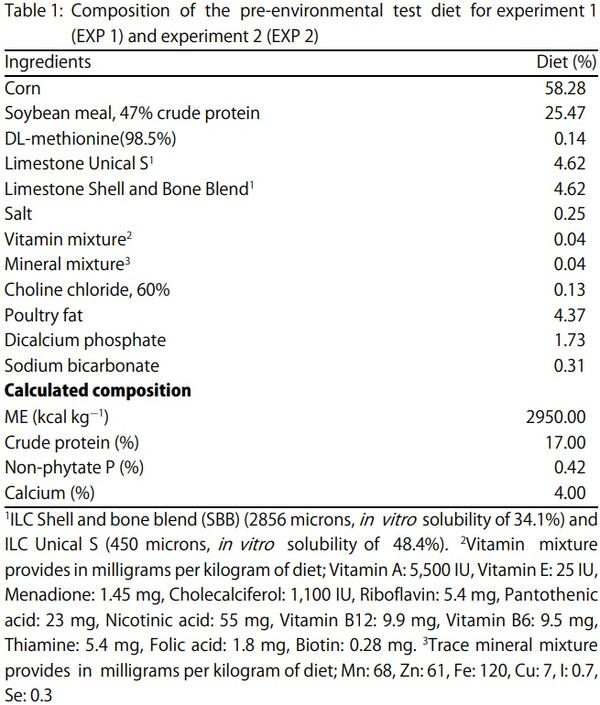
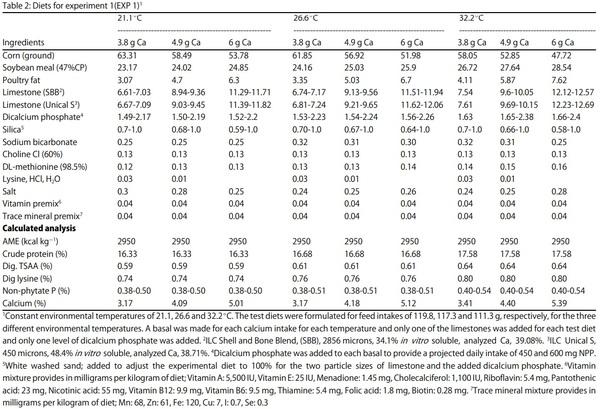
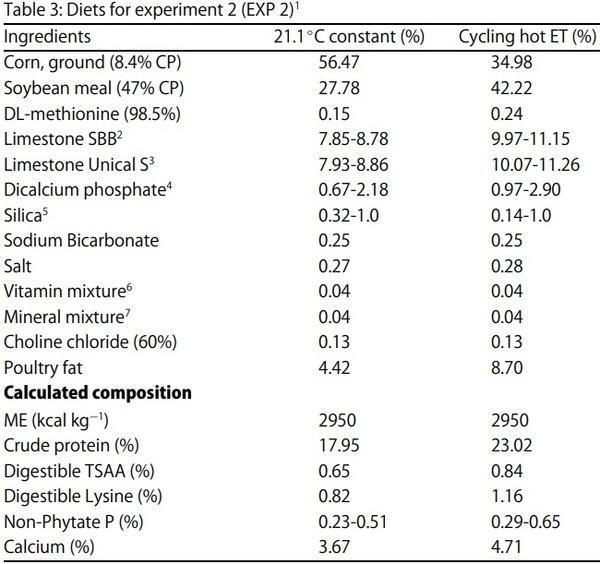
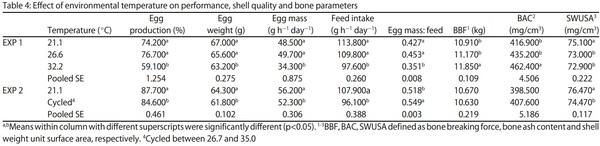
Egg Shell Quality and Bone Status as Affected by Environmental Temperature, Ca and Non-Phytate P Intake and in vitro Limestone Solubility in Single-Comb White Leghorn Hens
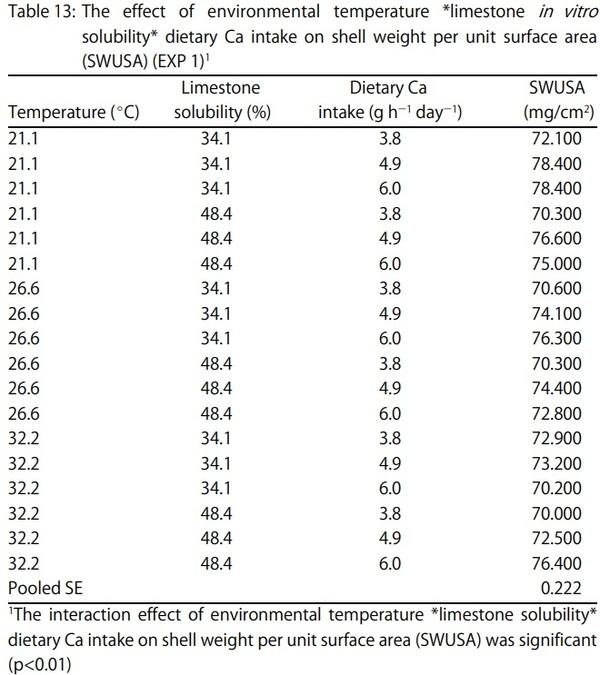
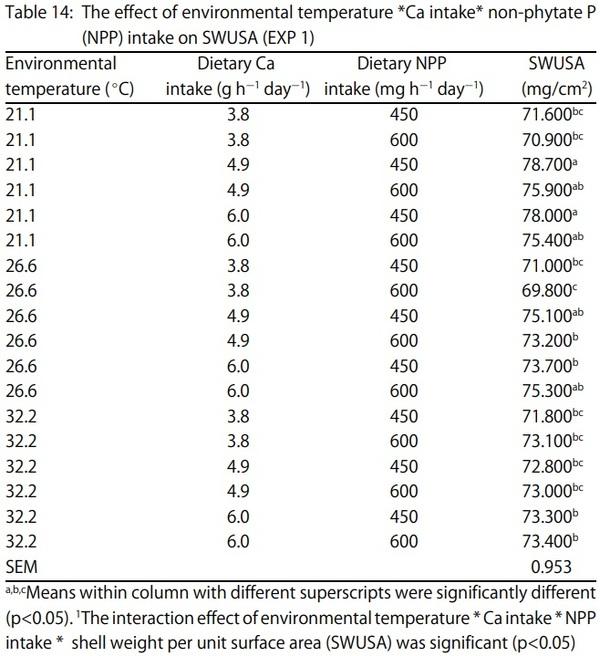
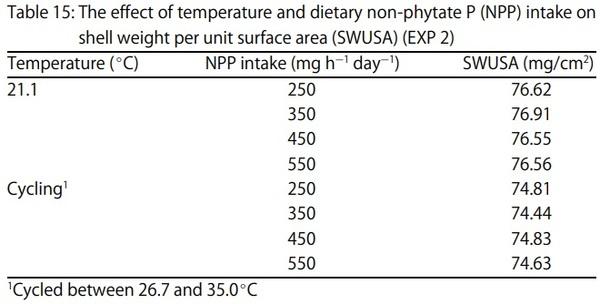
1. Keshavarz, K. and S. Nakajima, 1993. Re-evaluation of calcium and phosphorus requirements of laying hens for optimum performance and eggshell quality. Poult. Sci., 72: 144-153.
2. National Research Council, 1984. Nutrient Requirements of Poultry. 8th Rev. Edn., Washington, DC: The National
Academies Press, United States, Pages: 80.
3. NRC., 1994. Nutritional Requirements of Poultry. 9th Edn.,
National Academy Science, Washington DC., USA.
4. Scheideler, S.E., 1994. Review of the basics of Ca and phosphorus nutrition in the laying hen. Proceedings of the
Midwest Poultry Convention Egg Production Workshop, (MPCEPʼ94), Minnesota, USA.
5. Rostagno, H.S., L.F.T. Albino, M.I. Hannas, J.L. Donzele and
N.K. Sakomuraet al., 2017. Brazilian Tables for Poultry and
Swine. Feedstuff Composition and Nutritional Requirements.
4th Edn., Universidade Federal de Viçosa, Viçosa, Brazil.
6. FEDNA., 2018. Nutritional Needs of Poultry. In: FEDNA
Standards, 2nd Edn., Santoma, G. and G.G. Mateos (Eds.).,
Madrid, Spain.
7. Rodriguez, M., W.J. Owings and J.L. Sell, 1984. Influence of phase feeding available phosphorus on egg production characteristics, carcass phosphorus content and serum inorganic phosphorus levels of three commercial layer strains. Poult. Sci., 63: 1553-1562.
8. Scheideler, S.E. and J.L. Sell, 1986. Effects of calcium and phase-feeding phosphorus on production traits and phosphorus retention in two strains of laying hens. Poult. Sci.,
65: 2110-2119.
9. Said, N.W. and T.W. Sullivan, 1985. A comparison of continuous and phased levels of dietary phosphorus for commercial laying hens. Poult. Sci., 64: 1763-1771.
10. Singsen, E.P., A.H. Spandorf, L.D. Matterson, J.A. Serafin,
J.J. Tlustohowicz, 1962. Phosphorus in the nutrition of the adult hen: 1. Minimum phosphorus requirements. Poult. Sci.,
41: 1401-1414.
11. Singsen, E.P., L.D. Matterson, J.J. Tlustohowicz and
W.J. Pudelkiewicz, 1969. Phosphorus in the nutrition of the adult hen: 2. The relative availability of phosphorus from several sources for caged layers. Poult. Sci., 48: 387-393.
12. Harms, R.H., B.L. Damron and R.D. Miles, 1977. Phosphorus and laying hen nutrition. Part II: Inadequate phosphorus levels result in increased mortality and decreased egg production. Feedstuffs, 49: 33-34.
13. Mikaelian, K.S. and J.L. Sell, 1981. Performance of laying hens fed various phosphorus levels continuously or phase fed decremental phosphorus levels. Poult. Sci., 60: 1916-1924.
14. Sell, J.L., S.E. Scheideler and B.E. Rahn, 1987. Influence of different phosphorus phase-feeding programs and dietary calcium level on performance and body phosphorus of laying hens. Poult. Sci., 66: 1524-1530.
15. Garlich, J.D., 1979. The phosphorus requirements of laying hens. Proceedings of Georgia Nutrition Conference, (GNCʼ79),
Atlanta, GA., 104-114.
16. Charles, O.W., S. Duke and B. Reddy, 1978. Effect of phosphorus source and level on laying hen performance under varying temperature conditions. Georgia Nutr. Conf.,
77: 74-87.
17. Belay, T., C.J. Wiernusz and R.G. Teeter, 1992. Mineral balance and urinary and fecal mineral excretion profile of broilers housed in thermoneutral and heat-distressed environments.
Poult. Sci., 71: 1043-1047.
18. Cheng, T.K., R.L. Jevne and C.N. Coon, 1991. Calcium nutrition studies with layers. Proceedings of the 52nd Minnesota
Nutrition Conference, September 16-18, 1991, Bloomington,
MN., pp: 272-285.
19. Coon, C. and K. Leske, 1999. The effect of soluble calcium levels, nonphytate phosphorus levels and Natuphos phytase on laying hen performance. Proceedings of BASF Technical
Symposium, Use of Natuphos Phytase in Layer Nutrition and
Management, Atlanta, GA., (NPLNMʼ99), BASF Corporation,
Mt. Olive, New Jersey, pp: 62-77.
20. Coon, C., R. Ekmay and M. Manangi, 2011. Faecal, urinary and circulatory phosphorus flow in poultry. University of
Minnesota. Minnesota Extension Service. Retrieved from the
University of Minnesota Digital Conservancy, http://hdl handle.net/11299/204276.
21. Ekmay, R.D. and C.N. Coon, 2011. An examination of the P requirements of broiler breeders for performance, progeny quality and P balance 2. Ca particle size. Int. J. Poult. Sci.,
10: 760-765.
22. Cheng, T.K. and C.N. Coon, 1990. Comparison of various in vitro methods for the determination of limestone solubility. Poult. Sci., 69: 2204-2208.
23. Zhang, B.F. and C.N. Coon, 1997. Improved in vitro methods for determining limestone and oyster shell solubility. J.
Applied Poult. Res., 6: 94-99.
24. Cheng, T.K. and C.N. Coon, 1990. Effect of calcium source, particle size, limestone solubility in vitro and calcium intake level on layer bone status and performance. Poult. Sci.,
69: 2214-2219.
25. Paganelli, C.V., A. Olszowka and A. Ar, 1974. The avian egg:
Surface area, volume and density. Condor, 76: 319-325.
26. AOAC., 1990. Official Methods of Analysis: Association of
Analytical Chemistry. 5th Edn., AOAC Inc., Washington, DC,
USA.
27. Zhang, B. and C.N. Coon, 1997. The relationship of various tibia bone measurements in hens. Poult. Sci., 76: 1698-1701.
28. SAS., 2012. SAS User's Guide Statistics, Version 9.3. SAS
Institute Inc., Cary, NC., USA.
29. De Andrade, A.N., J.C. Rogler and W.R. Featherston, 1976.
Influence of constant elevated temperature and diet on egg production and shell quality. Poult. Sci., 55: 685-693.
30. Marsden, A. and T.R. Morris, 1987. Quantitative review of the effects of environmental temperature on food intake, egg output and energy balance in laying pullets. Br. Poult. Sci.,
28: 693-704.
31. Oguntunji, A.O. and O.M. Alabi, 2010. Influence of high environmental temperature on egg production and shell quality: A review. World's Poult. Sci. J., 66: 739-750.
32. Payne, C.G., 1966. Practical aspects of environmental temperature for laying hens. World's Poult. Sci. J., 22:
126-139.
33. De Andrade, A.N., J.C. Rogler, W.R. Featherston and
C.W. Alliston, 1977. Interrelationships between diet and elevated temperatures (Cyclic and Constant) on egg production and shell quality. Poult. Sci., 56: 1178-1188.
34. Vohra, P., W.O. Wilson and T.D. Siopes, 1979. Egg production, feed consumption and maintenance energy requirements of leghorn hens as influenced by dietary energy at temperatures of 15.6 and 26.7 C. Poult. Sci., 58: 674-680.
35. Tanor, M.A., S. Leeson and J.D. Summers, 1984. Effect of heat stress and diet composition on performance of white leghorn hens. Poult. Sci., 63: 304-310.
36. Alfredo, P. and C. Craig, 1991. Effect of temperature and dietary energy on layer performance. Poult. Sci., 70: 126-138.
37. Peguri, A. and C. Coon, 1993. Effect of feather coverage and temperature on layer performance. Poul. Sci., 72: 1318-1329.
38. Zollitsch, W., Z. Cao, A. Peguri, B. Zhang, T. Cheng and
C. Coon, 1996. Nutrient requirements of laying hens.
Proceedings of the International Symposium on Nutritional
Requirements of Poultry and Swine, (ISNRPS'96),
Departamento de Zootecnia, Vicosa-MG-Brasil, pp: 109-159.
39. Miller, P.C. and M.L. Sunde, 1975. The effects of precise constant and cyclic environments on shell quality and other lay performance factors with Leghorn pullets. Poult. Sci.,
54: 36-46.
40. Wolfenson, D., Y.F. Feri, N. Snapir and A. Berman, 1979. Effect of diurnal or nocturnal heat stress on egg formation. Br. Poult.
Sci., 20: 167-174.
41. Cufadar, Y., O. Olgun and A.O. Yildiz, 2011. The effect of dietary calcium concentration and particle size on performance, eggshell quality, bone mechanical properties and tibia mineral contents in moulted laying hens. Br. Poult.
Sci., 52: 761-768.
42. Skrivan, M., M. Marounek, I. Bubancova and M. Podsednicek,
2010. Influence of limestone particle size on performance and egg quality in laying hens aged 24-36 weeks and 56-68 weeks. Anim. Feed Sci. Technol., 158: 110-114.
43. Pelicia, K., E. Garcia, C. Mori, A.B.G. Faitarone and
A.P. Silva et al., 2009. Calcium levels and limestone particle size in the diet of commercial layers at the end of the first production cycle. Revista Brasileira de Ciencia Avicola,
11: 87-94.
44. Zhang, B. and C.N. Coon, 1992. Nutrition Institute on Minerals.
Chapter 7. Practical Applications. National Feed Ingredient
Association, Chicago, IL.
45. Zhang, B., J.V. Caldas and C.N. Coon, 2017. Effect of dietary calcium intake and limestone solubility on egg shell quality and bone parameters for aged laying hens. Int. J. Poult. Sci.,
16: 132-138.
46. Zhang, B., J. Weil, A.B. Guerra, P. Maharjan, K. Hilton,
N. Suesuttajit and C.N. Coon, 2019. The effect of feeding various limestone particle sizes, limestone solubility and calcium intake on bone status and shell quality of a commercial white layer strain from 18-65 weeks of age.
Int. J. Poult. Sci., 18: 372-378.
47. Cheng, T.K. and C.N. Coon, 1990. Sensitivity of various bone parameters of laying hens to different daily calcium intakes.
Poult. Sci., 69: 2209-2213.
48. Scott, M.L., S.J. Hull and P.A. Mullenhoff, 1971. The calcium requirements of laying hens and effects of dietary oyster shell upon egg shell quality. Poult. Sci., 50: 1055-1063.
49. Brister, Jr. R.D., S.S. Linton and C.R. Creger, 1981. Effects of dietary calcium sources and particle size on laying hen performance. Poult. Sci., 60: 2648-2654.
50. Keshavarz, K. and C.C. McCormick, 1991. Effect of sodium aluminosilicate, oyster shell and their combinations on acid-base balance and eggshell quality. Poult. Sci.,
70: 313-325.
51. El Hadi, H. and A.H. Sykes, 1982. Thermal panting and respiratory alkalosis in the laying hen. Br. Poult. Sci., 23: 49-57.
52. Barrett, N.W., K. Rowland, C.J. Schmidt, S.J. Lamont,
M.F. Rothschild, C.M. Ashwell and M.E. Persia 2019. Effects of acute and chronic heat stress on the performance, egg quality, body temperature and blood gas parameters of laying hens. Poult. Sci., 98: 6684-6692.
53. Cheng, T.K., C.N. Coon and M.L. Hamre, 1990. Effect of environmental stress on the ascorbic acid requirement of laying hens. Poult. Sci., 69: 774-780.
54. Odom, T.W., P.C. Harrison and W.G. Bottje, 1986. Effects of thermal-induced respiratory alkalosis on blood ionized calcium levels in the domestic hen. Poult. Sci., 65: 570-573.
55. Gordon, R.W. and D.A. Roland, Sr., 1997. The influence of environmental temperature on in vivo limestone solubilization, feed passage rate and gastrointestinal pH in laying hens. Poult. Sci., 76: 683-688.
56. Franco-Jimenez, D.J. and M.M. Beck, 2007. Physiological changes to transient exposure to heat stress observed in laying hens. Poult. Sci., 86: 538-544.
57. Franco-Jimenez, D.J., S.E. Scheideler, R.J. Kittok,
T.M. Brown-Brandl, L.R. Robeson, H. Taira and M.M. Beck,
2007. Differential effects of heat stress in three strains of laying hens. J. Applied Poult. Res., 16: 628-634.
58. Ebeid, T.A., T. Suzuki and T. Sugiyama, 2012. High ambient temperature influences eggshell quality and calbindin-D28k localization of eggshell gland and all intestinal segments of laying hens. Poult. Sci., 91: 2282-2287.
59. Manangi, M.K., P. Maharjan and C.N. Coon, 2018. Calcium particle size effects on plasma, excreta and urinary Ca and P changes in broiler breeder hens. Poult. Sci., 97: 2798-2806.
60. Rama Rao, S.V., M.V.L.N. Raju, S.S. Paul and B. Prakash,
2019. Effect of supplementing graded concentrations of non-phytate phosphorus on performance, egg quality and bone mineral variables in white leghorn layers. Br. Poult. Sci.,
60: 56-63.
61. Usayran, N., M.T. Farran, H.H. Awadallah, I.R. Al-Hawi,
R.J. Asmar and V.M. Ashkarian, 2001. Effects of added dietary fat and phosphorus on the performance and egg quality of laying hens subjected to a constant high environmental temperature. Poult. Sci., 80: 1695-1701.
62. Boling, S.D., M.W. Douglas, M.L. Johnson, X. Wang,
C.M. Parsons, K.W. Koelkebeck and R.A. Zimmermant, 2000.
The effects of dietary available phosphorus levels and phytase on performance of young and older laying hens.
Poult. Sci., 79: 224-230.

")






United States





