INTRODUCTION
Cool temperature storage of eggs prior to incubation is a frequent practice of commercial broiler hatcheries. However, continued storage beyond 7 d leads to an increase in early embryonic mortality. The cellular and molecular mechanisms responsible for early embryonic mortality associated with egg storage and means to reduce such mortality has been addressed by several investigators (Fasenko, 2007; Hamidu et al., 2011; Dymond et al., 2013). Although physical changes to the egg components (reduced yolk and albumen quality) are associated with egg storage and may impact hatchability (Reijrink et al., 2008), it has been suggested that early embryonic mortality due to egg storage is due to the lack of some critical number of viable blastodermal cells at the time the stored eggs are set in an incubator (Fasenko, 2007; Reijrink et al., 2008; Hamidu et al., 2011).
Blastodermal cell loss during egg storage has been attributed to apoptosis and necrosis (Hamidu et al., 2010; Hamidu et al., 2011; Dymond et al., 2013). Histological ramifications of egg storage to the blastoderm include the partial collapse of the subgerminal cavity and compaction of the epiblast cells forming the blastoderm (Bakst and Akuffo, 2002). Bloom et al. (1998) was the first to associate apoptosis with stress-induced blastodermal cell death. Hamidu et al. (2011) compared gene expression profiles from pooled blastoderms (40 per treatment) isolated from eggs stored for either 4 or 14 d and observed that selected pro-apoptotic genes were up-regulated as the duration of storage increased. Alternatively, when eggs subjected to the same storage treatments were incubated for 6 d, the same genes were down-regulated (Hamidu et al., 2011).
Preheating eggs either before the eggs were placed in cool storage (Fasenko, 2007; Reijrink et al., 2009; Gharib, 2013) or one to 4 times in the course of cool storage (Nicholson et al., 2013; Dymond et al., 2013) reduced but did not eliminate the inimical effects of

prolonged egg storage on embryo viability and subsequent hatchability. Dymond et al. (2013) suggested that the periodic preheating during cool egg storage on the blastoderm may reset the storage time perceived by the blastoderm and/or possibly provide an opportunity for blastodermal cells to perform basic cell function and functions otherwise suppressed during egg storage. Regardless of how, the beneficial effects of preheating eggs prior to incubation on hatchability appears to be due to the advancement of the developmental stage of the blastoderm during egg storage either to hypoblast formation when performed prior to cool egg storage (Fasenko, 2007), or early primitive streak formation (Dymond et al., 2013) when performed during egg storage.
It is known that certain pro-apoptotic and antiapoptotic genes are modulated during egg storage and after hatching (Hamidu et al, 2011). However, other gene pathways no doubt are influenced by egg storage. In the following study, the relative expression of selected genes associated with apoptosis, fatty acid metabolism, and oxidative stress were compared to blastoderms isolated from eggs that were or were not subjected to short periods of incubation during egg storage (SPIDES) over a 21-day period. Such information should provide insights both into the relative expression of selected genes associated with these pathways and how SPIDES treatment may ameliorate the inimical effects of prolonged cool egg storage on embryos.
MATERIALS AND METHODS
Eggs and Egg Treatments
In July 2014 (Trial 1) and February 2015 (Trial 2), Ross broiler eggs laid by 37- and 40-week-old hens, respectively, were shipped from a commercial egg depot overnight in a refrigerated truck (16°C) to the Beltsville Agricultural Research Center’s hatchery and placed in the hatchery’s walk-in cool room (16 to 17°C and 75% humidity) within 3 d of oviposition. Eggs were randomly selected and then placed on plastic incubation trays and assigned treatment groups (see Table 1 for descriptions of treatments). In Trial 2, an additional control treatment group was added that consisted of eggs stored for 4 d and then incubated for 10 h to approximate the embryonic stage of development observed in embryos following SPIDES treatments in Trial 1.
Details regarding egg storage, incubation, and hatching can be found in Dymond et al. (2013). Briefly, egg storage during treatments was in a walk-in cool room maintained at 16 to 17°C and 75% RH. Eggs were incubated at 37.5°C and 60% RH. After 19 d of incubation, eggs were transferred to a hatcher maintained at 37.0°C and 75% RH. To verify that the SPIDES treatment successfully augmented the hatchability of stored eggs, candling and true fertilities, and hatch of fertiles and hatch of eggs set were determined. This research met the guidelines approved by the Beltsville Area Animal Care and Use Committee.
Embryo Isolation and Staging
Specific details regarding blastoderm isolation and staging are found in Dymond et al., (2013) and Gupta and Bakst (1993). RNase Zap (Ambion, Foster City, CA) was applied to utensils and filter paper rings used to isolate and transfer the embryos from the surface of the ovum to a sterile 60 mm diameter, RNAse free plastic petri dish (Genesee Scientific, San Diego, CA) containing RNAse free PBS (Ambion 10X PBS, Foster City, CA; Hyclone HyPure Water, Logan, UT). Using a stereomicrocope (Nikon SMZ1500, Melville, NY) white and yellow yolk adhering to the ventral surfaces of the perivitelline layer and embryo were carefully removed to expose the ventral surface of the embryo. Embryo staging was based on the normal tables described by Eyal-Giladi and Kochav (1976, EGK) and Hamburger and Hamilton (1951, HH).
Staged blastoderms were transferred by pipette to RNAase free 1.5 mL centrifuge tubes (supplied with the Qiagen RNeasy Micro Kit # 74004) held on ice until there were 5 to 10 embryos per tube. Excess PBS in the tube was removed, the tube was immersed in liquid nitrogen, and the frozen sample was stored at −80°C until RNA extraction. Each tube with 5 to 10 embryos took 30 to 60 min to prepare before freezing.
RNA Extraction
Blastodermal cell RNA was extracted with Qiagen’s RNeasy Micro Kit (#74004) utilizing its MiniElute Columns (Qiagen Inc, Hilden, Germany). For each sample, RNA quantity was determined using a Nanodrop 2000 (Thermo Scientific, Wilmington DE). RNA quality was estimated using an Agilent Bioanalyzer (Agilent Technology, Santa Clara, CA) utilizing Agilent RNA 6000 Nano Reagents chip for loading up to 500 ng/μl of RNA.
cDNA and Real Time PCR
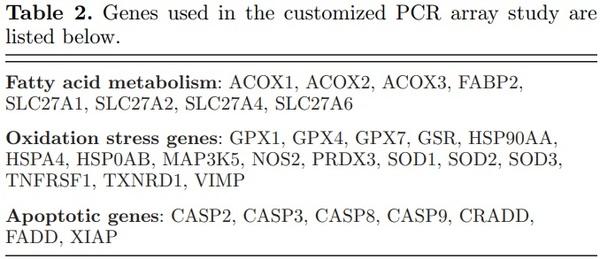
A customized 96-well PCR array for chicken RNA was purchased (Qiagen’s Custom RT2 Profiler PCR Array) using genes selected from its human PCR arrays but limited to those genes for which chicken primers could be designed (proprietary information) by Qiagen (Valencia, CA) (Table 2). One microgram of total RNA was DNase treated with the Genomic DNA elimination mix from the RT2 First Strand Kit, at 42°C for 5 min. Reverse transcription was carried out in a 20 μl volume by addition of 10 μl reverse transcription mix at 42°C
for 15 min and terminated at 95°C for 5 min. The addition of 91 μl RNase free water resulted in a final volume of 111 μl, which was then stored at −20°C before analyzed.
Twenty-five microliters of mastermix of cDNA and RT2 Sybr-Green qPCR Mastermix (Qiagen Valencia, CA), prepared per vendor’s protocol (Anonymous, 2013), was added to each well. Samples were prepared in duplicate. The wells were sealed and the plate was briefly centrifuged. The plate was run using a Real Time PCR system (Bio-Rad CFX96 and associated Bio-Rad CFX Manager software). Thermocycling conditions were as follows: DNA was denatured at 95°C for 10 min and then followed by 40 cycles at 95°C for 15 s, 60°C 1 min. Dissociation curves of the PCR product were produced by heating from 65 to 95°C in increments of 0.5°C every 5 s.
Statistical Analysis
Candling fertility and hatchability data were assessed with Fishers Exact Test for 3 × 2 Table (Freeman and Halton, 1951). Comparison of individual values was made with Fisher’s 2 × 2 Exact Test and P-values adjusted for multiple comparisons with Bonferoni’s correction (McDondald, 2014).
The differences among median values in stages of embryonic development among treatments in each trial were analyzed using Kruska-Wallis Analysis on ranks. Multiple comparisons were made with Dunn’s Method and all procedures were performed with Sigma Plot.
For relative gene expression, analyses were performed by ANOVA and comparisons of the means were made with the Holm-Siddak method. When the data did not meet the standard assumption for normality and variance, analyses were performed by the nonparametric Kruskal-Wallis Test with comparison of means by Dunn’s Test. When there were no differences among the 3 treatments, individual comparisons of means were not made. Relative gene expression was calculated as – ddCt = Log2 (fold changes) (see Livak and Schmittgen, 2001). Positive numbers indicated up-regulation and negative numbers down-regulation with a value of 1 representing about a 2-fold change. All values for relative gene expression were obtained by normalization to Ct values for C1 (unstored and unincubated control eggs). It should be noted that while values of relative gene expression less than 1 or greater than −1 may be
statistically different from 0, the biological significance of such values are questionable.
RESULTS AND DISCUSSION
Fertility and Hatchability
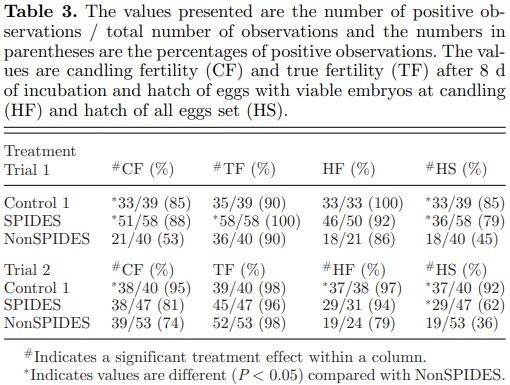
Mean true fertility (TF) of the 3 treatments within Trials 1 and 2 was 93 and 97%, respectively (Table 3). When comparing the hatch of eggs set (HS) among the 3 treatments, control fertility (CF) and fertility after SPIDES treatment were significantly greater than NonSPIDES fertility in both Trials 1 and 2, thus confirming that the SPIDES treatment in this study improved hatchability.
Distribution of Embryonic Stages by Treatment after 8 Days of Incubation
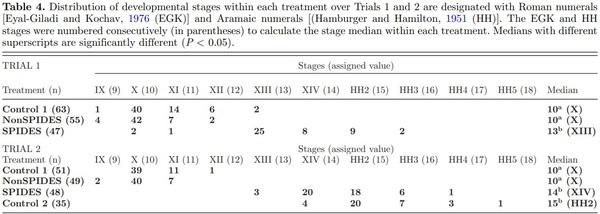
In both trials, Control 1 and NonSPIDES embryos ranged between Stages IX and XIII (Table 4) of development with both pairs of treatments having a median stage value of 10. In contrast, embryo development in the SPIDES eggs advanced to between Stages XIII and HH5 in both trials. The median values of embryo development after SPIDES were 13 in Trial 1 and 14 in Trial 2, both significantly different from the median values of the Control 1 and NonSPIDES treatments. Since there was no significant difference between median values of the stage of development in the Trial 2 SPIDES and Control 2 treatments (Table 4), it is inferred that the differences in relative gene expression between Control 2 and SPIDES embryos were due to the effects of cool egg storage.
Observations Regarding Blastoderm Morphology
After cool eggs storage, the area pellucida (AP) and area opaca (AO) remained discernible in the majority of Stage X blastoderms. Yet, in about 10% of the NonSPIDES blastoderms the AO could not be differentiated from the AP. It is suggested that this appearance is observed when the epiblast collapses into the subgerminal cavity. Bakst and Akuffo (2002) reported that after turkey eggs were stored in a cool room for 14 d, the fluid-filled subgerminal cavity is no longer apparent due to its obliteration by the collapsed epiblast layer. Given that the epiblast layer overlying the fluid-filled subgerminal cavity is visualized as the AP whereas visualization of the AO is a result of the epiblast cells positioned directly on the oolemma and underlying white yolk, a collapsed epiblast layer into the subgerminal cavity eradicates visualization of the AP.
Relative Gene Expression
Apoptotic Genes (Figure 1)Of the 7 apoptotic genes examined (Table 3) only the relative gene expressions of XIAP (X-linked inhibitor of apoptosis), CRADD (death domain protein), and CASP9 (Caspase 9, apoptosis-related cysteine peptidase) were significantly up- or down-regulated relative to 0 (Control 1, Stage X blastoderms). The relative expression of XIAP, an inhibitor of apoptosis by inhibiting the protease caspase, was significantly down-regulated in both SPIDES treatments (embryos at Stage XIV-Stage
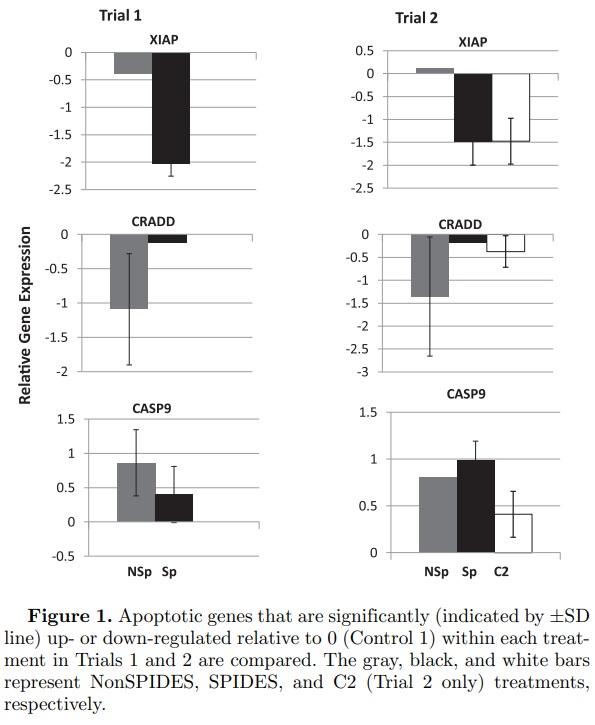
2 HH). That XIAP Control 2 (Stage 2 HH) treatment was also significantly down-regulated suggests that the down-regulation of XIAP in Trials 1 and 2 SPIDES treatments, but not the NonSPIDES treatment, represents the normal stage-specific expression of XIAP. The relative expression of the CRADD gene was significantly down-regulated in the NonSPIDES treatments in the both Trials 1 and 2, and in the Control 2 treatment in Trial 2. Unlike CRADD, and with the exception of the NonSPIDES treatment in Trial 2, the relative expression of CASP9 was significantly up-regulated in both trials within each treatment. The differential expression of the 2 pro-apoptotic genes CRADD and CASP9 may reflect the seemingly conflicting roles of apoptosis both in the normal morpho-differentiation of the embryo and in the response of the embryo to the stresses of prolonged cool egg storage.
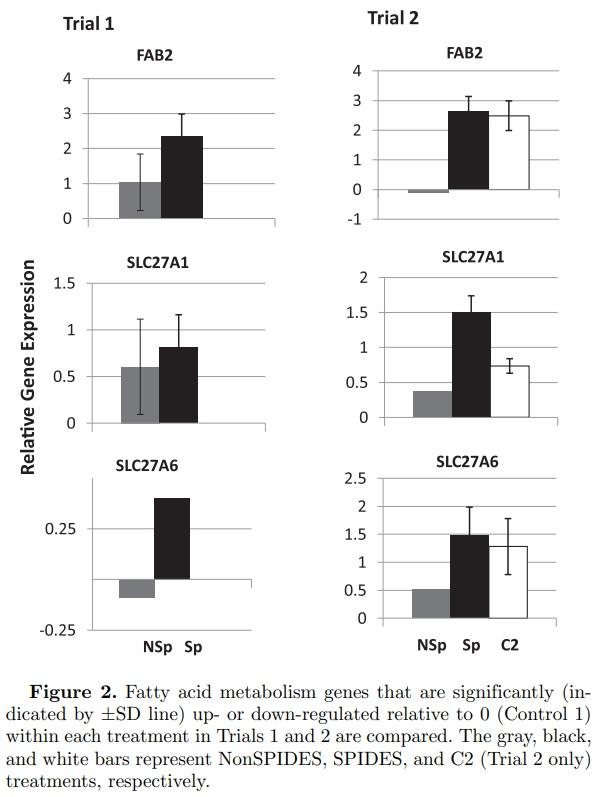
Fatty Acid (FA) Metabolism Genes (Figure 2) Of the 8 genes examined that are associated with FA metabolism (Table 3), FAB2 (fatty acid binding protein, intestinal), SLC27A1 (solute carrier protein 27 [fatty acid transporter], member 1), and SLC27A6 (solute carrier protein 27 [fatty acid transporter], member 6) were significantly up-regulated. In Trial 2, the upregulation of these genes in the SPIDES and Control 1 treatments, but not the NonSPIDES treatment, suggests that the normal stage-specific expression of FAB2, SLC27A1, and SLC27A6 was sustained through egg storage as a result of SPIDES. However, unlike Trial 2, Trial 1 NonSPIDES and SPIDES treatments resulted in a significant up-regulation in the relative expression of FAB2 and SLC27A1, suggesting that prolonged egg storage had little impact on the relative expression of these 2 genes.
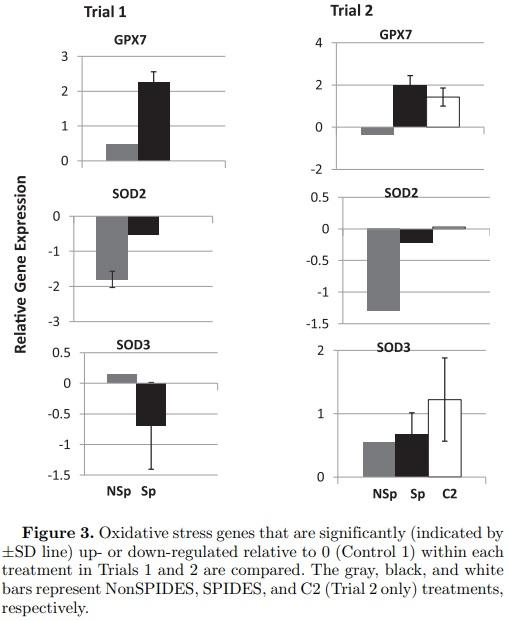
Oxidative Stress Genes (Figure 3) Sixteen oxidation stress genes were examined (Table 3). Only GPX7 (glutathione peroxidase 7) and SOD3 (superoxide dismutase 3, extracellular), both anti-oxidants, were upregulated significantly in both the SPIDES and Control 2 treatments in Trial 2, suggesting that the normal stage-specific expression of both genes was sustained through egg storage as a result of SPIDES. Though the magnitudes of the down-regulation of SOD2 (superoxide dismutase 2, mitochondrial) in the NonSPIDES treatments were similar in both trials, the relative expression of this gene was only significant in the NonSPIDES treatment in Trial 1. In contrast to Trial 2, the relative expression of SOD3 was significantly downregulated in the Trial 1 SPIDES treatment as was SOD2 in the Trial 1 NonSPIDES treatment.
Control 2 Embryos (Figures 1 to 3) In Trail 2, relative gene expressions were significantly up-regulated in 6 genes (Figure 1, CASP9; Figure 2, FAB2, SLC27A1, SLC27A6; Figure 3, GPX7, SOD3) and down regulated in 2 genes (Figure 1 XIAP, CRADD). This demonstrates stage-specific gene transcripts differentially expressed between Stage X (Control 1) and Stages XIV/2 HH (Control 2) embryos.
Prolonged Cool Egg Storage and the Establishment of Embryo Subpopulations As reviewed in the introduction, cool egg storage for 7 to 21 d is detrimental to the physical qualities of the egg, hatchability, and chick or poult quality. Given that a variable number of the stored eggs do hatch with or without SPIDES treatments, (Table 2; Dymond et al., 2013) one can categorize embryos in stored eggs into one of 3 subpopulations at the time of incubation: 1) Embryos that are resistant to the stress of cold storage and hatch as good-quality chicks or poults; 2) Embryos that are impacted by cold storage and hatch as poor-quality chicks or poults; and, 3) Embryos that die prior to or during incubation.
The ramifications of this premise extend into this study and other similar studies (Hamidu et al., 2011) that pool multiple embryos from the same treatment to get a single treatment sample. The relative changes in gene expression observed represents a homogenization of first 2 categories above (the hatching categories): the third category, represented by embryonic mortalities, may have died either due to the responses of the gene pathways described in this study, or most likely, some yet-to-be-identified gene pathway or cellular or molecular complications stemming from cool egg storage. While the data presented in this study may contribute to the understanding of the basis of embryonic mortality and poor chick or poult quality in stored eggs, identifying the specific gene pathways and/or the cellular and molecular mechanisms behind embryo mortality during storage through the initial 8 d off incubation will be challenging.
CONCLUSION
While limited in scope, this study provides a better understanding of both the relative gene expressions associated with embryonic death following prolonged egg storage as well as insight as to how SPIDES reduces embryo mortality rates in stored eggs. Identifying certain stage-specific genes in the Control 2 treatment that appear to be similarly expressed in the SPIDES treatment will be used to further refine the SPIDES protocol, possibly making it more adaptable to other species. The relative gene expression of some genes examined was not what was anticipated but this most likely reflected our lack of understanding of the specific roles of specific genes in the normal development of the early embryo. Finally, protein expression and gene regulated cell activities may not reflect a relative change in RNA expression. MicroRNAs, small, noncoding RNAs that alter the translational efficiency of messenger RNAs have the capacity to influence the expression of mRNA from apoptotic genes (Garofalo et al., 2010).
Future studies in this laboratory will include in-depth profiling of the transcriptomes of broiler embryos subjected to the same treatments used in Trial 2. With this new information, we may be better able to influence the cellular events and gene pathways that impact embryo mortality during egg storage by: genetic selection within breeder lines; temperature/light manipulation during egg storage and incubation; manipulating albumen, yolk/germinal disc composition through the hen’s diet; or by some yet-to-be described method.
This article was originally published in 2016 Poultry Science 95:1411–1417. http://dx.doi.org/10.3382/ps/pew038. This is an Open Access article is under a Creative Commons license.