolation of a genotypically unique H5N1 influenza virus from duck meat imported into Japan from China. Virol. 101-109.
Mase, M.K. Tsukamoto, T. Imada, N. Tanimura, K. Nakamura, Y. Yamamoto, T. Hitomi. T. Kira, T. Nakai, M. Kiso, T. Horimoto, Y, Kawaoka and S. Yamaguchi. 2005b. Characterization of H5N1 influenza A viruses isolated during the 2003-2004 influenza outbreaks in Japan. Virol. 332(1):167-176.
Munster, V.J., A. Wallensten, C. Baas, G. F. Rimmelzwaan, M. Schutten, B. Olsen, A.D.M.E. Osterhaus and R.A.M. Fouchier. 2005. Mallards and highly pathogenic avian influenza viruses, Northern Europe. Emerg. Infect. Dis. 10:1545-1551.
Murphy, B.R. and R.G. Webster. 1996. Orthomyxoviruses. In: Virology (Vol. 1) (B.N. Fields, D.M. Knipe and P.M. Howley, eds). Lippincott-Raven, Philadelphia, PA, pp.1397-1445.
Nakatani, H., K. Nakamura, Y. Yamamoto, M. Yamada and Y. Yamamoto. 2005. Epidemiology, pathology, and immunohistochemistry of layer hens naturally affected with H5N1 highly pathogenic avian influenza in Japan. Avian Dis. 49:436-441.
Nelson, H.K., Q. Shi, P. Van Dael, E.J. Schiffrin, S. Blum, D. Barclay, O.A. Levander and M.A. Beck. 2001. Host nutritional selenium status as a driving force for influenza virus mutations. FASEB 15:1846-1848.
Olsen, C.W. 2002. Emergence of novel strains of swine influenza virus in North America. In: Trends in Emerging Viral Infections of Swine (A. Morilla, K.J. Yoon and J.J. Zimmerman, eds). Blackwell Publishing, Ames, IA, pp. 37-43.
Olsen, S.J., K. Ungchusak, L. Sovann, T.M. Uyeki, S.F. Dowell, N.J. Cox, W. Aldis and S. Chunsuttiwat. 2005. Family clustering of avian influenza A (H5N1). Emerg. Infect. Dis. 11:1799-1801.
Peiris, J.S.M., W.C. Yu, C.W. Leung, C.Y., Cheung, W.F. Ng, J.M. Nicholls, T.K. Ng, K.H. Chan, S.T. Lai, W.L. Lim, K.Y. Yuen and Y. Gaun. 2004. Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet 363:617-619.
Perdue, M.L. and D.E. Swayne. 2005. Public health risk from avian influenza viruses. Avian Dis. 49:317-327.
Perkins, L.E. and D.E. Swayne. 2002. Pathogenicity of a Hong Kong-origin H5N1 highly pathogenic avian influenza virus for emu, geese, ducks and pigeons. Avian Dis. 46:53-63.
Reid, A.H., T.G. Fanning, J.V. Hultin and J.K. Taubenberger. 1999. Origin and evolution of the 1918 “Spanish” influenza virus hemagglutinin gene. Proc. Nat. Acad. Sci. USA 96:1651-1656.
Richt, J.A., K.M. Lager, B.H. Janke, R.D. Woods, R.G. Webster and R.J. Webby. 2003. Pathogenic and antigenic properties of phylogenetically distinct reassortant H3N2 swine influenza viruses cocirculating in the United States. J. Clin. Microbiol. 41:3198-3205.
Rimmelzwaan, G.F., D. van Riel, M. Baars, T.M. Bestebroer, G. van Amerongen, R.A.M. Fouchier, A.D. Osterhaus and T. Kuiken. 2006. Influenza A virus (H5N1) infection in cats causes systemic disease with potential novel routes of virus spread within and between hosts. Am. J. Pathol. 168:176-183.
Rott, R. 1992. The pathogenesis determinant of influenza virus. Vet. Microbiol. 33:303- 310.
Sappey, C., S. Legrand-Poels, M. Best-Belpomme, A. Favier, B. Rentier and J. Piette. 1994. Stimulation of glutathione peroxidase activity decreases HIV type 1 activation after oxidative stress. AIDS Res. Human Retrovir. 10:1451-1461.
Senne, D.A., B. Panigrahy, Y. Kawaoka, J.E. Pearson, J. Suss, M. Lipkind, H. Kida, and R.G. Webster. 1996. Survey of the hemagglutinin (HA) cleavage site sequence of H5 and H7 avian influenza viruses: amino acid sequence at the HA cleavage site as a marker of pathogenicity potential. Avian Dis. 40:425-437.
Southeastern Cooperative Wildlife Diseases Study. 2005. Fact Sheet: Highly Pathogenic Avian Influenza Virus H5N1 and Wild Birds. College of Veterinary Medicine, The University of Georgia, Athens, Georgia.
Stallknecht, D.E. and S.M. Shane. 1988. Host range of avian influenza virus in free-living birds. Vet. Res. Commun. 12:125-141.
Steineke, G.A., M. Vey, H. Angliker, E. Shaw, G. Thomas, C. Roberts, H.D. Klenk, and W. Garten. 1992. Influenza virus hemagglutinin with multibasic cleavage site is activated by furin, a subtilisin-like endoprotease. EMBO J. 11:2407-2414.
Stevens, J., A.L. Corper, C.F. Basler, J.K. Taubenberger, P. Palese and I.A. Wilson. 2004. Structure of the uncleaved human H1 hemagglutinin from the extinct 1019 influenza virus. Science 303:1866-1870.
Sturm-Ramirez, K.M., D.J. Hulse-Post, E.A. Govorkova, J. Humberd, P. Seiler, P. Puthavathana, C. Buranathai, T.D. Nguyen, A. Chaisingh, H.T. Long, T.S.P. Naipospos, H. Chen, T.M. Ellis, Y. Guan, J.S.M. Peiris and R.G. Webster. 2005. Are ducks contributing to the endemicity of highly pathogenic H5N1 influenza virus in Asia? J. Virol. 79:11269-11279.
Suarez, D.L., D.A. Senne, J. Banks, I.H. Brown, S.C. Essen, C-W. Lee, R.J. Manvell, C. Mathieu-Benson, V. Moreno, J.C. Pedersen, B. Panigrahy, H. Rojas, E. Spackman and D.J. Alexander. 2004. Recombination resulting in virulence shift in avian influenza outbreak, Chile. Emerg. Infect. Dis. 10:693-699.
Subbarao, K., A. Klimov, J. Katz, H. Regnery, W. Lim, H. Hall, M. Perdue, D. Swayne, C. Bender, J. Huang, M. Hemphill, T. Rowe, M. Shaw, X. Xu, K. Fukuda and N. Cox. 1998. Characterization of an avian influenza A (H5N1) virus isolated from a child with a fatal respiratory illness. Science 279:393-396.
Suzuki, Y. 2005. Sialobiology of influenza: molecular mechanism of host range variation of influenza viruses. Biol. Pharmaceut. Bull. 28:399-408.
Swayne, D.E. 2004. Application of new vaccine technologies for the control of transboundary diseases. Dev. Biol. 119:219-228.
Swayne, D.E. and D.A. Halvorson. 2003. Influenza. In: Diseases of Poultry (Y.M. Saif, H.J. Barnes, A.M. Fadly, J.R. Glisson, L.R. McDougald and D.E. Swayne, eds). 11th Ed. Iowa State University Press, Ames, IA, pp. 135-160.
Swayne, D.E., J.R. Beck and N. Kinney. 2000a. Failure of a recombinant fowl poxvirus vaccine containing and avian influenza hemagglutinin gene to provide consistent protection against influenza in chickens preimmunized with a fowl pox vaccine. Avian Dis. 44:132-137.
Swayne, D.E., M. Garcia, J.R. Beck, N. Kinney and D.L. Suarez. 2000b. Protection against highly pathogenic H5 avian influenza viruses in chickens immunized with a recombinant fowlpox vaccine containing an H5 avian influenza hemagglutinin gene insert. Vaccine 18:1088-1095.
Taylor, J., R. Weinberg, Y. Kawaoka, R.G. Webster and E. Paoletti. 1988. Protective immunity against avian influenza induced by a fowlpox virus recombinant. Vaccine 6:504- 508.
Thanawongnuwech, R., A. Amonsin, R. Tantilertcharoen, S. Damrongwatanapokin, A. Theamboonlers, S. Payungporn, K. Nanthapornphiphat, S. Ratanamungklanon, E. Tunak, T. Songserm, V. Vivatthanavanich, T. Lekdumrongsak, S. Kesdangsakonwut, S. Tunhikorn and Y. Poovorawan. 2005. Probable tiger-to-tiger transmission of avian influenza H5N1. Emerg. Infect. Dis. 11:699-701.
Tiensin, T., P. Chaitaweesub, T. Songserm, A. Chaisingh, W. Hoonsuwan, C. Buranathal, T. Parakamawongsa, S. Premashthira, A. Amonsin, M. Gilbert, M. Nielsen and A. Stegeman. 2005. Highly pathogenic avian influenza H5N1 in Thailand, 2004. Emerg. Infect. Dis. 11:1664-1672.
Uiprasertkul, M., P. Puthavathana, K. Sangsiriwut, P. Pooruk, K. Srisook, M. Peiris, J. M. Nicholls, K. Chokephaibulkit, N. Vanprapar and P. Auewarakul. 2005. Influenza A H5N1 replication sites in humans. Emerg. Infect. Dis. 11:1036-1041.
Webster, R.G., W.J. Bean, O.T. Gorman, T.M. Chambers and Y. Kawaoka. 1992. Evolution and ecology of influenza A viruses. Microbiol. Rev. 56:152-179.
Webster, R.G., Y. Kawaoka, J. Taylor, R. Weinberg and E. Paoletti. 1991. Efficacy of nucleoprotein and haemagglutinin antigens expressed in fowlpox virus as a vaccine for influenza in chickens. Vaccine 9:303-308.
Webster, R.G., M. Peiris, H. Chen and Y. Guan. 2006. H5N1 outbreaks and enzootic influenza. Emerg. Infect. Dis. 12:3-8.
Weis, W.I., A.T. Brünger, J.J. Skehel, D.C. Wiley. 1990. Refinement of the influenza virus hemagglutinin by simulated annealing. J. Mol. Biol. 212:737-761.
White, J.M., L.R. Hoffman, J.H. Arevalo and I.A. Wilson. 1997. Attachment and entry of influenza virus into host cells. Pivotal roles of hemagglutinin. In: Structural Biology of Viruses (W. Chiu, R.M. Burnett and R.L. Garcea, eds). Oxford University Press, NY, pp. 80-104.
Wood, G.W., J.W. McCauley, J.B. Bashiruddin and D.J. Alexander. 1993. Deduced amino acid sequences at the hemagglutinin cleavage site of avian influenza A viruses of H5 and H7 subtypes. Arch. Virol. 130:209-217.
World Health Organization. 2006. Confirmed human cases of avian influenza A (H5N1). http://www.who.int/csr/disease/avian_influenza/country/en/. Accessed January 16, 2006.
Xu, X., K. Subbarao, N. Cox and Y. Guo. 1999. Genetic characterization of the pathogenic influenza A/goose/Guangdong/1/96 (H5N1) virus: similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virol. 261:15-19.
Yasuda, J., K.F. Shortridge, Y. Shimizu and H. Kida. 1991. Molecular evidence for a role of domestic ducks in the introduction of avian H3 influenza viruses to pigs in southern China, where the A/Hong Kong/68 (H3N2) strain emerged J. Gen. Virol. 72:2007-2010.
Yu, M.W., I.S. Horng, Y.C. Chiang, Y.F. Liaw and C.J. Chen. 1999. Plasma selenium levels and the risk of hepatocellular carcinoma among men with chronic hepatitis virus infection. Am. J. Epidemiol. 150:367-374.
Yu, S.Y., Y.J. Zhu and W.G. Li. 1997. Protective role of selenium against hepatitis B virus and primary liver cancer in Qidong. Biol. Trace Elem. Res. 56:117-124.
Yuen, K.Y., P.K. Chan, M. Peiris, D.N. Tsang, T.L. Que, K.F. Shortridge, P.T. Cheung, W.K. To, E.T. Ho, R. Sung and A.F. Cheng. 1998. Clinical features and rapid viral diagnosis of human disease associated with avian influenza A H5N1 virus. Lancet, 351:467-471.
Zitzow, L.A., T. Rowe, T. Morken, W.J. Shieh, S. Zaki and J.M. Katz. 2002. Pathogenesis of avian influenza A (H5N1) viruses in ferrets. J. Virol. 76:4420-9.

















