High quality poultry meat - the ultimate goal. Can trace mineral nutrition have an impact?
Poultry producers have long used preventative measures to ensure that a high yield, disease-free product goes to market. Traditionally, those preventative measures have included proper husbandry, vaccination, antimicrobial feed additives and biosecurity.
The re-evaluation of current drug programs has forced producers to place a larger focus on husbandry practices, specifically those involving nutrition. Trace minerals are involved in metabolic processes ranging from tissue formation to overall immune function. For this reason, the industry is revisiting trace minerals as a nutritional preventative to mortality and morbidity. This review focuses on two specific aspects of quality: bird health and meat quality.
Trace minerals affect bird health and overall immunocompetence
Larsen et al. (1997) determined the effects of selenium (Se) on antibody response to an antigen. White Leghorn chicks of a low antibody strain were used to ensure measurable responsiveness to antigen presentation. Chicks were raised on a standard corn-soybean meal diet to 35 days of age. At 35 days birds were fed diets containing either: 0, 0.2, 0.3, 0.4, 0.6 or 0.8 ppm Se. At 36 days of age chicks were inoculated with sheep red blood cells. Six days post-inoculation, plasma was collected and antibody titers were determined. Researchers noted a 1.4 log2 increase in antibody titers with all levels of selenium inclusion (P< 0.01).
A 4 x 3 factorial treatment design was used to study the effects of trace mineral supplementation at three different states of disease challenge (Hegazy and Adachi, 2000).
Four experimental diets were formulated: basal diet (0.15 ppm selenium (Se) and 40 ppm zinc (Zn); non-medicated), Se- and Zn-fortified diet (basal diet supplemented with 1 and 60 ppm Se and Zn, respectively), Se-fortified diet (basal diet supplemented with 1 ppm Se), and Zn-fortified diet (basal diet supplemented with 60 ppm Zn).
Disease challenge included: aflatoxin and Salmonella typhimurium infection (T1), S. typhimurium infection (T2), and (T3) unchallenged birds. At five days of age, pathogen-free White Leghorn chicks were inoculated intragastrically with 0.5 mL (3.5 x 106 cells) of salmonella inoculum. Aflatoxin B1 (2.6 ppm based on body weight) was injected into the peritoneal cavity 3 days post-salmonella challenge. T1 and T2 groups, which were supplemented with selenium, had improved antibody response compared with other birds given the same disease challenge (Table 1).
| Table 1.Effect of Se, Zn or Se + Zn dietary enhancement on geometric mean of agglutination titers against S. typhimurium during salmonella or salmonella plus aflatoxin inoculation. |
 |
| ab Values within rows differ (P≤0.05) xyz Values within columns differ (P≤0.05) Adapted from Hegazy and Adachi (2000) |
Forty-eight female Nicholas poults were used to determine the effects of trace mineral supplementation on the mononuclear-phagocytic system with and without E. coli challenge (Kidd et al., 1994). Dietary treatments consisted of a commercial control (130 μg Zn/g) and zinc methionine (Zn-Met) supplemented group (control diet with 40 μg Zn-Met/g; 166 μg Zn/g). Both dietary treatments were formulated to have equal levels of Zn-Met and inorganic Zn.
Researchers reported poults receiving the Zn-Met supplemented diet had improved 0 to 21 day weight gain compared with unsupplemented controls. In order to create a harvestable macrophage population, an inflammatory agent (Sephadex® G-50) was injected into the abdominal cavity of nine poults per treatment (on day 20) to promote the migration of inflammatory cells.
Forty-two hours postinjection, poults were killed and abdominal exudate cells (AEC), a direct measure of the bird’s ability to respond to an inflammatory challenge, were collected. Though there was no difference in the number of AEC between treatments, there was an increase in mean adherent cells when Zn-Met was supplemented, which reflects the viability and integrity of the macrophage (Table 2).
In order to measure the effectiveness of enhancing the monuclear-phagocytic system, the ability to clear E. coli from circulation was measured in six poults per treatment. Bacterial clearance was determined by measuring the number of E. coli colony-forming units at regular intervals post intravenous (IV) injection of 1 x 106 E. coli cells. Poults fed Zn-Met diets had improved bacterial clearance over control counterparts (Table 3).
| Table 2.Response to Sephadex® by turkey abdominal macrophages. |
 |
| Adapted from Kidd et al. (1994) |
Trace mineral effects on meat quality: organic vs. inorganic
All of the aforementioned studies confirmed that trace mineral status is an important aspect of immune function, particularly in the context of stress. The studies further imply that current trace mineral recommendations may not be sufficient for today’s market animal.
Organic minerals may allow us to increase trace mineral bioavailability without increasing overall mineral excretion, thereby minimizing environmental impact (Leeson and Sefton, 2003). In terms of meat quality, Mahan et al. (1999) reported a depression in meat quality with excess elemental trace mineral supplementation. The following studies focus on the effects of organic minerals on tissue accretion and mineral incorporation.
| Table 3.Blood clearance of IV-administered E. coli as influenced by diet. |
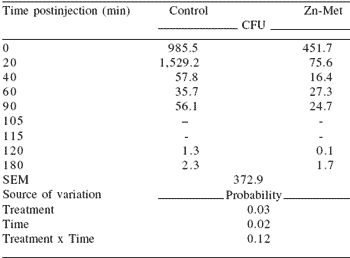 |
| Adapted from Kidd et al. (1994) |
Trace mineral requirements are typically met via feed ingredients. Supplementation usually involves a premix composed of minerals in their elemental salt forms. In contrast, organic minerals are typically chelates composed of peptides or amino acids.
Leeson (2005) stated that inorganic trace minerals have a wide range of availability (40 to 78%) and the addition of a peptide improves the bioavailability of the mineral complex (90 to 95%). The difference in bioavailability is most likely due to how organic minerals are absorbed through the brush border membrane and the increased stability of the complex in the lumen of the gastrointestinal tract. Mineral salts are not readily absorbed and require additional conversion before being transported across the luminal wall.
Selenium (Se), as sodium selenite, is a good example. Selenite absorption relies on passive transport and is incorporated as selenocysteine into functional proteins such as glutathione peroxidase (GSH-Px). Selenomethionine, an organic form of selenium, can also undergo conversion to selenocysteine and subsequent functional protein incorporation.
However, in contrast to selenite, selenomethionine can undergo active absorption through the intestinal wall via the methionine pathway. Once absorbed, selenomethionine can be incorporated nonspecifically into distinct proteins, e.g., muscle (Bou et al., 2005). Studies have found that supplementing sodium selenite will result in higher plasma concentrations of GSH-Px, and less tissue selenium compared with supplementing organic selenium sources (Mahan and Parrett, 1996; Mahan et al., 1999).
This finding suggests that the availability of selenium in tissue would be improved with the use of organic forms of selenium.
Bou and coworkers (2005) utilized a factorial treatment design to evaluate the effect of fat source and mineral supplementation on product nutrient composition. Mineral supplementation included: zinc, as Zn sulfate; selenium, as sodium selenite and Sel- Plex® Se-enriched yeast (Alltech Inc.). Each mineral was supplemented at three levels; Zn (0, 300, or 600 ppm) and Se (0, 1.2 ppm Se from sodium selenite or 0.2 ppm Se from Sel-Plex®).
At 39 days of age, birds were slaughtered and processed according to commercial procedures. Carcasses, including skin, were cut longitudinally and divided into left and right sides. These halves were then deboned, ground, mixed, and frozen for subsequent mineral content analysis. In regard to bird performance, similar weight gain, feed intake, feed conversion and mortality were observed between mineral treatments.
Fat source had no effect on trace mineral concentration of meat. In addition, inorganic zinc had no effect on zinc mineral content of the meat. When broilers were fed a diet supplemented with Sel-Plex® (0.2 ppm Se), meat selenium content increased compared with meat from chickens receiving no selenium supplementation (Table 4).
Table 4.Selenium content of mixed raw dark and white chicken meat with skin (per 100 g of edible portion). |
 |
| ab Values differ (P≤0.05) Adapted from Bou et al. (2005) |
Virden and coworkers (2003) used a randomized complete block design to evaluate the effect of mineral addition on live performance and carcass characteristics of broiler breeder progeny. Five hundred broiler breeders were randomly assigned to one of four dietary treatments: control (75 ppm Zn and 83 ppm manganese (Mn) from inorganic mineral premix), ZnIMnI (control plus 75 ppm Zn from ZnSO4 and 80 ppm Mn from MnSO4), ZnOMnI (control plus 75 ppm Zn from an amino acid complex (organic) and 80 ppm Mn from MnSO4), ZnOMnO (control plus 75 ppm Zn and 80 ppm Mn from amino acid complex).
Breeders were fed treatments in mash form from 21 weeks of age to end of experiment. At 37 weeks of age 1,200 eggs were obtained and set with 960 chicks placed in pens and raised to day 42. All broilers were fed a common diet after placement to evaluate the effect of breeder diet on growth and performance. Overall, supplementing breeder diets with minerals had no effect on progeny performance or carcass characteristics (dressing, breast yield and fat percentage).
However, progeny from breeders fed diets supplemented with organic trace minerals displayed improved livability through day 34 compared with progeny from control breeders (Table 5). The authors theorized that progeny livability improved as a result of enhanced immune function.
| Table 5.Livability of progeny (day 0 to 34) from broiler breeders fed diets differing in Zn and Mn level and source. |
 |
| 1I = Inorganic; O = organic ab Values differ (P≤0.05) Adapted from Virden et al. (2003) |
Chromium (Cr) is a component of the molecular complex, glucose tolerance factor (GTF), which facilitates cellular binding and augments the action of insulin. Ahmed and coworkers (2005) sought to determine whether chromium could augment the utilization of dietary energy, supplementing diets with either inorganic or organic chromium.
Ninety Hubbard broiler chicks were assigned to one of six dietary treatment groups, each consisting of five replicates with three birds per replicate. A BIS (Bureau of Indian Standards, 1992) recommended energy diet and a low energy diet (10% ME reduction) were formulated for starter and finisher periods and fed to 35 days of age. Both BIS and low energy diets were supplemented with either: 0 ppm Cr (Cr-), 0.2 ppm Cr as CrCl3 • 6H2O (Cr+), or 0.2 ppm Cr as CrCl3 • 6H2O with 50 ppm ascorbic acid (CrAA+). The inclusion of ascorbic acid was meant to increase the solubility of inorganic chromium through the formation of a chelate that has been reported to prevent the precipitation of chromium out of luminal contents.
The authors reported that liver, breast and thigh chromium content was increased with chromium supplementation (Table 6). Furthermore, the addition of CrAA+ increased elemental chromium deposition in both breast and thigh tissue compared with Cr+ (P<0.1). Carcass composition (hot carcass, carcass protein and fat, and ash) was also improved by the use of organic chromium, further enhancing carcass composition (P<0.1) (Table 7).
Authors surmised that the inclusion of chromium led to an apparent shift in energy deposition from adipose to lean tissues. Interestingly, birds consuming increased levels of chromium also had lower blood glucose levels suggesting that glucose utilization by adipocytes and muscles cells had become more efficient.
The aforementioned conclusions focused on the effect of chromium addition to BIS energy diets, however, it is worth noting that dietary energy level was a limiting factor in terms of positive impact of chromium supplementation when evaluating carcass composition (fat accretion, breast and thigh weight and percent water-holding capacity). Energy density appeared to be a critical factor that affected performance, but also was important when maximizing the potential of chromium supplementation.
| Table 6. Concentration of chromium (ppm) in the liver and heavy muscles (dry matter basis) of broiler chickens fed a diet supplemented with chromium (CrCl3 • 6H2O) and a combination of chromium and ascorbic acid. |
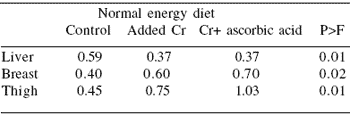 |
| Adapted from Ahmed et al. (2005) |
| Table 7.Carcass composition of broiler chickens fed diets supplemented with chromium (CrCl3 • 6H2O) or a combination of chromium and ascorbic acid. |
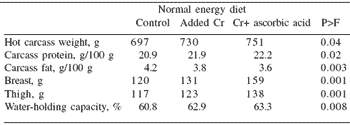 |
| Adapted from Ahmed et al. (2005) |
Implications
Management is beginning to overshadow drug programs in disease prevention and maintenance of bird health. Current standards of trace mineral inclusion may no longer apply to today’s commercial birds. However, although trace minerals are relatively cheap to supplement, high supplementation may lead to environmental concerns regarding excess trace mineral excretion. The increased bioavailability of organic minerals may allow for more efficient incorporation of minerals into tissue, and thus correct for excess mineral excretion. Meat has long been considered a dietary source of trace minerals, however, compared with red meat, poultry products contain lower concentrations. Organic minerals have the potential to increase mineral incorporation into tissue. Development of trace mineral enhanced poultry meat may allow the industry to be more competitive via designer products.
References
Ahmed, N., S. Haldar, M.C. Pakhira and T.K. Ghosh. 2005. Growth performances, nutrient utilization and carcass traits in broiler chickens fed with a normal and a low energy diet supplemented with inorganic chromium (as chromium chloride hexahydrate) and a combination of inorganic chromium and ascorbic acid. J. Agric. Sci. 143:427- 439.
Bou, R., F. Guardiola, A.C. Barroeta and R. Codony. 2005. Effect of dietary fat sources and zinc and selenium supplements on the composition and consumer acceptability of chicken meat. Poult. Sci. 84:1129-1140.
Bureau of Indian Standards. 1992. Nutrient requirements for poultry. In: Animal Feeds and Feeding Stuffs – Determination of Calcium and Magnesium in Mineral Supplements. IS: 13574, New Delhi.
Hegazy, S.M. and Y. Adachi. 2000. Comparison of the effects of dietary selenium, zinc, and selenium and zinc supplementation on growth and immune response between chick groups that were inoculated with Salmonella and aflatoxin or Salmonella. Poult. Sci. 79:331-335.
Kidd, M.T., M.A. Qureshi, P.R. Ferket and L.N. Thomas. 1994. Blood clearance of Escherichia coli and evaluation of mononuclear-phagocytic system as influenced by supplemental dietary zinc methionine in young turkeys. Poult. Sci. 73:1381-1389.
Larsen, C.T., F.W. Pierson and W.B. Gross. 1997. Effect of dietary selenium on the response of stressed and unstressed chickens to Escherichia coli challenge and antigen. Biol. Trace Elem. Res. 58:169-176.
Leeson, S. 2005. Trace mineral requirements of poultry-validity of the NRC recommendations. In: Re-defining Mineral Nutrition (J.A. Taylor-Pickard and L.A. Tucker, eds). Nottingham University Press, UK, pp. 107-117.
Leeson, S. and A. Sefton. 2003. Response of broilers fed organic vs. inorganic trace minerals, Alltech communication.
Mahan, D.C. and N.A. Parrett. 1996. Evaluating the efficacy of selenium-enriched yeast and sodium selenite on tissue selenium retention and serum glutathione peroxidase activity in grower and finisher swine. J. Anim. Sci. 74:2967-2974.
Mahan, D.C., T.R. Cline and B. Richert. 1999. Effects of dietary levels of seleniumenriched yeast and sodium selenite as selenium sources fed to growing-finishing pigs on performance, tissue selenium, serum glutathione peroxidase activity, carcass characteristics, and loin quality. J. Anim. Sci. 77:2172-2179.
Virden, W.S., J.B. Yeatman, S.J. Barber, C.D. Zumwait, T.L. Ward, A.B. Johnson and M.T. Kidd. 2003. Hen mineral nutrition impacts progeny livability. J. Appl. Poult. Res. 12:411-416.
Authors: CURTIS NOVAK and CATALINA TROCHE
Virginia Polytechnic Institute and State University, Blacksburg, Virginia, USA








.jpg&w=3840&q=75)