Protoporphyrin IX in Shell and Cuticle of Brown Shelled Eggs
Published: July 24, 2013
By: S. Samiullah and J.R. Roberts (University of New England)
Eggs were collected from Hy-Line brown flocks aged 33, 50 and 67 wk. Thirty eggs from each flock were analyzed to determine the reliability of MST (MS Technologies, U.K.) cuticle blue stain as an indicator of the presence of cuticle and the effective removal of cuticle by use of an EDTA solution. Another 30 eggs, collected at the same time from each flock, were processed for the quantification of protoporphyrin IX (PP IX) from the eggshell with and without the presence of cuticle. The L*a* components of the colour space system were significantly different among the age groups. There was a high degree of correlation between the extent of MST cuticle blue staining and the amount of cuticle on the eggshell as recorded by scanning electron microscopy. PP IX pigment was quantified by spectrophotometric analysis of digested eggshell solutions. The average amount of PP IX in the shell without cuticle (8.046 x 10-8 g/g of shell) was higher than the amount present in cuticle (1.669 x 10-8 g/g of shell). The amount of PP IX in the cuticle of 1g of shell in the 33 wk eggs was significantly lower than for eggs from the 50 wk and 67 wk flocks whereas, in the shell without cuticle, it was not significantly different among flock ages. PP IX in cuticle, as a percentage of total PP IX in shell was 13% in 33 wk, 20 % in 50 wk and 18 % in 67 wk eggs.
I. INTRODUCTION
Brown, white or tinted eggs are linked to the genotype of the hen. The eggshell colour of brown eggs is a quality aspect for consumers (Curtis et al., 1985; Jones et al., 2010) and may be related to shell ultrastructure (Richards and Deeming, 2001). Eggshell pigment has been shown to have some antimicrobial properties (Ishikawa et al., 2010). Protoporphyrin IX (PP IX) is believed to be the main eggshell pigment, but other pigments such as zinc porphyrin, biliverdin and zinc biliverdin (Kennedy and Vevers, 1976) may contribute to shell colour.
I. INTRODUCTION
Brown, white or tinted eggs are linked to the genotype of the hen. The eggshell colour of brown eggs is a quality aspect for consumers (Curtis et al., 1985; Jones et al., 2010) and may be related to shell ultrastructure (Richards and Deeming, 2001). Eggshell pigment has been shown to have some antimicrobial properties (Ishikawa et al., 2010). Protoporphyrin IX (PP IX) is believed to be the main eggshell pigment, but other pigments such as zinc porphyrin, biliverdin and zinc biliverdin (Kennedy and Vevers, 1976) may contribute to shell colour.
Previous research has suggested that most of the PP IX is located in the cuticle (Miksik et al., 2007). In contrast, however, the study of Nys et al. (1991), which investigated the kinetics of PP IX deposition, found that approx. 75% of the PP IX was laid down in association with the calcareous part of the shell. Therefore, the purpose of the present study was to quantify the amount of PP IX from the cuticle and true shell layer in brown shelled eggs.
II. MATERIALS AND METHODS
Eggs were collected from conventional cage flocks (Hy-Line brown) which were 33, 50 and 67 wk of age. Eggs were divided into two groups and used for the following two investigations.
The first experiment was conducted to verify the reliability of MST cuticle blue dye as an indicator of the presence of cuticle. It also investigated the effectiveness of the use of EDTA to remove the cuticle without eroding into the true shell. Thirty eggs from each age group (33, 50 and 67 week) were included in this study. Shell colour (L*a*) was measured before staining using a Konica Minolta spectrophotometer (CM-2600d). “L*” has a maximum of 100 (white) and a minimum of 0 (black). For “a*”, green is towards the negative end of the scale and red towards the positive end. Eggs were soaked in MST Cuticle blue dye for 1 minute and rinsed in distilled water to remove excess stain. After drying, shell colour (L*a*) was measured on the stained eggs. For the thorough removal of the cuticle, the method described by Leleu et al. (2011) was followed with slight modification. Each egg, individually, was soaked in an EDTA solution (0.34 M, pH 7.5) for 5 minutes and the cuticle was carefully scrubbed off in running tap water using a small soft brush. Shell colour (L*a*) was measured on the eggs without cuticle. Eggs were restained as described earlier and shell colour (L*a*) was again measured. A small piece from around the equator of the shell was cut out, mounted on an aluminium stub using silver paint, sputter coated with gold for 5 min in a Neocoater (Nikon, Japan) and viewed under a Scanning Electron microscope (SEM) at various magnifications.
The second experiment measured the amount of protoporphyrin IX (PP IX) in the eggshell. Eggs from the 33, 50 and 67 week flocks were analyzed for PP IX in whole eggshell including the cuticle as well as in eggshell from which the cuticle had been removed. A method described by Poole (1965) was used with some modification. Following removal of egg contents via a hole at the blunt end of the eggshell, a clean thin stick was passed across the long axis of the eggshell in such a way as to immerse one longitudinal half of the shell in a glass container of 0.34 M EDTA (pH 7.5) for 5 minutes at the same time as maintaining the other half outside the solution. The cuticle of the soaked longitudinal half side of the eggshell was washed away in running tap water. The eggshell was cut off longitudinally into two equal halves; one having cuticle and one without cuticle. A 0.250 g sample from the equator of each dried shell (without shell membrane) was weighed into a clean 10 mL plastic centrifuge tube into which 4 mL of methanol- concentrated HCl (2:1) solvent was added. All the tubes were wrapped in aluminium foil and placed in a refrigerator for 12 hours. The samples were centrifuged at 3000 rpm for 1 hr. After centrifugation, the supernatant solution was decanted into spectrophotometer cuvettes (4mL) and the absorbance of the supernatant read at 412 nm (Shimadzu, UV-1201). In order to confirm that the shell had dissolved completely, the sediment remaining in the bottom of the centrifuge tubes was viewed under a light microscope at various magnifications.
II. MATERIALS AND METHODS
Eggs were collected from conventional cage flocks (Hy-Line brown) which were 33, 50 and 67 wk of age. Eggs were divided into two groups and used for the following two investigations.
The first experiment was conducted to verify the reliability of MST cuticle blue dye as an indicator of the presence of cuticle. It also investigated the effectiveness of the use of EDTA to remove the cuticle without eroding into the true shell. Thirty eggs from each age group (33, 50 and 67 week) were included in this study. Shell colour (L*a*) was measured before staining using a Konica Minolta spectrophotometer (CM-2600d). “L*” has a maximum of 100 (white) and a minimum of 0 (black). For “a*”, green is towards the negative end of the scale and red towards the positive end. Eggs were soaked in MST Cuticle blue dye for 1 minute and rinsed in distilled water to remove excess stain. After drying, shell colour (L*a*) was measured on the stained eggs. For the thorough removal of the cuticle, the method described by Leleu et al. (2011) was followed with slight modification. Each egg, individually, was soaked in an EDTA solution (0.34 M, pH 7.5) for 5 minutes and the cuticle was carefully scrubbed off in running tap water using a small soft brush. Shell colour (L*a*) was measured on the eggs without cuticle. Eggs were restained as described earlier and shell colour (L*a*) was again measured. A small piece from around the equator of the shell was cut out, mounted on an aluminium stub using silver paint, sputter coated with gold for 5 min in a Neocoater (Nikon, Japan) and viewed under a Scanning Electron microscope (SEM) at various magnifications.
The second experiment measured the amount of protoporphyrin IX (PP IX) in the eggshell. Eggs from the 33, 50 and 67 week flocks were analyzed for PP IX in whole eggshell including the cuticle as well as in eggshell from which the cuticle had been removed. A method described by Poole (1965) was used with some modification. Following removal of egg contents via a hole at the blunt end of the eggshell, a clean thin stick was passed across the long axis of the eggshell in such a way as to immerse one longitudinal half of the shell in a glass container of 0.34 M EDTA (pH 7.5) for 5 minutes at the same time as maintaining the other half outside the solution. The cuticle of the soaked longitudinal half side of the eggshell was washed away in running tap water. The eggshell was cut off longitudinally into two equal halves; one having cuticle and one without cuticle. A 0.250 g sample from the equator of each dried shell (without shell membrane) was weighed into a clean 10 mL plastic centrifuge tube into which 4 mL of methanol- concentrated HCl (2:1) solvent was added. All the tubes were wrapped in aluminium foil and placed in a refrigerator for 12 hours. The samples were centrifuged at 3000 rpm for 1 hr. After centrifugation, the supernatant solution was decanted into spectrophotometer cuvettes (4mL) and the absorbance of the supernatant read at 412 nm (Shimadzu, UV-1201). In order to confirm that the shell had dissolved completely, the sediment remaining in the bottom of the centrifuge tubes was viewed under a light microscope at various magnifications.
A standard stock solution was prepared by dissolving 0.0018 g of powder PP IX disodium salt (Sigma Aldrich Australia) in 30 mL methanol-concentrated HCl (2:1) solvent. Serial dilutions were prepared until a 1:128 dilution was reached and their absorbance was read in a spectrophotometer. A standard curve was constructed by plotting the concentrations of protoporphyrin in the standards diluted 1:16 (6.87 x 10-6 mM), 1:32 (3.43 x 10-6 mM), 1:64 (1.72 x 10-6 mM), and 1:128 (8.59 x 10-7 mM) from the stock solution against the absorbance reading for each standard. The absorbance values were converted into concentration of PP IX in the sample solvent in mmol/L and the amount or protoporphyrin in 1 g of eggshell (with and without cuticle present) was calculated. For determination of the amount of PP IX in the cuticle, the values of the eggshell samples without cuticle were subtracted from the values of the eggshell samples with the cuticle still present. Data were analyzed using Statview Software (SAS Institute Inc., Version 5.0.1.0).
III. RESULTS
In Experiment 1, the Specular Component Included (SCI) L* component of the L*a* space system was not significantly (P=0.1860) affected by flock age when the cuticle was intact. However, following the removal of the cuticle, there were differences among flocks with values being highest for the 50 wk flock. There was a significant effect of cuticle treatment on mean values of L* among each age group. Staining of eggs with intact cuticle resulted in lower L* values whereas cuticle removal increased these values. There was a significant effect of flock age on the SCI a* reading for all cuticle treatments with the 67 wk flock having the highest values. There was also a significant effect of cuticle treatment on values for SCI a* for all flocks. The a* reading decreased following staining in eggs with intact cuticle. Removal of the cuticle with EDTA resulted in lower readings than for shells with intact cuticle. Staining with cuticle blue dye of eggs with cuticle removed resulted in reduction in the a* value for all flocks. A high correlation between the presence of cuticle blue stain and the amount of cuticle as viewed under the scanning electron microscope was recorded. Eggs with good quality intact cuticle stained well; eggs with patchy cuticle acquired patchy stain whereas, in the absence of the cuticle, the eggs did no stain at all. SEM observations of the restained eggshell after cuticle was removed by EDTA confirmed that, in the absence of cuticle, eggshells did not stain with cuticle blue dye.
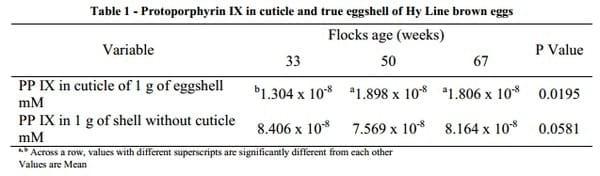
In Experiment 2, for a 1 g piece of eggshell, there was more PP IX in the shell with cuticle intact, as compared with a piece of shell from the same egg with cuticle removed. When the difference between the two was calculated, there was more protoporphyrin present in the true (calcareous) shell than in the cuticle from the same amount of shell, as shown in Table 1. The total amount of PP IX in 1 g of shell with cuticle intact was not significantly different among the flocks. The total amount of PP IX in 1 g of shell without cuticle was not significantly different among the three flocks, although it tended to be lowest in the 55 week and highest in 33 week flock with the 67 week flock intermediate. However, when the amount of PP IX in the cuticle alone of 1 g of total eggshell was calculated, it was significantly higher (P<0.02) in the 50 and 67 week flocks as compared with the 33 week flock. For a given weight of whole eggshell, the percentage of total PP IX found in the cuticle was 13, 20 and 18% in 33, 50 and 67 wk flock eggs, respectively. Microscopic observations of the digested shell precipitates showed only shell membranes which confirmed that all shells had been dissolved in the solvent.
III. RESULTS
In Experiment 1, the Specular Component Included (SCI) L* component of the L*a* space system was not significantly (P=0.1860) affected by flock age when the cuticle was intact. However, following the removal of the cuticle, there were differences among flocks with values being highest for the 50 wk flock. There was a significant effect of cuticle treatment on mean values of L* among each age group. Staining of eggs with intact cuticle resulted in lower L* values whereas cuticle removal increased these values. There was a significant effect of flock age on the SCI a* reading for all cuticle treatments with the 67 wk flock having the highest values. There was also a significant effect of cuticle treatment on values for SCI a* for all flocks. The a* reading decreased following staining in eggs with intact cuticle. Removal of the cuticle with EDTA resulted in lower readings than for shells with intact cuticle. Staining with cuticle blue dye of eggs with cuticle removed resulted in reduction in the a* value for all flocks. A high correlation between the presence of cuticle blue stain and the amount of cuticle as viewed under the scanning electron microscope was recorded. Eggs with good quality intact cuticle stained well; eggs with patchy cuticle acquired patchy stain whereas, in the absence of the cuticle, the eggs did no stain at all. SEM observations of the restained eggshell after cuticle was removed by EDTA confirmed that, in the absence of cuticle, eggshells did not stain with cuticle blue dye.
In Experiment 2, for a 1 g piece of eggshell, there was more PP IX in the shell with cuticle intact, as compared with a piece of shell from the same egg with cuticle removed. When the difference between the two was calculated, there was more protoporphyrin present in the true (calcareous) shell than in the cuticle from the same amount of shell, as shown in Table 1. The total amount of PP IX in 1 g of shell with cuticle intact was not significantly different among the flocks. The total amount of PP IX in 1 g of shell without cuticle was not significantly different among the three flocks, although it tended to be lowest in the 55 week and highest in 33 week flock with the 67 week flock intermediate. However, when the amount of PP IX in the cuticle alone of 1 g of total eggshell was calculated, it was significantly higher (P<0.02) in the 50 and 67 week flocks as compared with the 33 week flock. For a given weight of whole eggshell, the percentage of total PP IX found in the cuticle was 13, 20 and 18% in 33, 50 and 67 wk flock eggs, respectively. Microscopic observations of the digested shell precipitates showed only shell membranes which confirmed that all shells had been dissolved in the solvent.
IV. DISCUSSION
In Experiment 1, as expected, a significant change in L*a* components of the colour space system with the various cuticle treatments confirmed the effectiveness of the procedure used for cuticle removal. For unstained eggs with cuticle intact, there was no difference among the flocks for L* values. Following cuticle removal (but prior to staining), L* values were highest for the 50 wk flock. The SCI a* value of eggshells stained with MST cuticle blue dye is the most important indicator of the amount of cuticle present on an eggshell. The higher SCI a* values for +cuticle, +stain eggs in the 67 wk flock indicated the presence of less cuticle. When cuticle blue stain was applied to shells with cuticle removed, all flocks showed slight reductions in a* values, indicating a small amount of staining which was occurring where cuticle was present in crevices and pores. Experiment 1 also verified that the MST cuticle blue dye is a reliable indicator of the presence of cuticle on eggshells. SEM of shells with cuticle removed confirmed that the EDTA treatment reliably removed the cuticle without eroding the calcium carbonate of the eggshell.
In Experiment 1, as expected, a significant change in L*a* components of the colour space system with the various cuticle treatments confirmed the effectiveness of the procedure used for cuticle removal. For unstained eggs with cuticle intact, there was no difference among the flocks for L* values. Following cuticle removal (but prior to staining), L* values were highest for the 50 wk flock. The SCI a* value of eggshells stained with MST cuticle blue dye is the most important indicator of the amount of cuticle present on an eggshell. The higher SCI a* values for +cuticle, +stain eggs in the 67 wk flock indicated the presence of less cuticle. When cuticle blue stain was applied to shells with cuticle removed, all flocks showed slight reductions in a* values, indicating a small amount of staining which was occurring where cuticle was present in crevices and pores. Experiment 1 also verified that the MST cuticle blue dye is a reliable indicator of the presence of cuticle on eggshells. SEM of shells with cuticle removed confirmed that the EDTA treatment reliably removed the cuticle without eroding the calcium carbonate of the eggshell.
The results of Experiment 2 showed that there is more pigment (protoporphyrin IX) in the calcareous components of the eggshell than in the cuticle of commercial brown eggs. These results are in contrast to previous research that reports more pigment in the cuticle than in the calcareous layers of the eggshell (Miksik et al., 2007; Wang et al., 2009). However, the results of the present study are consistent with the findings of Nys et al. (1991) who reported that 75% of the protoporphyrin was found in the calcareous layers of the shell of brown eggs. In the present study, the amount of protoporphyrin IX in the eggshell with the cuticle removed (P=0.0581) and shell with the cuticle (P=0.4363) was not significantly different among the three age group eggs which supports the suggestion of Odabasi et al. (2007) of a constant rate of secretion of PP IX in the shell gland throughout the production cycle of laying hen. However, there was a statistically significant (P=0.0195) difference among the three flocks in the amount of PP IX in the cuticle alone. A reduced amount of PP IX in the cuticle of the shell does not necessarily indicate reduced cuticle cover as, in the current study, the 33 week flock eggs, which had the lower amount of PP IX in per gram of shell compared to 67 wk, showed more cuticle than 67 week eggs, measured by spectrophotometer (L*a*b* space system). Our finding that there is a greater percentage of total pigment in the calcareous part of the shell than in the cuticle raises questions about the stages of eggshell formation and the times at which protoporphyrin deposition is maximal. Most authors have suggested that pigment is secreted in the last hour of oviposition and deposited in the cuticle (Kennedy and Vevers, 1976; Poole, 1965).
ACKNOWLEDGMENTS: This study was supported by funding from Australian Egg Corporation Limited.
ACKNOWLEDGMENTS: This study was supported by funding from Australian Egg Corporation Limited.
REFERENCES
Curtis PA, Gardner FA, Mellor DB (1985) Poultry Science 64, 302-306.
Ishikawa S, Suzuki K, Fukuda E, Arihara K, Yamamoto Y, Mukai T, Itoh M (2010) FEBS Letters 584, 770-774.
Jones DR, Musgrove MT, Anderson KE, Thesmar HS (2010) Poultry Science 89, 582-587.
Kennedy GY, Vevers HG (1976) Comparative Biochemistry and Physiology 55B, 117-123.
Leleu S, Bain M, Herman L, Heyndrickx M, De Baerdemaeker J, Michiels CW, Perianu C,
Messens W (2011) Proceedings of the European Symposium on the Quality of Eggs and Egg Products, Leipzig, Germany, 2011.
Miksik I, Eckhardt A, Sedlakova P, Mikulikova K (2007) Connective Tissue Research 48, 1-8.
Nys Y, Zawadzki J, Gautron J, Mills AD (1991) Poultry Science 70, 1236-1245.
Odabasi AZ, Miles RD, Balaban MO, Porier KM (2007) Poultry Science 86, 356-363.
Poole HK (1965) Proceedings of the Society of Experimental Biology and Medicine 119, 547-551.
Richards PDG, Deeming DC (2001) British Poultry Science 42, 338-343.
Wang XL, Zheng JX, Ning ZH, Qu LJ, Xu GY, Yang N (2009) Poultry Science 88, 1485-1492.
Curtis PA, Gardner FA, Mellor DB (1985) Poultry Science 64, 302-306.
Ishikawa S, Suzuki K, Fukuda E, Arihara K, Yamamoto Y, Mukai T, Itoh M (2010) FEBS Letters 584, 770-774.
Jones DR, Musgrove MT, Anderson KE, Thesmar HS (2010) Poultry Science 89, 582-587.
Kennedy GY, Vevers HG (1976) Comparative Biochemistry and Physiology 55B, 117-123.
Leleu S, Bain M, Herman L, Heyndrickx M, De Baerdemaeker J, Michiels CW, Perianu C,
Messens W (2011) Proceedings of the European Symposium on the Quality of Eggs and Egg Products, Leipzig, Germany, 2011.
Miksik I, Eckhardt A, Sedlakova P, Mikulikova K (2007) Connective Tissue Research 48, 1-8.
Nys Y, Zawadzki J, Gautron J, Mills AD (1991) Poultry Science 70, 1236-1245.
Odabasi AZ, Miles RD, Balaban MO, Porier KM (2007) Poultry Science 86, 356-363.
Poole HK (1965) Proceedings of the Society of Experimental Biology and Medicine 119, 547-551.
Richards PDG, Deeming DC (2001) British Poultry Science 42, 338-343.
Wang XL, Zheng JX, Ning ZH, Qu LJ, Xu GY, Yang N (2009) Poultry Science 88, 1485-1492.
Get ready for the 2014 edition, have a look!
Content from the event:
Related topics:
Authors:

University of New England
Recommend
Comment
Share
CALPIS AMERICA
14 de diciembre de 2018
Do you have any information on the MST cuticle blue dye?
Recommend
Reply

Would you like to discuss another topic? Create a new post to engage with experts in the community.







.jpg&w=3840&q=75)