Introduction
L-carnitine (LC) is a naturally occurring and widely distributed in nature compound. It was discovered in 1905 in Liebig’s meat extracts, a popular ‘dietary supplement’ at that time, by Gulewitsch and Krimberg [1] who named the substance from the Latin word for flesh (“carnus”). Carnitine chemical structure was established in 1927 and its function was related to long-chain fatty oxidation in liver and heart in 1959 [2,3]. Therefore, investigations from the 1960s onwards have led to the uncovering of its biosynthetic pathway, transport mechanisms and deficiency syndromes [4]. Under certain stress conditions, the demand for LC may exceed an individual’s capacity to synthesize it, making it a conditionally essential nutrient. There is also evidence that both primary and secondary deficiencies of LC can occur and they are linked with inflammation, neuromuscular dysfunction, endocrine dysregulation, immune dysfunction, and abnormalities in energy metabolism [5]. Today, LC and its derivatives are widely used in genetic or primary disorders of LC metabolism and some other disorders resulted in secondary carnitine deficiency. They include end-stage renal diseases, cirrhosis, organic acidemias, Fanconi’s syndrome, long-term use of valproate and its associated hyperammonemic encephalopathy. LC is also effective in chronic fatigue syndrome, complications associated with antiretroviral therapy and impaired ketogenesis and fat metabolism in premature neo-nates, which have been linked to mitochondrial dysfunction [6].
During last decades there has been a growing interest in the potential uses of L-carnitine as a medicinal agent and as a nutritional/ dietary supplement. In addition to a great interest from medical sciences, carnitine received a substantial attention from poultry industry [7]. In a recent review it was concluded that in poultry production, LC has a multi-functional purpose, which includes: growth promotion, strengthening the immune system and antioxidant effects [8]. Indeed, feeding studies have shown that the performance of animals sharply drops when the LC content falls below 15-20 mg per kg of feed [7]. Because of the cereal grains are low in carnitine but represent the major component of poultry diets and recently animal feed ingredients have been excluded from the chicken diet in various countries, it could well be that chickens may face carnitine deficiency in various stress conditions.
The molecular mechanisms accounting for the positive effect of LC on many physiological parameters in farm animals and poultry are not yet fully understood, but many protective effects of LC reported in literature have been postulated to be related to its antioxidant action. However, by the time of writing no comprehensive review on this subject has appeared. Therefore, the aim of this paper is a critical analysis of recent data related to antioxidant action of carnitine in vitro and in vivo as a background for using LC for improvement of a strategy to fighting stresses in animal and poultry production.
Absorption and metabolism of carnitine
Carnitine homeostasis in human and animals is maintained by acquisition of carnitine from dietary sources, a modest rate of endogenous carnitine biosynthesis, and its efficient reabsorption [9]. The kinetics and pharmacokinetics of LC have been reviewed extensively elsewhere [9-10] and can be summarized as follows. Dietary LC is absorbed by active and passive transfer across enterocyte membranes with bioavailability of dietary LC to be 54-87%, depending on the amount of L-carnitine in the meal. Bioavailability of LC dietary supplements is lower, comprising about 14–18% of dose and unabsorbed LC is mostly degraded by microorganisms in the large intestine. Free
LC, absorbed from dietary intake or synthesized in liver and kidney, reaches the blood stream and the extracellular fluid and is then taken up by other tissues through active transport systems. L-Carnitine and its short-chain esters do not bind to plasma proteins. Excess carnitine is excreted via the kidneys [9]. L-Carnitine is eliminated from the body mainly via urinary excretion. For example, in the rat about 7% of the body pool is excreted in urine each day [11] and various physiological and pathophysiological factors as well as diet influence the rate of carnitine excretion and reabsorption. Indeed, the level of carnitine in blood is regulated mainly by the kidneys and is age and sex-dependent.
Dietary sources of LC are mainly meat products, while plant foods contain considerably lower carnitine levels. For example, carnitine content in ground beef is shown to be 582 mg/100g, while chicken, macaroni and corn flakes contain only 24, 0.13 and 0.013 mg carnitine per 100g [12]. Carnitine contents in grains (wheat, barley, corn, etc.) are about 5-7 mg/kg, while in soya meal and sunflower meal carnitine concentrations are 15 and 5 mg/kg respectively [7]. Since carnitine is present primarily in foods of animal origin, farm animal and poultry commercial diets formulated mainly from plant ingredients could be deficient in this element.
Carnitine biosynthesis involves a complex series of reactions in several tissues. In animals and man, LC is synthesized mainly in the liver from where it is transported to other tissues. It is interesting to note that about 98% of the carnitine body pool is localized in tissues that utilize fatty acids as their primary dietary fuel namely skeletal and heart muscle. It is well known, that synthesis of LC requires the essential amino acids lysine and methionine as well as such micronutrients as iron, ascorbic acid, vitamin B6 and niacin. Therefore, an incomplete diet, physiological stress situations and some clinical cases can create a need for external LC supplementation [13]. It should be taken into account that carnitine synthesis is a reasonably slow process and does not readily keep up with fast changes of the metabolic requirements in stress conditions [14]. In fact, about 75% of carnitine is coming from the diet, while only 25% from internal de novo synthesis [11]. The human body contains about 20–25g of LC and an additional 100–300 mg per day can be taken in through our diet [15].
In mammalian cells and body fluids, carnitine is present either as free carnitine, short-chain acyl carnitine, or long-chain acylcarnitine. In particular, acylcarnitines are shown to account for about 20% of total carnitine in serum and 10-15% in muscle and liver [16]. For example, acyl and free carnitine serum concentrations are, respectively, 12.8 ± 7.4 and 67 ± 21.8 µmol/L [17] and circulating carnitine accounts for only about 0.5% of body carnitine [18]. Plasma carnitine concentration is comparable with those of other antioxidants, including vitamin E (20-30 µM) and ascorbic acid (26.1–84.6 µM [19]). Mammalian tissues contain relatively high amounts of LC, ranging between low µM to low mM, with the highest concentrations in heart and skeletal muscles. Indeed, the carnitine concentrations are ≈1 mM in rat skeletal muscle, ≈3 mM in human muscle and may be up to 15 mM in ruminant muscle [20]. Concentrations of LC and acetyl-carnitine (ALC) change under altered dietary conditions. ALC is the most extensively investigated derivative of the carnitine formulations, largely due to its pharmacokinetic advantages such as reliable absorption and efficient transport [21]. The uptake of LC into most tissues against a concentration gradient involves carrier-mediated transport systems, which maintain high tissue/plasma concentration ratios [22]. Delivery of carnitine into cells, distribution of carnitine within the body and intracellular homeostasis of carnitine are controlled by novel organic cation transporters (OCTN; [23]). In particular, active sodium-dependent high affinity OCTN2 transporter is responsible for LC transport in the kidney and other tissues [24]. Mean turnover time for carnitine for such tissues as liver and kidney is 12h, while for slow-turnover tissues (e.g. skeletal muscle) it is 191h. It is interesting to note that for extracellular fluids carnitine turn-over time is only 1. 1h and mean whole-body turnover time is 66h, ranging from 38 to 119h [9].
As LC is not regarded as an essential nutrient, no values for dietary reference intake or recommended daily allowance have been set. Main carnitine function in the body include [25-26] :
- Transport of activated long-chain fatty acids from the cytosol to the mitochondrial matrix, where β-oxidation takes place;
- Transfer of the products of peroxisomal β-oxidation, including acetyl-CoA, to the mitochondria for oxidation to CO2 and H2O in the Krebs cycle;
- Modulation of the acyl-CoA/CoA ratio;
- Storage of energy as acetyl-carnitine;
- Modulation of toxic effects of poorly metabolized acyl groups by excreting them as carnitine esters.
- Preservation of membrane integrity and mitochondria functions and apoptosis inhibition
Antioxidant systems of the cell and whole body
Antioxidant systems of the living cell are based on three major levels of defense [27-29] and include several options [29] :
- Decrease localized oxygen concentration;
- Decrease activity of pro-oxidant enzymes and improve efficiency of electron chain in the mitochondria and decreasing electron leakage leading to superoxide production;
- Prevention of chain initiation by scavenging initial radicals due to induction of various transcription factors (e.g., Nrf2, NF-κB and others) and ARE-related synthesis of AO enzymes (SOD, GSH-Px, CAT,GR, GST, etc.);
- Vitagene activation, synthesis and increased expression of protective molecules (GSH, thioredoxins, heat shock proteins, sirtuins, etc.);
- Binding metal ions (metal-binding proteins) and metal chelating;
- Decomposition of peroxides by converting them to non-radical, nontoxic products (Se-GSH-Px);
- Chain breaking by scavenging intermediate radicals such as peroxyl and alkoxyl radicals (vitamins E, C, GSH, uric acid, ubiquinol, bilirubin, etc.)
- Repair and removal of damaged molecules (methionine sulfoxide reductase, DNA-repair enzymes, chaperons, etc.).
Antioxidant action of carnitine
The cytoprotective effects of carnitine are believed to be associated with a decrease in oxidative stress. It should be noted that carnitine can contribute to the antioxidant defenses in different ways. Firstly, by direct free radical scavenging. Secondly, by preventing free radical formation by inhibiting specific enzymes responsible for free radical production or by maintaining the integrity of electron-transport chain of mitochondria in stress conditions. Thirdly, by participating in the maintenance of optimal redox status of the cell by activating a range of antioxidant enzymes and non-enzymatic antioxidants, mainly via transcription factors, including Nrf2 and NF-κB.
Finally, by activating an array of vitagenes, responsible for the synthesis of protective molecules, including HSP, thioredoxin (Trx), sirtuins, etc., and providing additional protection in stress conditions.
Protective effects of carnitine on mitochondria
Mitochondria are the primary cellular consumers of oxygen and contain numerous redox enzymes capable of transferring single electrons to oxygen, generating the ROS superoxide (O2–). It is well appreciated that mitochondrial enzymes known to generate ROS include the tricarboxylic acid cycle enzymes aconitase and α-ketoglutarate dehydrogenase; the electron-transport chain (ETC) complexes I, II and III; pyruvate dehydrogenase and glycerol-3-phosphate dehydrogenase; dihydroorotate dehydrogenase; the monoamine oxidases A and B; and cytochrome b5 reductase [40]. Furthermore, mitochondrial insults, including oxidative damage itself, can cause an imbalance between ROS production and removal, resulting in net ROS production. For example, ROS, which is an inevitable by-product of oxidative phosphorylation, induce protein modifications, lipid peroxidation and mitochondrial DNA damage, which ultimately results in mitochondrial dysfunction [41]. Many studies have focused on the detrimental effects of ROS, but it is now clear that mitochondrially generated ROS are also involved in regulating intracellular signal transduction pathways that result in cell adaptation to stress [42].
One of the mechanisms responsible for the decrease in oxidative stress is the protective effect of carnitine on mitochondrial structure and function. Indeed LC protects mitochondria from pathological events by triggering pro-survival cell signaling. For example, LC (300 mg/kg b.w., i.p.) improved the electron transport chain complexes levels in heart and skeletal muscles of aged rats when compared with young rats in duration dependent manner [43]. There is a range of publications showing protective effect of LC on mitochondria invitro. For example, ALC provides mitochondrial support and conserves growth factor receptors [44]. In fact, at micromolar concentrations (50 µM), LC reduced the three markers of oxidative stress (MDA, ROS formation and mitochondrial dysfunction) in brain synaptosomal fractions treated with a combination of quinolinic acid and 3-nitropropionic acid [45]. Furthermore, pre-treatment with LC (10-100 µM) reduced oxidant formation and increased mitochondrial membrane potential in insulinoma cells and isolated rat islet cells chronically exposed to oleic acid [46]. ALC (1 mM) effectively suppressed the oxidative stress in and around mitochondria of thioredoxin 2-deficient DT40 cells thereby preventing mitochondrial signaling pathway leading to apoptosis [47]. LC (10-100 µM) could protect human proximal tubule epithelial cells from H2O2-induced injury through the inhibition of oxidative damage, mitochondria dysfunction and inhibition of cell apoptosis [48]. Indeed, pre-treatment with LC for 12h inhibited H2O2-induced cell viability loss, intracellular ROS generation and lipid peroxidation in a concentration-dependent manner. Furthermore, mitochondrial dysfunction associated with cell apoptosis including membrane potential loss, down-regulation of Bcl-2 and upregulation of Bax and the release of cytochrome c were abrogated in the presence of LC. It was demonstrated that 4-week pre-treatment with ALC (100 µM) effectively protected human neuroblastoma cells against rotenone-induced mitochondrial dysfunction and oxidative damage [49].
In HepG2 cells LC (1 mM) prevented free fatty acid-induced apoptosis (16% vs. 3%, P < 0.05) and damages to mitochondria and their dysfunction by increasing mitochondrial β-oxidation and reducing intracellular oxidative stress [50]. Pre-treatment with ALC (0.1 mM) significantly inhibited TGF-β1-induced mitochondrial ROS production in bone marrow derived mesenchymal stem cells [51]. LC (50, 100 and 200 µM) protects retinal ganglion cells from high glucose-induced injury through the inhibition of oxidative damage, mitochondrial dysfunction and, ultimately, cell apoptosis [52]. It should be mentioned that all antioxidants in the body and cells are working together in a cooperative manner to build the effective antioxidant defense network. In such a system, effective concentrations of individual antioxidants can be substantially reduced. For example, it was found that when combined, LA and ALC (0.1 µM + 0.1 µM or 1 µM + 1 µM) worked at 100-1000-fold lower concentrations than they did individually [53].
There is also in vivo evidence of stabilizing effect of carnitine on mitochondria function and decreasing ROS production. For example, the activities of citric acid cycle enzymes, such as isocitrate dehydrogenase, alpha-ketoglutarate dehydrogenase, succinate dehydrogenase, and malate dehydrogenase, as well as that of electron-transferring enzymes such as NADH dehydrogenase and cytochrome c oxidase were found to be decreased in aged rats. However, after supplementation of carnitine to aged rats, the activities of these enzymes showed nearer to that of young control rats [54]. Furthermore, LC supplementation (5g/kg diet) significantly augmented the oxidative capacity of both liver and muscle by enhancing the activity of carnitine palmitoyl transferase and the respiratory chain enzymes in rat mitochondria after dietary lipid manipulation and physical activity [55]. Exogenous LC (300 mg/kg b.w.) improved function of respiratory chain and antioxidant capacity (increased SOD activity and decreased MDA) in mitochondria of myocardium after exhaustive running in rats [56]. ALC activates the peroxisome proliferator-activated receptor-γ coactivators PGC-1α/PGC-1β-dependent signaling cascade of mitochondrial biogenesis, partially reverses the mitochondrial SOD2 decline and decreases the oxidized peroxiredoxins content in old rat liver [57]. Indeed, the antioxidant mechanism of LC and ALC supplementation appears to be associated with reductions in mitochondrial ROS production [58]. In fact, in TNF-α stimulated human intestinal microvascular endothelial cells PLC acted at the mitochondrial level by reducing superoxide generation and inhibiting the downstream-regulated IL-8 and MCP-1 cytokine secretion [59].
Therefore, as can be seen from the aforementioned analysis of the recent data, carnitine can be considered as a mitochondria-specific antioxidant, responsible for mitochondria integrity maintenance and regulation of ROS production and ROS signaling.
Inhibition of free-radical generating enzymes by carnitine
Xanthine oxidase
Xanthine oxidase (XO) catalyzes the terminal two steps of purine degradation, converting hypoxanthine to uric acid and considered to be a critical source of both O2- and H2O2 in inflammatory diseases [60]. In particular, XO is considered to be the main source of oxidative stress and destructive free radicals in various clinical conditions including heart attacks and stroke [61]. The in vitro effect of ALC on spontaneous and induced lipid peroxidation in rat skeletal muscle was studied. A significant reduction of XO activity was detected in the presence of 10-80 mM ALC [31]. However, the concentration of ALC used in the aforementioned study is not achievable in vivo. In several other publications, carnitine is shown to decrease XO activity in vivo. For example, exercise-induced increases in human plasma XO was significantly attenuated by L-carnitine L-tartrate (LCLT; [62]. In the study, the lower XO response during LCLT was also associated with less accumulation of ROS (as measured by MDA). There was also indirect evidence that ALC inhibits XO activity in
MPTP-treated rats [63]. Indeed, ALC pre-treatment caused a significant decrease in the uric acid produced, likely due to its inhibition of XO. A decreased XO activity in serum of post-exercised patients due to LC (1 or 2g) supplementation was also shown [64]. Indeed, LC significantly prevented increase in XO activity in serum of men and women during acute resistance exercise challenge [65]. In fact, two grams of LC supplementation significantly attenuated XO and MDA in serum of the patients.
NADPH oxidase
ROS generation by NADPH oxidase enzyme complex plays a critical role in several physiological processes including host defense, post-translational modification of proteins, cell differentiation and regulation of gene expression [66]. Indeed, NADPH oxidase, the major enzyme involved in the production of superoxide anion [67]. Incubation of platelets with LC significantly reduced arachidonic acid-mediated NADPH oxidase activation [68]. In cardiac fibroblasts in vitro, LC (1-30 mM) attenuated angiotensin II-induced NADPH oxidase activity, ROS formation, extracellular signal-regulated kinase phosphorylation, activator protein-1-mediated reporter activity and sphingosine-1-phosphate generation [69]. In primary cultured neonatal rat cardiomyocytes LC (1-30 mM) inhibited doxorubicin-induced ROS generation and NADPH oxidase activation, reduced the quantity of cleaved caspase-3 and cytosol cytochrome c, and increased Bcl-x(L) expression [70]. In cultured TNF-α-stimulated human intestinal microvascular endothelial cells, propionyl-L-carnitine (PLC) counteracted NADPH oxidase 4-generated oxidative stress-induced cell adhesion molecules expression and leukocyte adhesion [71]. In human umbilical vein endothelial cells in vitro, PLC pre-treatment restored serum-deprived and TNF-α-induced impaired mitochondrial β-oxidation by reducing flavin adenine dinucleotide level and counteracting increased CAM and Nox4 expression, leukocyte adhesion and inflammatory cytokine secretion [59]. PLC (1 mM) increased human umbilical vascular endothelial cell proliferation and rapidly reduced inducible nitric oxide synthase and NADPH-oxidase 4-mediated ROS production in human umbilical vascular endothelial cells [72]. Indeed, the authors clearly showed that the PLC antioxidative effect is mainly NADPH oxidase 4-mediated. Furthermore, there was PLC-induced NF-κB–related gene expression reduction [72]. The left ventricles of hypertensive rats showed a higher expression of p22phox subunit of NADPH oxidase (2.1-fold), an alteration that was corrected after treatment with LC and captopril [73]. ALC treatment almost completely neutralized the effects of alcohol on NADPH oxidase1 protein expression in the rat brain [74]. It was shown that LC is helpful in modulating oxidative stress and platelet activation during major abdominal surgery-dependent oxidative damage by decreasing soluble NOX2-derived peptide (sNOX2-dp), a marker of NADPH oxidase activation [75]. It was also shown that L-NAME-treated rats suffered a 44 % increase in NADPH oxidase activity compared with normotensive animals. LC simultaneous treatment managed to reduce the activity of this enzyme up to basal levels [76]. Furthermore, a remarkable increase of NOX4 protein expression in rats treated with L-NAME was observed, an alteration that was also prevented by simultaneous treatment for these animals with LC. In another study from the same department an elevation in NADPH oxidase subunits (NOX2 and NOX4), in hypertensive rats, when compared with normotensive ones, was observed. Those changes were accompanied by a down-regulation of PPAR-γ in the heart of hypertensive animals. When hypertensive rats were treated with LC, all these alterations were reversed [77]. Similarly, an increase in the expression of Nox2, and Nox4 was found in the kidney of L-NAME-treated rats. Simultaneous treatment with LC attenuated the renal fibrosis and the pro-oxidative and pro-inflammatory status reported in L-NAME groups, with a concomitant increase in the expression of PPAR-γ [78]. Therefore, it is clear that LC and its derivatives can decrease activities of ROS-generating enzymes and in this way contributing to improved antioxidant defenses.
In vitro antioxidant effects of carnitine
Important evidence of protective effect of carnitine against oxidative stress caused by various chemicals came from in vitro studies with cell culture or isolated cells or organelles. For example, treatment of human dermal fibroblasts with 1-5 mM carnitine significantly reduced the inhibitory effects of cGMP on catalase and other peroxisomal enzyme activities [79]. LC (1-10 mM) protects against glutamate- and kainic acid-induced neurotoxicity in cerebellar granular cell culture of rats [80]. L-carnitine (100 µM) protected the lipid as well as protein part of LDL particles against oxidative modifications. Indeed, incorporation of LC into LDL, incubated with Cu2+ ions, caused a decrease in the level of conjugated dienes, lipid hydroperoxide, MDA, and dityrosine. Furthermore, LC caused a significant two-fold increase in α-tocopherol content in oxidized LDL [81]. LC (0.1-1 mM) protected human hepatocyte cells against cytotoxity induced by H2O2 by regulation PPAR-α, scavenging ROS, promotion of SOD and CAT activity and expression, and prevention of lipid peroxidation [82]. LC (9, 12 and 25 µM) decreased lipid peroxidation in the cultured porcine oocytes treated with H2O2 [83]. It was shown that LC pre-treatment (100 µM) could increase neuroblastoma cell viability; inhibit apoptosis and ROS accumulation caused by H2O2 or tunicamycin [84]. Administration of LC (30-100 µM) to neurons harvested from the forebrain of new-born rats significantly diminished ROS generation and provided near complete protection of neurons from ketamine-induced cell death [85]. It was shown that Leucine and α-ketoisocaproic acid caused increased DNA damage and LC (120 or 150 µM) was able to significantly prevent this damage [86]. There is also experimental evidence that LC (30-150 µM) can reduce the in vitro DNA injury induced by high concentrations of phenylalanine [87] or by propofol [88]. Indeed, the aforementioned data confirmed antioxidant action of carnitine in physiologically-relevant concentrations in various in vitro systems.
Antioxidant effects of carnitine against oxidative stress in vivo: Protection against toxicants
CCl4
CCl4 is considered as an important hepatotoxin due to its severe oxidative effect on this organ. Indeed, metabolism of CCl4 via CYP2E1 to highly reactive ROS plays a major role in the mode of action of the toxicant [89]. It is well established that CCl4 inhibits AO enzymes (SOD, GSH-Px and CAT) and GSH in the liver samples [90], increases the secretion of ALT, AST, ALP, γ-GT due to hepatic injuries caused by ROS [91] and enhances TBARS in the liver [92] and serum [93]. It has been shown that practically all elevated indexes of the oxidative stress caused by CCl4 were restored almost to the initial physiological levels by carnitine treatment. For example, ALC (200 mg/kg b.w. for 4 days) prevents CCl4-induced oxidative stress in various tissues of rats including restoration of vitamin C, vitamin E, GSH levels and SOD, GSH-Px and CAT activities in hemolysate, liver, kidney and brain tissues [94]. Similarly, LC (100 mg/kg, i.p.) prevented increase in MDA and myeloperoxidase levels in the liver tissue samples of rats treated with CCl4 [95]. It is interesting to note, that in CCl4-induced liver fibrosis, the protective efficacy against hepatic oxidative stress was in the following order: melatonin > L-carnitine > vitamin E, while in STZ-induced diabetes, the efficacy order was vitamin E > or = melatonin > L-carnitine [96].
Cisplatin
Cisplatin (CDDP) is a chemotherapeutic drug widely used against a variety of cancers and its nephrotoxicity is mainly due to ROS production and oxidative stress [97]. It was shown that CDDP caused decreased activities of AO enzymes (SOD and GSH-Px) and GSH and increased MDA in rat liver [98-99] and significantly elevated serum activities of LDH and creatine kinase [99]. PLC (500 mg/kg b.w.) improved antioxidant defenses (increased GSH and decreased TBARS) in rat cardiac tissues and prevented the progression of cisplatin-induced cardiomyopathy in a carnitine-depleted rat model [100]. Furthermore, ALC (500 mg/kg b.w., i.p) significantly attenuated the cisplatin-evoked disturbances in antioxidant defenses (SOD and GSH) in cardiac tissues of rats [99]. It is interesting to note that protective mechanisms of ALC against cisplatin-induced apoptosis are related to activation of anti-apoptotic Bcl family members’ genes, and in an Akt-related gene expression dependent manner [101]. In addition, in cultured human tubular cells, cisplatin reduced SIRT3, resulting in mitochondrial fragmentation, while restoration of SIRT3 with ALC (200 or 500 mg/kg b.w., i.p.) improved cisplatin-induced mitochondrial dysfunction [102].
Ethanol
Oxidative stress plays an important role in the pathogenesis of alcoholic liver damage. During ethanol metabolism, ROS and reactive nitrogen species (RNS) are forms causing oxidative stress [103]. Indeed, ALC (50 μM) stabilized SOD activity in primary human brain endothelial cells during alcohol-induced oxidative stress [104]. In fact, long-term ethanol exposure reduces SOD activity in cells causing mitochondrial injury and ALC prevented the changes. It has been shown that a single oral dose of 200 mg/kg body weight of PLC 1h before alcohol intake prevented alcohol-induced increase in TBARS and increased the gastric content of GSH, besides it increased the enzymatic activities of gastric SOD and GST in rats [105]. Similarly, LC (500 mg/kg, intragastrically) prevented an increase in plasma and gastric lipid peroxidation and stabilized gastric GSH levels in ethanol-intoxicated rats [106]. In fact, LC administration (1.5 g/L of drinking water for a week) to ethanol-intoxicated rats significantly protects phospholipids and proteins against oxidative modifications [107]. The administration of LC (1.5 g/L of drinking water) to ethanol-intoxicated rats partially normalized the activity of the antioxidant enzymes (Cu, Zn-SOD, GSH-Px, GR and CAT) and the level of the non-enzymatic antioxidants (vitamin C, E, A and GSH [108]).
Moreover, LC significantly protects lipids and proteins against oxidative modifications.
In the aforementioned studies, the free radical scavenging and antioxidant properties of LC and its derivatives are demonstrated by:
(a) restoration of the endogenous AO enzymes (SOD, CAT, GSH-Px, GR and GST) and non-enzymatic antioxidants (vitamins E and C) in the liver and other tissues of stressed animals; (b) increased intracellular concentration of GSH in liver and other tissues; (c) decreased lipid and protein oxidation, detected as reduced MDA/TBARS and carbonyl content; (d) decreased DNA fragmentation/damage and apoptosis; (e) reduced secretion of ALT, AST, ALP, γ-GT from the liver into the plasma due to hepatic injuries caused by ROS. Indeed, antioxidant action of carnitine was significant and important in prevention of negative consequences of toxicities caused by various agents.
Effect of carnitine on vitagene network and transcription factors activation
The term “vitagene” was introduced in 1998 by Rattan [109] and later vitagene concept has been further developed by Calabrese and colleagues [21, 110-119]. In accordance with Calabrese., et al. [111, 116] the term vitagenes refers to a group of genes that are strictly involved in preserving cellular homeostasis during stress conditions and the vitagene family includes heat shock proteins (HSPs), heme oxygenase-1 (HSP32, HO-1), HSP60 and HSP70, the thioredoxins (Trx)/thioredoxins reductase (TrxR) system and sirtuins. The list of potential candidates to the vitagene family can be extended. In particular, it seems likely that SOD, a major inducible enzyme of the first level of antioxidant defense, can meet selecting criteria to be included into the vitagene family [29]. The products of the mentioned genes actively operate in detecting and controlling diverse forms of stress and cell injuries. The cooperative mechanisms of the vitagene network are reviewed in recently published comprehensive reviews [116,119-120] with a major conclusion indicating an essential regulatory role of the vitagene network in cell and whole organism adaptation to various stresses.
As can be seen from the data presented above, main protective effects of LC and ALC were associated with preservation or increased activity of antioxidant enzymes and GSH in various stress conditions. The mechanisms involved in the regulation of antioxidant enzymes by LC in vivo have not been precisely determined yet. However, it seems likely that transcription factors, including Nrf2, are involved in this regulation. First, it was shown in vitro that treatment of astrocytes with acetyl-L-carnitine (30-100 μM) induces vitagene HO-1 in a dose- and time-dependent manner and that this effect was associated with up-regulation of another vitagene HSP60 as well as high expression of the redox-sensitive transcription factor Nrf2 in the nuclear fraction of treated cells [121]. Furthermore, treatment for 4 months of senescent rats with ALC (150 mg/kg/day, orally) induces vitagene HO-1 as well as other vitagenes namely Hsp70 and SOD-2. This effect was associated with up-regulation of GSH levels, prevention of age-related changes in mitochondrial respiratory chain complex expression, and decrease in protein carbonyls and HNE formation [122]. In human endothelial cells in culture carnitine and its acyl derivatives (at 0.5-2 mM) were shown to increase gene and protein expression of HO-1 [123]. Furthermore, in humans and in an animal model it was shown that carnitine-mediated improved response to erythropoietin involves induction of HO-1 [124].
Pre-treatment of primary cortical neuronal cultures with ALC (100 μM) significantly attenuated amyloid-beta peptide 1-42-induced cytotoxicity, protein oxidation, lipid peroxidation, and apoptosis in a dose-dependent manner by up regulation of HSPs [125]. The authors showed that ALC exerted protective effects against oxidative stress in part by up-regulating the levels of GSH and HSPs. Indeed, LC treatment was associated with an increased level of HO-1 in the retinal ganglion cells [126]. LC (50 μM, 100 μM and 200 μM) has protective effects on high glucose-induced oxidative stress in the retinal ganglion cells (RGCs). Indeed, in high glucose stimulated RGCs, LC treatment was associated with an increased level of Nrf2, HO-1 and γ-glutamyl cysteine synthetase [126]. Furthermore, ALC administration to human lens epithelial cells treated with homocysteine, restored (increased) the levels of antioxidant proteins, including SOD, GSH-Px, CAT, Nrf2, Keap1 and GSH [127].
Both oxidative stress and mitochondrial damage are associated with reduced levels of renal sirtuin 3 (SIRT3). Treatment with ALC restored SIRT3 expression and activity, improved renal function, and decreased tubular injury in animals [102]. Thus, ALC and sirtuins together affect mitochondria acetylation/de-acetylation and thereby have the potential to regulate the cellular redox state and energy homeostasis [128]. From the aforementioned data it is clear that carnitine can be considered as an important regulator of the vitagene network.
Recently, it has been confirmed that the molecular regulation of antioxidant enzymes through an inhibition of the renin-angiotensin system and a modulation of the NF-κB/IκB system seems to be responsible for the antioxidant effect of carnitine [129]. Indeed, the chronic administration of LC was able to produce a transcriptional up regulation of GSH-Px and SOD enzymes, leading to an improvement of the cellular antioxidant defenses. It has also been reported that LC can activate another transcription factor, namely the peroxisome proliferator activated receptor alpha (PPARα). It has been proved that PPARα plays an important role in LC anti-apoptotic effect in renal tubular cells [130]. In fact, it was found that in NRK-52E cells LC increased PPARα activity more than 5-fold [130]. These results reveal the crucial role of PPARα activation in the LC protective function on gentamicin-induced apoptosis in NRK-52E cells. A decrease in the expression of transcription factors Nrf2 and PPARα, together with an increase in NF-kB expression, was observed in the renal cortex of L-NAME-induced hypertensive rats compared with control rats (0.3-, 0.8-, and 13-fold, respectively). The simultaneous administration of LC attenuated these alterations, reaching values similar to those found in control rats [76-78]. The authors suggested that the beneficial effect of LC supplementation was associated with upregulation of both antioxidant enzymes and eNOS, and with a down regulation of both NADPH oxidase and RAS components.
Carnitine-vitamin E interactions
It is well known that vitamin E is main chain-breaking antioxidant in biological membranes which cannot be replaced by other antioxidants [27,131]. In fact, vitamin E recycling is considered to be the most important part of vitamin E efficacy in antioxidant defenses. Indeed, when all necessary elements of vitamin E recycling are present together with other antioxidant mechanisms, even low vitamin E level in membranes, for example, in the brain, can effectively protect the tissue against lipid peroxidation [27,132]. As a part of the antioxidant systems carnitine can have a sparing effect on vitamin E. For example, dietary LC (150 mg/kg diet) enhanced the rates and amounts of lymphatic absorption of α-tocopherol and fat in ovariectomized rats [133] and increased liver α-tocopherol and lowers liver and plasma triglycerides in aging ovariectomized rats [134]. Similarly, carnitine supplementation lowers lipid peroxidation and promotes conservation of retinol and α-tocopherol in free-living women [135]. Furthermore, administration of LC (1.5g/L with drinking water) to rats intoxicated with ethanol significantly protects lipids and proteins against oxidative modifications in the serum and liver and the level of vitamin E was increased by about 20% in the liver and blood serum in comparison to the ethanol group [136]. In the irradiated rats treated with LC (1.5 mg/kg b.w, i.p.) concentrations of vitamins A, C and E were higher than in those rats that were only exposed to 2.45-GHz radiation [137]. On the other hand, metabolomics analysis shows that α-tocopherol deficiency in rats was associated with an increase in carnitine content in the liver [138]. Molecular mechanisms of carnitine-vitamin E interactions are not known yet, but the effect of such interactions on the total antioxidant systems of the body could be quite significant.
Carnitine supplementation as a part of antioxidant mixtures
Based on the concept of integrated antioxidant systems in the body one can expect that dietary supplementation of synergistic mixtures of various antioxidants could have higher protective effects in comparison with individual antioxidants, including carnitine.
For example, LC and vitamin E in combination have the ability to ameliorate ochratoxin A-altered haematological and serum biochemical parameters in White Leghorn cockerels [139]. It was shown that the risk of ischemia-induced necrosis in flap attempts made in damaged tissues might be reduced by the combination of L-carnitine and vitamin C [140]. Similarly, supplementation of vitamin E, vitamin C, and L-carnitine in combination was shown to be beneficial in attenuating the oxidative stress associated with intermittent hypobaric hypoxia in rats [141]. This idea of combining antioxidant usage was also proven in a number of studies when carnitine was used simultaneously with another mitochondria-related antioxidant, namely lipoic acid [43,54,142-149]. Furthermore, a more complex antioxidant mixture, containing CoQ, LC, α-tocopherol and selenium was shown to decrease DNA damage in the liver of fumonisin
B1-treated rats [150]. A combination of ALC, folate and vitamin E provided a synergistic protection against oxidative stress resulting from exposure of human neuroblastoma cells to amyloid-beta [151]. Indeed, vitamin E prevents de novo membrane oxidative damage, folate maintains levels of the endogenous antioxidant GSH and ALC prevents A-beta-induced mitochondrial damage and ATP depletion providing superior protection to that derived from each agent alone [151]. Similarly, supplementation of pregnant and lactating sow diet with carnitine-containing bioactive substances (a blend of flax seed, rapeseed, linden inflorescence, taurine, LC and tocopherol acetate) enhanced maturation of the small intestinal epithelium in their offspring during the early postnatal period [152]. Recently, it was shown that ALC, L-α-lipoic acid and silymarin had similar antioxidant effects in cisplatin-induced myocardial injury [99]. It would be worthwhile to study an antioxidant effect of a combination of carnitine and silymarin taking into account that both are considered to be hepato-protectors and both are characterized by antioxidant properties [29]. Indeed, the therapeutic effect of silymarin combined with LC on non-alcoholic fatty liver disease in patients was higher that silymarin alone [153].
Future directions in the development of carnitine-based antioxidant composition
Taking into account the aforementioned data and results of our recent research [28, 29, 39, 131, 154-155] it is clear that in order to deal with commercially-relevant stresses in poultry and pig production it is necessary to develop a product meeting at least three important requirements:
- Vitagene activation and redox-signaling (carnitine, betaine, vitamins A, E, D, C, Se, Zn, Mn, silymarin and other phytochemicals);
- Vitamin E and system of its recycling (vitamin C, Se, Vitamins B1 and B2);
- Carnitine synthesis (lysine and methionine, ascorbic acid, vitamin B6 and niacin).
Indeed, it would be difficult to accommodate all the aforementioned requirements in a single product and this could be addressed in a few different products. Inclusion of various protective compounds into the diet of farm animals and poultry to decrease negative consequences of stress conditions is complicated, firstly, by a decreased feed consumption at time of stress. Secondly, such an approach has a low flexibility, since existing feeding systems do not allow to include anything into the feed loaded into the feed storage bins located near the poultry/pig house (usually several tons of feed for several days feeding). Therefore, before the previous feed is consumed, nothing can be added to the feed. However, sometimes it is necessary to supplement animals/poultry with specific additives very quickly to deal with consequences of unexpected stresses (e.g. mycotoxins in the feed, immunosuppression, high temperature, etc.). In such a case additive supplementation via drinking system is a valuable option. In fact, modern commercial poultry and pig houses have water medication equipment installed, which can be perfectly used for the aforementioned supplementations. For example, an attempt to address the aforementioned option was implemented in a commercial product PerforMax, containing a synergistic mixture of 28 compounds, including carnitine, vitamins, minerals, betaine and amino acids and supplied via drinking water. Its efficacy in fighting stresses in commercial poultry production has been recently reviewed [155] and prospects of its use to maintain gut health in weaned piglets and newly hatched chicks were considered [39]. Indeed, it is well known that commercial animal/poultry production is associated with a range of stress conditions including environmental (high temperature), nutritional (mycotoxins and oxidized fat) or internal (vaccinations, disease challenges, etc.) stresses. In such conditions, supplying the PerforMax with drinking water was shown to have protective effects in growing birds [157-158] as well as in adult birds [155] helping maintain their health, productive and reproductive performance. Therefore, the aforementioned results are the first step to go from the development of the vitagene concept to design of the commercial product and testing it in the commercial conditions of poultry and pig production. We can suggest that this idea could be realized in human nutrition as well. Clearly, more research is needed to understand a fundamental role of carnitine and vitagenes in adaptation to various stresses. Indeed, it is just a matter of time before commercial products based on this idea found their way to the shelves of healthy nutrition shops and veterinary clinics.
Conclusions
Carnitine is an important element regulating many various functions in the animal/human body, including transport of activated long-chain fatty acids from the cytosol to the mitochondrial matrix, where β-oxidation takes place; transfer of the products of peroxisomal β-oxidation, including acetyl-CoA, to the mitochondria for oxidation to CO2 and H2O in the Krebs cycle; modulation of the acyl-CoA/CoA ratio; storage of energy as acetyl-carnitine; modulation of toxic effects of poorly metabolized acyl groups by excreting them as carnitine esters [25]. It is also known, that about 75% carnitine is coming from the diet and about 25% is synthesized in the body from amino acids methionine and lysine. Furthermore, it is well established that carnitine synthesis depends on many various conditions and nutritional status of farm animals/birds and the amount of carnitine obtained from endogenous synthesis and feedstuffs may be insufficient in some situations such as stress and high performance. Furthermore, because of the cereal grains and their by-products are low in carnitine but represent the major component of poultry and pig diets and recently animal feed ingredients have been excluded from the chicken/animal diets in various countries, probability of carnitine insufficiency in poultry and pig nutrition increased.
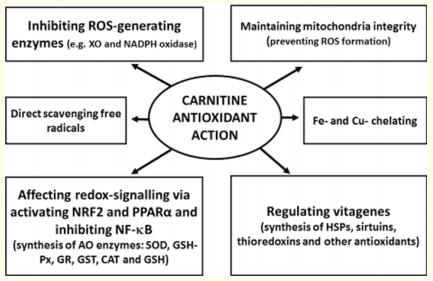
Therefore, it seems likely that carnitine dietary supplementation will receive more attention in future. The molecular mechanisms accounting for the positive effect of LC on many physiological parameters in farm animals and poultry are not yet fully understood, but many protective effects of LC reported in literature have been postulated to be related to its antioxidant action. Based on the analysis of the recent publications it could be concluded that antioxidant action of carnitine is related to (Figure 1):
- Its direct scavenging free radicals;
- Chelating catalytic metals-promoters of ROS, such as Fe and Cu;
- Maintaining mitochondria integrity in stress conditions and preventing ROS formation;
- Inhibiting ROS-generating enzymes, such as XO and NADPH-oxidases;
- Affecting redox-signaling via activation of Nrf2 and PPARα and inhibition of NF-κB with additional synthesis of antioxidant enzymes (SOD, GSH-Px, GR, GST, CAT, etc.) and small antioxidant molecules (GSH);
- Regulating vita genes and synthesis of HSPs, sirtuins, thioredoxins and other antioxidant molecules.
Figure 1: Carnitine antioxidant action.
It could well be that in biological systems in vivo all the aforementioned mechanisms are interacting and providing an important place for carnitine to be a crucial part of the integrated antioxidant systems of the animal and human body. It seems likely, that carnitine participation in redox signaling and affecting transcription factors (Nrf2, PPARα, NF-κB, etc.) as well as activating the vitagene network are main mechanisms responsible for antioxidant action of LC and its derivatives. Furthermore, protective roles of carnitine in stress conditions of commercial poultry and pig production are difficult to overestimate. Therefore, a development of carnitine-containing antioxidant compositions supplying via drinking water seems an important way forward in decreasing detrimental consequence of various stresses in poultry and pig production.
Conflict of Interest
There is no conflict of interest.
This article was originally published in EC Veterinary Science 2.1 (2015): 66-84.
Bibliography
- Gulewitsch W., et al. “ZurKenntnis der Extraktivstoffe der Muskeln”. Hoppe-Seyler’s Zeitschrift für physiologische Chemie 45 (1905): 326-330.
- Karlic H., et al. “Supplementation of L-carnitine in athletes: does it make sense?” Nutrition 20.7-8 (2004): 709-715.
- Zurbriggen E. “L-carnitine: Historical review”. Annals of Nutrition & Metabolism 44 (2000): 78-79.
- Wolf G. “The discovery of a vitamin role for carnitine: The first 50 years”. Journal of Nutrition 136.8 (2006): 2131-2134.
- Crentsil V. “Mechanistic contribution of carnitine deficiency to geriatric frailty”. Ageing Research Reviews 9.3 (2010): 265-268.
- Jafari A., et al. “Potential nephroprotective effects of l-carnitine against drug-induced nephropathy: a review of literature”. ExpertOpinion on Drug Safety 12.4 (2013): 523-543
- Arslan C. “L-Carnitine and its use as a feed additive in poultry feeding a review”. Revue de Médecine Vétérinaire 157.3 (2006): 134-142.
- GolzarAdabiSH., et al. “L-carnitine and its functional effects in poultry nutrition”. World’s Poultry Science Journal 67.2 (2011): 277-295.
- Rebouche CJ. “Kinetics, pharmacokinetics, and regulation of L-carnitine and acetyl-L-carnitine metabolism”. Annals of the NewYork Academy of Sciences 1033 (2004): 30-41.
- Evans AM., et al. “Pharmacokinetics of L-carnitine”. Clinical Pharmacokinetics 42.11 (2003): 941-967.
- Lohninger A., et al. “L-Carnitine: new aspects of a known compound- A brief survey”. Monatshefte fur Chemie 136 (2005): 1255-1268.
- Krahenbuhl S. “L-carnitine and vegetarianism”. Annals of Nutrition & Metabolism 44.2 (2000):75-96.
- Walter P. “Introduction”. Annals of Nutrition & Metabolism 44.2 (2000): 77.
- Bohles H. “Basic concept of L-carnitine supplementation”. Annals of Nutrition & Metabolism 44.2 (2000): 77-78.
- Siebrecht S “L-carnitine: Physiological and pharmacological effects”. Annals of Nutrition & Metabolism 44.2 (2000): 79-80.
- Ferrari R., et al. “L-Carnitine and its Role in Medicine: From Function to Therapy”. London, England, Academic Press (1992).
- Rubio JC., et al. “Cerebrospinal fluid carnitine levels in patients with Alzheimer’s disease”. Journal of Neurological Sciences 155.2 (1998): 192-195.
- Stanley CA. “Carnitine deficiency disorders in children”. Annals of the New York Academy of Sciences 1033 (2004): 42-51.
- Hu ML. “Dietary polyphenols as antioxidants and anticancer agents: more questions than answers”. Chang Gung Medical Journal 34.5 (2011): 449-460.
- Bremer J. “Carnitine-metabolism and functions”. Physiological Reviews 63.4 (1983): 1420-1480.
- Calabrese V., et al. “Vitagenes, cellular stress response, and acetylcarnitine: relevance to hormesis”. BioFactors 35.2 (2009): 146-160.
- Pochini L., et al. “OCTN cation transporters in health and disease: role as drug targets and assay development”. Journal of Biomolecular Screening 18.8 (2013): 851-867.
- Ringseis R., et al. “Influence of pharmacological PPARalpha activators on carnitine homeostasis in proliferating and non-proliferating species”. Pharmacological Research 60.3 (2009): 179-184.
- Tamai I., et al. “Molecular and functional identification of sodium ion-dependent, high affinity human carnitine transporter OCTN2”. Journal of Biological Chemistry 273.32 (1998): 20378-20382.
- Vaz FM., et al. “Carnitine biosynthesis in mammals”. Biochemical Journal 361 (2002): 417-429.
- Ribas GS., et al. “L-carnitine supplementation as a potential antioxidant therapy for inherited neurometabolic disorders”. Gene 533.2 (2014): 469-476
- Surai PF, “Natural Antioxidants in Avian Nutrition and Reproduction” (2002). Nottingham University Press, Nottingham, UK.
- Surai PF. “Selenium in Nutrition and Health” (2006) Nottingham University Press, Nottingham, UK.
- Surai PF. “Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives”. Antioxidants 4 (2015): 204-247.
- Reznick AZ., et al. “Antiradical effects in L-propionyl carnitine protection of the heart against ischemia-reperfusion injury: the possible role of iron chelation”. Archives of Biochemistry and Biophysics 296.2 (1992): 394-401.
- Di Giacomo C., et al. “Effect of acetyl-L-carnitine on lipid peroxidation and xanthine oxidase activity in rat skeletal muscle”. Neurochemical Research 18.11 (1993): 1157-1162.
- Vanella A., et al. “L -propionyl-carnitine as superoxide scavenger, antioxidant, and DNA cleavage protector”. Cell Biology and Toxicology 16.2 (2000): 99-104.
- Galli G., et al. “Activation of apoptosis by serum deprivation in a teratocarcinoma cell line: inhibition by L-acetylcarnitine”. Experimental Cell Research 204.1 (1993): 54-60.
- Gülçin I. “Antioxidant and antiradical activities of L-carnitine”. Life Sciences 78.8 (2006): 803-811.
- Solarska K., et al. “The antioxidant properties of carnitine in vitro”. Cell and Molecular Biology Letters 15.1 (2010): 90-97.
- Kolodziejczyk J., et al. “L-Carnitine protects plasma components against oxidative alterations”. Nutrition 27.6 (2011): 693-699.
- Bell FP., et al. “Plasma and liver carnitine (free and esterified) levels and their interrelationships in moderately hypercholesterolemic monkeys (Macacaarctoides)”. Canadian Journal of Biochemistry and Cell Biology 61.6 (1983): 328-332.
- Bertelli A., et al. “L-propionyl carnitine protects erythrocytes and low-density lipoproteins against peroxidation”. Drugs underExperimental and Clinical Research 20.5 (1994): 191-197.
- Surai PF., et al. “Antioxidant-Prooxidant Balance in the Intestine: Applications in Chick Placement and Pig Weaning”. Journal ofVeterinary Science and Medicine 3 (2015): 1-16.
- Lin MT., et al. “Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases”. Nature 443.7113 (2006): 787-795.
- Sekine S., et al. “Mitochondrial proteolysis: Its emerging roles in stress responses”. Biochimica et Biophysica Acta 1850.2 (2015): 274-280.
- Calabrese V., et al. “Cellular stress responses, mitostress and carnitine insufficiencies as critical determinants in aging and neurodegenerative disorders: Role of hormesis and vitagenes”. Neurochemical Research 35.12 (2010): 1880-1915.
- Kumaran S., et al. “Age-associated decreased activities of mitochondrial electron transport chain complexes in heart and skeletal muscle: role of L-carnitine”. Chemico-Biological Interactions 148.1-2 (2004): 11-18.
- Kidd PM. “Neurodegeneration from mitochondrial insufficiency: nutrients stem cells, growth factors, and prospects for brain rebuilding using integrative management”. Alternative Medicine Review 10.4 (2005): 268-293.
- Silva-Adaya D., et al. “ Excitotoxic damage, disrupted energy metabolism, and oxidative stress in the rat brain: antioxidant and neuroprotective effects of L-carnitine”. Journal of Neurochemistry 105.8 (2008): 677-689.
- Shen W., et al. “Protective effects of R-alpha-lipoic acid and acetyl-L-carnitine in MIN6 and isolated rat islet cells chronically exposed to oleic acid”. Journal of Cell Biochemistry 104.4 (2008): 1232-1243.
- Zhu X., et al. “Acetyl-L-carnitine suppresses apoptosis of thioredoxin 2-deficient DT40 cells”. Archives of Biochemistry and Biophysics 478.2 (2008):154-160.
- Ye J., et al. “L-carnitine attenuates oxidant injury in HK-2 cells via ROS-mitochondria pathway”. Regulatory Peptides 161.1-3 (2010): 58-66.
- Zhang H., et al. “Combined R-alpha-lipoic acid and acetyl-L-carnitine exerts efficient preventative effects in a cellular model of Parkinson’s disease”. Journal of Cellular and Molecular Medicine 14.1-2 (2010): 215-225.
- Jun DW., et al. “Prevention of free fatty acid-induced hepaticlipotoxicity by carnitine via reversal of mitochondrial dysfunction”. Liver International 31 (2011): 1315-1324
- Wu J., et al. “TGF-β1 induces senescence of bone marrow mesenchymal stem cells via increase of mitochondrial ROS production”. BMC Developmental Biology 14 (2014): 21.
- Cao Y., et al. “Effects of L-carnitine on high glucose-induced oxidative stress in retinal ganglion cells”. Pharmacology” 94 (2014): 123-130.
- Zhang H., et al. “Combined R-alpha-lipoic acid and acetyl-L-carnitine exerts efficient preventative effects in a cellular model of Parkinson’s disease”. Journal of Cellular and Molecular Medicine 14.1-2 (2010): 215-25.
- Kumaran S., et al. “Age-associated deficit of mitochondrial oxidative phosphorylation in skeletal muscle: role of carnitine and lipoic acid”. Molecular and Cellular Biochemistry 280.1-2 (2005): 83-89.
- Karanth J., et al. “Effect of carnitine supplementation on mitochondrial enzymes in liver and skeletal muscle of rat after dietary lipid manipulation and physical activity”. Indian Journal of Experimental Biology 48.5 (2010): 503-510.
- Li J., et al. “Effects of exogenous carnitine on function of respiratory chain and antioxidant capacity in mitochondria of myocardium after exhaustive running in rats” Chinese Journal of Applied Physiology 28.5 (2012): 405-409.
- Pesce V., et al. “Acetyl-L-carnitine activates the peroxisome proliferator-activated receptor-γ coactivators PGC-1α/PGC-1β-dependent signaling cascade of mitochondrial biogenesis and decreases the oxidized peroxiredoxins content in old rat liver”. Rejuvenation Research 15.2 (2012): 136-139.
- Kizhakekuttu TJ., et al. “Natural antioxidants and hypertension: promise and challenges”. Cardiovascular Therapeutics 28.4 (2010): e20-32.
- Scioli MG., et al. “Propionyl-L-Carnitine is Efficacious in Ulcerative Colitis Through its Action on the Immune Function and Microvasculature”. Clinical and Translational Gastroenterology 5 (2014): e55.
- Doehner., et al. “Xanthine oxidase and uric acid in cardiovascular disease: Clinical impact and therapeutic options”. Seminars inNephrology 31.5 (2011): 433-440.
- Pacher P., et al. “Therapeutic effects of xanthine oxidase inhibitors: renaissance half a century after the discovery of allopurinol”. Pharmacological Reviews 58.1 (2006): 87-114.
- Volek JS., et al. “L-Carnitine L-tartrate supplementation favorably affects markers of recovery from exercise stress”. American Journal of Physiology. Endocrinology and Metabolism 282.2 (2002): E474-482.
- Loots DT., et al. “Acetyl-L-carnitine prevents total body hydroxyl free radical and uric acid production induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in the rat”. Life Sciences 75.10 (2004): 1243-1253.
- Spiering BA., et al. “Responses of criterion variables to different supplemental doses of L-carnitine L-tartrate”. Journal of Strengthand Conditioning Research 21.1 (2007): 259-264.
- Ho JY., et al. “L-Carnitine l-tartrate supplementation favorably affects biochemical markers of recovery from physical exertion in middle-aged men and women”. Metabolism 59.8 (2010): 1190-1199.
- Bedard K and Krause KH. “The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology”. PhysiologicalReviews 87.1 (2007): 245-313.
- Sinha N and Dabla PK. “Oxidative stress and antioxidants in Hypertension. A current review”. Current Hypertension Reviews 11.2 (2015): 132-142.
- Pignatelli P., et al. “Carnitine inhibits arachidonic acid turnover, platelet function, and oxidative stress”. American Journal of Physiology. Heart and Circulatory Physiology 284.1 (2003): 41-48.
- Chao HH., et al. “L-Carnitine attenuates angiotensin II-induced proliferation of cardiac fibroblasts: role of NADPH oxidase inhibition and decreased sphingosine-1-phosphate generation”. Journal of Nutritional Biochemistry 21.7 (2010): 580-588.
- Chao HH., et al. “L-carnitine reduces doxorubicin-induced apoptosis through a prostacyclin-mediated pathway in neonatal rat cardiomyocytes”. International Journal of Cardiology 146.2 (2011): 145-152.
- Scioli MG., et al. “Antioxidant treatment prevents serum deprivation- and TNF-α-induced endothelial dysfunction through the inhibition of NADPH oxidase 4 and the restoration of β-oxidation”. Journal of Vascular Research 51.5 (2014): 327-337.
- Stasi MA., et al. “Propionyl-L-carnitine improves postischemic blood flow recovery and arteriogenetic revascularization and reduces endothelial NADPH-oxidase 4-mediated superoxide production”. Arteriosclerosis, Thrombosis and Vascular Biology 30.3 (2010): 426-435.
- Miguel-Carrasco JL., et al. “Comparative effects of captopril and l-carnitine on blood pressure and antioxidant enzyme gene expression in the heart of spontaneously hypertensive rats”. European Journal of Pharmacology 632.1 (2010): 65-72.
- Rump TJ., et al. “Acetyl-L-carnitine protects neuronal function from alcohol-induced oxidative damage in the brain”. Free RadicalBiology and Medicine 49.10 (2010): 1494-1504.
- Pignatelli P., et al. “Effect of L-carnitine on oxidative stress and platelet activation after major surgery”. Acta AnaesthesiologicaScandinavica 55.8 (2011): 1022-1028.
- Zambrano S., et al. “The renoprotective effect of L-carnitine in hypertensive rats is mediated by modulation of oxidative stress-related gene expression”. European Journal of Nutrition 52.6 (2013): 1649-1659.
- Zambrano S., et al. “L-Carnitine protects against arterial hypertension-related cardiac fibrosis through modulation of PPAR-γ expression”. Biochemical Pharmacology 85.7 (2013): 937-944.
- Zambrano S., et al. “L-carnitine attenuates the development of kidney fibrosis in hypertensive rats by upregulating PPAR-γ”. Ameri-can Journal of Hypertension 27.3 (2014): 460-470.
- Dhaunsi GS., et al. “Carnitine prevents cyclic GMP-induced inhibition of peroxisomal enzyme activities”. Cell Biochemistry and Function 22.6 (2004): 365-371.
- Tastekin A., et al. “L-carnitine protects against glutamate- and kainic acid-induced neurotoxicity in cerebellar granular cell culture of rats”. Brain and Development 27.8 (2005): 570-573.
- Augustyniak A., et al. “The Influence of L-Carnitine on Oxidative Modification of LDL In Vitro”. Toxicology Mechanisms and Meth-ods18.6 (2008): 455-462.
- Li JL., et al. “Effects of L-carnitine against oxidative stress in human hepatocytes: involvement of peroxisome proliferator-activated receptor alpha”. Journal of Biomedical Science 19 (2012): 32.
- Yazaki T., et al. “L-carnitine improves hydrogen peroxide-induced impairment of nuclear maturation in porcine oocytes”. AnimalScience Journal 84.5 (2013): 395-402.
- Ye J., et al. “L-carnitine attenuates H2O2-induced neuron apoptosis via inhibition of endoplasmic reticulum stress”. NeurochemistryInternational 78 (2014): 86-95.
- Liu F., et al. “Ketamine-induced neuronal damage and altered N-methyl-D-aspartate receptor function in rat primary forebrain culture”. Toxicological Sciences 131.2 (2013): 548-557.
- Mescka CP., et al. “Prevention of DNA damage by L-carnitine induced by metabolites accumulated in maple syrup urine disease in human peripheral leukocytes in vitro”. Gene 548.2 (2014): 294-298.
- Deon M., et al. “Protective effect of L-carnitine on Phenylalanine-induced DNA damage”. Metabolic Brain Disease 30.4 (2015): 925-933.
- Liu F., et al. “Protective effect of acetyl-L-carnitine on propofol-induced toxicity in embryonic neural stem cells”. Neurotoxicology 42 (2014): 49-57.
- Manibusan MK., et al. “Postulated carbon tetrachloride mode of action: A review”. Journal of Environmental Science and Health. PartC, Environmental Carcinogenesis &. Ecotoxicology Reviews 25.3 (2007): 185-209.
- Raj S and Gothandam KM. “Hepatoprotective effect of polyphenols rich methanolic extract of Amorphophalluscommutatus var. wayanadensis against CCl4 induced hepatic injury in swiss albino mice”. Food and Chemical Toxicology 67 (2014): 105-112.
- Alkreathy HM., et al. “CCl4 induced genotoxicity and DNA oxidative damages in rats: Hepatoprotective effect of Sonchus arvensis”. BMC Complementary and Alternative Medicine 14 (2014): 452.
- Krishnappa P., et al. “Antioxidant and prophylactic effects of Delonix elata L., stem bark extracts, and flavonoid isolated quercetin against carbon tetrachloride-induced hepatotoxicity in rats”. BioMed Research International (2014) 507851.
- Shaker E., et al. “Silymarin, the antioxidant component and Silybummarianum extracts prevent liver damage”. Food and ChemicalToxicology 48.3 (2010): 803-806.
- Annadurai T., et al. “Acetyl-L-carnitine prevents carbon tetrachloride-induced oxidative stress in various tissues of Wistar rats”. Journal of Physiology and Biochemistry 67.4 (2011): 519-530.
- Cetinkaya A., et al. “The effects of L-carnitine and N-acetylcysteine on carbontetrachloride induced acute liver damage in rats”. Bratislavske Lekarske Listy 114.12 (2013): 682-688.
- Shaker ME., et al. “Comparison of vitamin E, L-carnitine and melatonin in ameliorating carbon tetrachloride and diabetes induced hepatic oxidative stress”. Journal of Physiology and Biochemistry 65.3 (2009): 225-233.
- Dos Santos NA., et al. “Cisplatin-induced nephrotoxicity and targets of nephroprotection: An update”. Archives of Toxicology 86.8 (2012): 1233-1250.
- Mansour HH., et al. “Silymarin modulates Cisplatin-induced oxidative stress and hepatotoxicity in rats”. Journal of Biochemistryand Molecular Biology 39.6 (2006): 656-661.
- El-Awady el-SE., et al. “Cisplatin-induced cardiotoxicity: Mechanisms and cardioprotective strategies”. European Journal of Pharmacology 650.1 (2011): 335-341.
- Al-Majed AA., et al. “Propionyl-L-carnitine prevents the progression of cisplatin-induced cardiomyopathy in a carnitine-depleted rat model”. Pharmacological Research 53.3 (2006): 278-286.
- Altun Z., et al. “Protective effect of acetyl-l-carnitine against cisplatin ototoxicity: role of apoptosis-related genes and pro-inflammatory cytokines”. Cell Proliferation 47.1 (2014): 72-80.
- Morigi M., et al. “Sirtuin 3-dependent mitochondrial dynamic improvements protect against acute kidney injury”. The Journal ofClinical Investigation 125.2 (2015): 715-726.
- Galicia-Moreno M and Gutiérrez-Reyes G. “The role of oxidative stress in the development of alcoholic liver disease”. Revista deGastroenterologia de Mexico 79.2 (2014): 135-144.
- Haorah J., et al.“Stabilization of superoxide dismutase by acetyl-l-carnitine in human brain endothelium during alcohol exposure: novel protective approach”. Free Radical Biology and Medicine 51.8 (2011): 1601-1609.
- Arafa HM and Sayed-Ahmed MM. “Protective role of carnitine esters against alcohol-induced gastric lesions in rats”. Pharmacological Research 48.3 (2003): 285-290.
- Dokmeci D., et al. “L-carnitine inhibits ethanol-induced gastric mucosal injury in rats”. Pharmacological Reports57.4 (2005): 481-488.
- Dobrzynska I., et al. “Effect of L-carnitine on liver cell membranes in ethanol-intoxicated rats”. Chemico-Biological Interactions 188.1 (2010): 44-51.
- Augustyniak A and Skrzydlewska E. “The influence of L-carnitine supplementation on the antioxidative abilities of serum and the central nervous system of ethanol-induced rats”. Metabolic Brain Disease 25.4 (2010): 381-389.
- Rattan SI. “The nature of gerontogenes and vitagenes. Antiaging effects of repeated heat shock on human fibroblasts”. Annals ofthe New York Academy of Sciences 854 (1998): 54-60.
- Calabrese V., et al. “Nitric oxide and cellular stress response in brain aging and neurodegenerative disorders: The role of vita-genes”. In Vivo18.3 (2004): 245-267.
- Calabrese V., et al. “Redox regulation of cellular stress response in aging and neurodegenerative disorders: Role of vitagenes”. Neurochemical Research 32.4 (2007): 757-773.
- Calabrese V., et al. “Redox regulation of cellular stress response by ferulic acid ethyl ester in human dermal fibroblasts:Role of vitagenes”. Clinical Dermatology 26.4 (2008): 358-363.
- Calabrese V., et al. “Vitagenes, dietary antioxidants and neuroprotection in neurodegenerative diseases”. Frontiers in Bioscience 14 (2009) 376-397.
- Calabrese V., et al. “Cellular stress responses, the hormesis paradigm, and vitagenes: Novel targets for therapeutic intervention in neurodegenerative disorders”. Antioxidants &. Redox Signaling 13.11 (2010): 1763-1811.
- Calabrese V., et al. “Cellular stress responses, hermetic phytochemicals and vitagenes in aging and longevity”. Biochimica et Biophysica Acta 1822.5 (2012): 753-783.
- Calabrese V., et al. “Sex hormonal regulation and hormesis in aging and longevity: Role of vitagenes”. Journal of Cell Communication and Signaling 8.4 (2014): 369-384.
- Cornelius C., et al. “Stress responses, vitagenes and hormesis as critical determinants in aging and longevity: Mitochondria as a “chi”. Immunity and Ageing 10.1 (2013): 15.
- Cornelius C., et al. “Hormesis and vitagenes in aging and longevity: Mitochondrial control and hormonal regulation”. HormoneMolecular Biology and Clinical Investigation 16.2 (2013): 73-89.
- Cornelius C., et al. “Osteoporosis and Alzheimer pathology:Role of cellular stress response and hormetic redox signaling in aging and bone remodelling”. Frontiers in Pharmacology 5 (2014): 120.
- Trovato Salinaro A., et al. “Cellular stress response, redox status, and vitagenes in glaucoma: A systemic oxidant disorder linked to Alzheimer’s disease”. Frontiers in Pharmacology 5 (2014): 129.
- Calabrese V., et al. “Acetylcarnitine induces hemeoxygenase in rat astrocytes and protects against oxidative stress: involvement of the transcription factor Nrf2”. Journal of Neuroscience Research 79.4 (2005): 509-521.
- Calabrese V., et al. “Redox modulation of heat shock protein expression by acetylcarnitine in aging brain: relationship to antioxidant status and mitochondrial function”. Antioxidants and Redox Signaling 8.4 (2006): 404-416.
- Calò LA., et al. “Antioxidant effect of L-carnitine and its short chain esters: relevance for the protection from oxidative stress related cardiovascular damage”. International Journal of Cardiology 107.1 (2006): 54-60.
- Calò LA., et al. “Carnitine-mediated improved response to erythropoietin involves induction of haem oxygenase-1: studies in humans and in an animal model”. Nephrology, Dialysis, Transplantation 23.3 (2008): 890-895.
- Abdul HM., et al. “Acetyl-L-carnitine-induced upregulation of heat shock proteins protects cortical neurons against amyloid-beta peptide 1-42-mediated oxidative stress and neurotoxicity: implications for Alzheimer’s disease”. Journal of Neuroscience Research 84.2 (2006): 398-408.
- Cao Y., et al. “Role of NF-E2-related factor 2 in neuroprotective effect of l-carnitine against high glucose-induced oxidative stress in the retinal ganglion cells”. Biomedicine & Pharmacotherapy 69 (2015): 345-348.
- Yang SP., et al. “Acetyl-l-carnitine prevents homocysteine-induced suppression of Nrf2/Keap1 mediated antioxidation in human lens epithelial cells”. Molecular Medicine Reports12.1 (2015): 1145-1150.
- Marcovina SM., et al. “Translating the basic knowledge of mitochondrial functions to metabolic therapy: role of L-carnitine”. Translational Research 161.2 (2013): 73-84.
- Miguel-Carrasco JL., et al. “Comparative effects of captopril and l-carnitine on blood pressure and antioxidant enzyme gene expression in the heart of spontaneously hypertensive rats”. European Journal of Pharmacology 632.3 (2010): 65-72.
- Chen HH., et al. “Peroxisome proliferator-activated receptor alpha plays a crucial role in L-carnitine anti-apoptosis effect in renal tubular cells”. Nephrology, Dialysis, Transplantation 24.10 (2009): 3042-3049.
- Surai PF. “Polyphenol compounds in the chicken/animal diet: from the past to the future”. Journal of Animal Physiology and Animal Nutrition 98.1 (2014): 19-31.
- Surai PF., et al. “Tissue-specific differences in antioxidant distribution and susceptibility to lipid peroxidation during development of the chick embryo”. Biochimica et Biophysica Acta 1304.1 (1996): 1-10.
- Zou W., et al. “Dietary L-carnitine enhances the lymphatic absorption of fat and alpha-tocopherol in ovariectomized rats”. Journalof Nutrition 135.4 (2005): 753-756.
- Clark RM., et al. “L-carnitine increases liver alpha-tocopherol and lowers liver and plasma triglycerides in aging ovariectomized rats”. Journal of Nutritional Biochemistry18.9 (2007): 623-628.
- Sachan DS., et al. “Decreasing oxidative stress with choline and carnitine in women”. Journal of the American College of Nutrition 24.3 (2005): 172-176.
- Augustyniak A and Skrzydlewska E. “L-Carnitine in the lipid and protein protection against ethanol-induced oxidative stress”. Alcohol 43.3 (2009): 217-223.
- Türker Y., et al. “Selenium and L-carnitine reduce oxidative stress in the heart of rat induced by 2.45-GHz radiation from wireless devices”. Biological Trace Element Research 143.3 (2011): 1640-1650.
- Moazzami AA., et al. “Changes in the metabolic profile of rat liver after α-tocopherol deficiency as revealed by metabolomics analysis”. NMR in Biomedicine 24.5 (2011): 499-505.
- Abidin Z., et al. “Ameliorative effects of L-carnitine and vitamin E (α-tocopherol) on haematological and serum biochemical parameters in White Leghorn cockerels given ochratoxin A contaminated feed”. British Poultry Science 54.4 (2013): 471-477.
- Arslan E., et al. “The additive effects of carnitine and ascorbic acid on distally burned dorsal skin flap in rats”. Medical ScienceMonitor 11.6 (2005): 176-180.
- Devi SA., et al. “Intermittent hypobaric hypoxia-induced oxidative stress in rat erythrocytes: protective effects of vitamin E, vitamin C, and carnitine”. Cell Biochemistry and Function 25.2 (2007): 221-231.
- Hagen TM., et al. “Mitochondrial decay in the aging rat heart: evidence for improvement by dietary supplementation with acetyl-L-carnitine and/or lipoic acid”. Annals of the New York Academy of Sciences 959 (2002): 491-507.
- Hagen TM., et al. “Feeding acetyl-L-carnitine and lipoic acid to old rats significantly improves metabolic function while decreasing oxidative stress”. Proceedings of the National Academy of Science of the United States of America 99.4 (2002): 1870-1875.
- McMackin CJ., et al. “Effect of combined treatment with alpha-Lipoic acid and acetyl-L-carnitine on vascular function and blood pressure in patients with coronary artery disease”. Journal of Clinical Hypertension 9.4 (2007): 249-255.
- Liu J., et al. “Age-associated mitochondrial oxidative decay: improvement of carnitine acetyltransferase substrate-binding affinity and activity in brain by feeding old rats acetyl-L- carnitine and/or R-alpha -lipoic acid”. Proceedings of the National Academy ofScience of the United States of America 99.4 (2002): 1876-1881.
- Savitha S., et al. “Efficacy of levocarnitine and alpha lipoic acid in ameliorating the decline in mitochondrial enzymes during aging”. Clinical Nutrition 24.5 (2005): 794-800.
- Savitha S and Panneerselvam C. “Mitochondrial membrane damage during aging process in rat heart: potential efficacy of L-carnitine and DL alpha lipoic acid”. Mechanisms of Ageing and Development127.4 (2006): 349-355.
- Abdul HM., et al. “Involvement of PI3K/PKG/ERK1/2 signaling pathways in cortical neurons to trigger protection by cotreatment of acetyl-L-carnitine and alpha-lipoic acid against HNE-mediated oxidative stress and neurotoxicity: implications for Alzheimer's disease”. Free Radical Biology and Medicine 42.3 (2007): 371-384.
- Tamilselvan J., et al. “Age-dependent upregulation of p53 and cytochrome c release and susceptibility to apoptosis in skeletal muscle fiber of aged rats: role of carnitine and lipoic acid”. Free Radical Biology and Medicine 43.12 (2007): 1656-1669.
- Atroshi F., et al. “Fumonisin B1-induced DNA damage in rat liver and spleen: effects of pretreatment with coenzyme Q10, L-carnitine, alpha-tocopherol and selenium”. Pharmacological Research 40.6 (1999): 459-467.
- Dhitavat S., et al. “Folate, vitamin E, and acetyl-L-carnitine provide synergistic protection against oxidative stress resulting from exposure of human neuroblastoma cells to amyloid-beta”. Brain Research1061.2 (2005): 114-117.
- Strzalkowski AK., et al. “The effect of supplementing sow with bioactive substances on neonatal small intestinal epithelium”. Journal of Physiology and Pharmacology 58.3 (2007): 115-122.
- Liu Z., et al. “Clinical effect of silymarin combined with levocarnitine on non-alcoholic fatty liver disease”. China Modern Medicine 7 (2009): R575.
- Fotina AA., et al. “Recent developments in usage of natural antioxidants to improve chicken production and quality”. BulgarianJournal of Agricultural Science 19.5 (2013): 889-896.
- Shatskih E., et al. “Molecular mechanisms and new strategies to fight stresses in egg-producing birds”. Agricultural Science andTechnology 7 (2015): 3-10.
- Surai PF., et al. “Antioxidant systems of the body: From vitamin E to polyphenols and beyond”. In Proc. 35th Western Nutrition Conference, Edmonton, Alberta (2014): 265-277.
- Velichko O., et al. “Effect of an antistress composition supplied with water on chick growth and development”. In Proc. XIVth European Poultry Conference, Stavanger, Norway (2014): 551.
- Fotina A., et al. “Effect of a water-soluble antistress composition on broiler chickens”. In Proc. XIVth European Poultry Conference, Stavanger, Norway (2014): 555.







.jpg&w=3840&q=75)