Tolerance of weanling pigs and effects on growth performance of supplementing corn-soybean meal-based diets with graded levels of a novel exogenous ß-mannanase
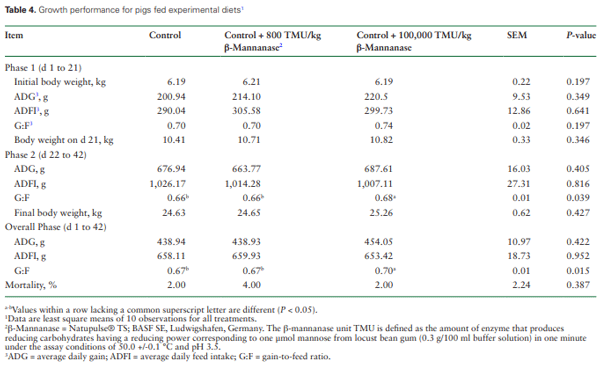
The hypothesis that a novel endo-β-mannanase can be used in diets for weanling pigs without negatively impacting growth performance, serum chemistry, hematological characters, or organ weights was tested. A total of 150 newly weaned pigs (75 castrated male and 75 female pigs; initial body weight: 6.20 ± 0.68 kg) were used. Pigs were allotted to three experimental diets (i.e., control, control plus 800 thermostable mannanase units (TMU)/kg, or control plus 100,000 TMU/kg). Pigs were allotted to pens with 5 pigs per pen for a total of 10 replicate pens per treatment. Pigs were fed phase 1 diets from d 1 to 21, and phase 2 diets from d 22 to 42 post-weaning. Average daily gain (ADG), average daily feed intake (ADFI), and gain:feed (G:F) were calculated. Blood samples from two pigs per pen (one male and one female pig) were collected on d 1, 21, and 42. One pig per pen from the control treatment and two pigs per pen from each of the β-mannanase treatments were euthanized at the end of the experiment and organs were collected. Data were analyzed using the Proc MIXED procedure of SAS with pen as the experimental unit. Results indicated that for the overall experiment, there were no differences in ADG, ADFI, or final body weight among treatments. However, pigs fed the diet with 100,000 TMU/kg of β-mannanase had greater (P < 0.05) G:F from d 22 to 42 and for the overall experimental period compared with pigs fed the control diet or the diet with 800 TMU/kg of β-mannanase. Most serum chemistry markers and blood hematological characters were not different among pigs fed experimental diets and concentrations were within the normal biological range for pigs. However, serum phosphorus was greater (P < 0.05) in pigs fed the diet with 100,000 TMU/kg of β-mannanase compared with pigs fed the other diets, but red cell distribution width and mean platelet volume were greater (P < 0.05) in pigs fed the control diet compared with pigs fed the control diet + 800 TMU/kg of β-mannanase. Abnormalities in liver, kidney, spleen, heart, stomach, or the small intestine were not observed, and the weight of these organs was not affected by dietary treatments. In conclusion, pigs fed diets containing 100,000 TMU/kg of β-mannanase had greater G:F from d 1 to 42 post-weaning compared with pigs fed control diets or the diets with 800 TMU/kg, and β-mannanase did not negatively impact general health and growth of the pigs even if included at a very high dose.
INTRODUCTION
MATERIALS AND METHODS
Animals, Housing, and Experimental Design
Blood Sample Collection and Chemical Analyses
Necropsy and Histopathology Examinations
Calculations and Statistical Analyses
RESULTS
Growth Performance
Serum Chemistry Markers
Hematological Characters
Necropsies and Organ Weights



DISCUSSION
CONCLUSIONS
- Agyekum, A. K., B. A. Slominski, and C. M. Nyachoti. 2012. Organ weight, intestinal morphology, and fasting whole-body oxygen consumption in growing pigs fed diets containing distillers dried grains with solubles alone or in combination with a multienzyme supplement. J. Anim. Sci. 90:3032–3040. doi:10.2527/jas.2011-4380
- AOAC Int. 2019. Official methods of analysis of AOAC Int. 21st ed. Rockville, MD, USA: AOAC Int.
- Bach Knudsen, K. E. 1997. Carbohydrate and lignin contents of plant materials used in animal feeding. Anim. Feed Sci. Technol. 67:319–338. doi:10.1016/S0377-8401(97)00009-6
- Baker, J. T., Z. Deng, A. Sokale, B. Frederick, and S. W. Kim. 2024. Nutritional and functional roles of β-mannanase on intestinal health and growth of newly weaned pigs fed two different types of feeds. J. Anim. Sci. 102:skae206. doi:10.1093/jas/skae206
- Campbell, J. M., J. D. Crenshaw, and J. Polo. 2013. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 4:19. doi:10.1186/2049-1891-4-19
- Chegeni, A., M. Torki, and A. Kamyab. 2011. Effects of β-mannanase-based enzyme in corn-soy and corn-soy-canola diets on broiler performance. J. Appl. Anim. Res. 39:261–268. doi:10.1080/09712119.2011.605319
- Chen, J., C. S. Robb, F. Unfried, L. Kappelmann, S. Markert, T. Song, J. Harder, B. Avci, D. Becher, P. Xie, et al. 2018. Alpha- and beta-mannan utilization by marine Bacteroidetes. Environ. Microbiol. 20:4127–4140. doi:10.1111/1462-2920.14414
- Choi, H., S. Y. Ji, H. Jo, M. Song, and B. G. Kim. 2021. Excessive dietary lead reduces growth performance and increases lead accumulation in pigs. Anim. Biosci. 34:102–108. doi:10.5713/ajas.20.0220
- Cooper, C. A., L. E. Moraes, J. D. Murray, and S. D. Owens. 2014. Hematologic and biochemical reference intervals for specific pathogen free 6-week-old Hampshire-Yorkshire crossbred pigs. J. Anim. Sci. Biotechnol. 5:5. doi:10.1186/2049-1891-5-5
- de Vries, R. P., and J. Visser. 2001. Aspergillus enzymes involved in degradation of plant cell wall polysaccharides. Microbiol. Mol. Biol. Rev. 65:497–522. doi:10.1128/MMBR.65.4.497-522.2001
- Fickler, A., K. Kore, A. Matthews, D. Moore, and A. B. Mandal. 2023. Efficacy of a novel β-mannanase on intestinal digesta viscosity in broiler chickens fed diets with high levels of guar meal (Cyamopsis tetragonoloba). Indian J. Poult. Sci. 58:123–128. doi:10.5958/0974-8180.2023.00022.3
- Hsiao, H. Y., D. M. Anderson, and N. M. Dale. 2006. Levels of β-mannan in soybean meal. Poult. Sci. 85:1430–1432. doi:10.1093/ps/85.8.1430
- Huntley, N. F., C. M. Nyachoti, and J. F. Patience. 2018. Lipopolysaccharide immune stimulation but not β-mannanase supplementation affects maintenance energy requirements in young weaned pigs. J. Anim. Sci. Biotechnol. 9:47. doi:10.1186/s40104-018-0264-y
- Iowa State University Clinical Pathology Laboratory. 2011. Reference intervals. https://vetmed.iastate.edu/vpath/services/diagnosticservices/clinical-pathology/testing-and-fees/reference-intervals/ (Accessed September 2024)
- Jackson, M. E., K. Geronian, A. Knox, J. McNab, and E. McCartney. 2004. A dose-response study with the feed enzyme beta-mannanase in broilers provided with corn-soybean meal based diets in the absence of antibiotic growth promoters. Poult. Sci. 83:1992–1996. doi:10.1093/ps/83.12.1992
- Jang, J., K. H. Kim, Y. D. Jang, and Y. Y. Kim. 2020. Effects of dietary β-mannanase supplementation on growth performance, apparent total tract digestibility, intestinal integrity, and immune responses in weaning pigs. Animals. 10:703. doi:10.3390/ani10040703
- Jang, K. B., Y. I. Kim, M. E. Duarte, and S. W. Kim. 2024. Effects of β-mannanase supplementation on intestinal health and growth of nursery pigs. J. Anim. Sci. 102:skae052. doi:10.1093/jas/skae052
- Kaneko, J., J. Harvey, and M. Bruss. 1997. Clinical biochemistry of domestic animals. San Diego, CA, USA: Academic Press.
- Kiarie, E., L. Romero, and C. Nyachoti. 2013. The role of added feed enzymes in promoting gut health in swine and poultry. Nutr. Res. Rev. 26:71–88. doi:10.1017/S0954422413000048
- Kiarie, E. G., S. Steelman, M. Martinez, and K. Livingston. 2021. Significance of single β-mannanase supplementation on performance and energy utilization in broiler chickens, laying hens, turkeys, sows, and nursery-finish pigs: a meta-analysis and systematic review. Transl. Anim. Sci. 5:txab160. doi:10.1093/tas/txab160
- Kim, J. S., S. L. Ingale, A. R. Hosseindoust, S. H. Lee, J. H. Lee, and B. J. Chae. 2017. Effects of mannan level and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites of growing pigs. Animal. 11:202–208. doi:10.1017/S1751731116001385
- Kim, J. S., S. L. Ingale, S. H. Lee, K. H. Kim, J. S. Kim, J. H. Lee, and B. J. Chae. 2013. Effects of energy levels of diet and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites of growing pigs. Anim. Feed Sci. Technol. 186:64–70. doi:10.1016/j.anifeedsci.2013.08.008
- Koepke, J. R., R. S. Kaushik, W. R. Gibbons, M. Brown, and C. L. Levesque. 2017. Evaluation of a bioprocessed soybean meal on nursery pig performance and immune status. J. Anim. Sci. 95:5030–5039. doi:10.2527/jas2017.1679
- Lee, J. T., and K. D. Brown. 2022. Mannanase, alpha-galactosidase and pectinase: minor player or yet to be exploited? In: Bedford, M. R. and G. G. Partridge, editors, Enzymes in farm animal nutrition. Oxfordshire, UK: CAB International. doi:10.1079/9781789241563.0005
- Lee, S. A., C. Kong, O. Adeola, and B. G. Kim. 2016. Different coefficients and exponents for metabolic body weight in a model to estimate individual feed intake for growing-finishing pigs. Asian-Australas. J. Anim. Sci. 29:1756–1760. doi:10.5713/ajas.16.0420
- Lessard, P. A., X. Li, J. N. Broomhead, M. H. Parker, C. Bailey, and R. M. Raab. 2021. Properties of corn-expressed carbohydrase AC1 in swine diets and its effects on apparent ileal digestibility, performance, hematology, and serum chemistry. Heliyon. 7:e07696. doi:10.1016/j.heliyon.2021.e07696
- Lindemann, M. D., and B. G. Kim. 2007. Technical note: A model to estimate individual feed intake of swine in group feeding. J. Anim. Sci. 85:972–975. doi:10.2527/jas.2006-412
- NRC. 2012. Nutrient requirements of swine. 11th rev. ed. Washington, DC, USA: National Academies Press.
- Oxley, D., G. Currie, and A. Bacic. 2004. Monosaccharide composition analysis: alditol acetates. In: Simpson, R. J. editor, Purifying proteins for proteomics. Cold Spring Harbor, NY, USA: Cold Spring Harbor Laboratory Press.
- Pariza, M. W., and M. Cook. 2010. Determining the safety of enzymes used in animal feed. Regul. Toxicol. Pharmacol. 56:332–342. doi:10.1016/j.yrtph.2009.10.005
- Pettey, L. A., S. D. Carter, B. W. Senne, and J. A. Shriver. 2002. Effects of β-mannanase addition to corn-soybean meal diets on growth performance, carcass traits, and nutrient digestibility of weanling and growing-finishing pigs. J. Anim. Sci. 80:1012–1019. doi:10.2527/2002.8041012x
- Pond, W. G., V. H. Varel, J. S. Dickson, and W. M. Haschek. 1989. Comparative response of swine and rats to high-fiber or high-protein diets. J. Anim. Sci. 67:716–723. doi:10.2527/jas1989.673716x
- Rymut, H. E., L. A. Rund, C. R. Bolt, M. B. Villamil, B. R. Southey, R. W. Johnson, and S. L. Rodriguez-Zas. 2021. The combined effect of weaning stress and immune activation during pig gestation on serum cytokine and analyte concentrations. Animals. 11:2274. doi:10.3390/ani11082274
- Sánchez-Uribe, P., et al. 2022. Effect of β-mannanase addition during whole pigs fattening on production yields and intestinal health. Animals. 12:3012. doi:10.3390/ani12213012
- Schliffka, W., et al. 2019. Safety and efficacy evaluation of a novel dietary muramidase for swine. Heliyon. 5:e02600. doi:10.1016/j.heliyon.2019.e02600
- Tajudeen, H., et al. 2025. The immunomodulatory activities of a thermostable galacto-mannanase and their impact on the performance of weaned piglets. Anim. Feed Sci. Technol. 319:116186. doi:10.1016/j.anifeedsci.2024.116186
- Tukey, J. W. 1977. Exploratory data analysis. Boston, MA, USA: Addison-Wesley.
- Wilson, G. D. A., D. G. Harvey, and C. R. Snook. 1972. A review of factors affecting blood biochemistry in the pig. Br. Vet. J. 128:596–610. doi:10.1016/s0007-1935(17)36632-0




















United States

