Introduction
Cereal grains are the principal component in swine diets due to the efficiency of cost per calorie provided compared to other ingredients. Nevertheless, fungal infection often occurs in the field and during grain storage, and these fungi leave behind secondary metabolites known as mycotoxins, which can have adverse effects on livestock if ingested in sufficient quantities. The bioavailability of some of the major mycotoxins of concern (e.g. aflatoxins or zearalenone) can be reduced by including inexpensive adsorbent compounds, such as specific clays or hydrated sodium calcium aluminosilicates, in the diet. The inclusion of these compounds results in a reduction in mycotoxin uptake and decreased distribution to the blood and target organs (EFSA, 2009).
However, according to a 3-year survey of global mycotoxin occurrence (Rodrigues and Naehrer, 2012), the most prevalent (65% of finished feed) mycotoxin in North American feedstuffs is deoxynivalenol (DON), which is known for its feed suppression and immunomodulatory effects in pigs when present in diets at more than 1 ppm. Despite DON’s prevalence and known effects, adsorbent compounds have proven largely ineffective against DON in both in vitro models and in vivo growth studies (Dänicke, 2000). Although no DON-detoxifying agents have efficacy claims that are approved by the U.S. Food and Drug Administration, some products are reported to be of benefit in field studies. One such compound is Product X (Nutriquest LLC, Mason City, IA), a proprietary blend of adsorbent clays and preservatives. Because nursery pigs are known for their sensitivity to anti-nutritional factors such as mycotoxins, the aim of the experiment was to test the growth performance of nursery pigs fed a naturally DON-contaminated diet in the presence or absence of Product X, and to investigate the effects of DON absorption and excretion using a urinary balance model. Sodium metabisulfite (SMB; Na2 S2 O5 ), a known biotransforming agent of DON that reacts with DON in the presence of heat and moisture to form a non-toxic DON-sulfonate adduct (DONS; Beyer et al., 2010) and sulfur dioxide gas, was also incorporated into naturally contaminated diets to further evaluate SMB’s potential for use in naturally DON-contaminated diets.
Procedures
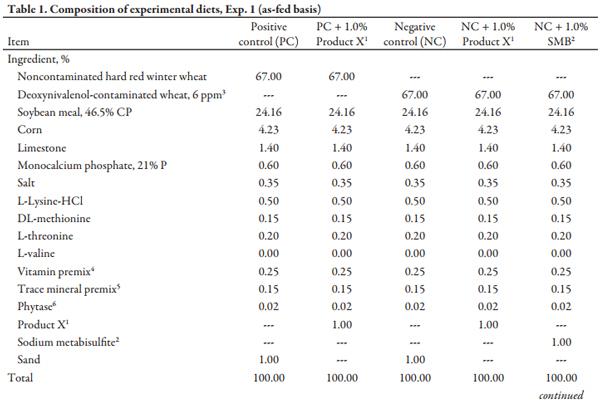
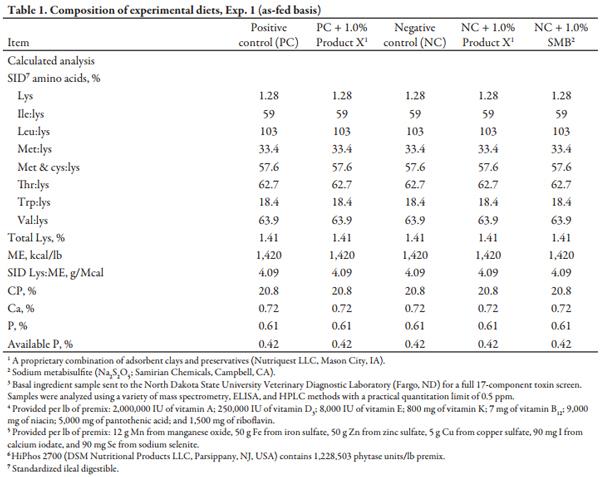
The Kansas State University Institutional Animal Care and Use Committee approved the protocol used in this experiment. Sources of naturally DON-contaminated hard red winter (HRW) wheat and low-DON HRW wheat were acquired, and an initial 17-component mycotoxin screen was performed at North Dakota State University Veterinary Diagnostic Laboratory (NDSU) using a combination of mass spectrometry, ELISA, and HPLC methods. Based on the analyzed DON concentration, an equal amount of high-DON (6.03 ppm) or low-DON (0.05 ppm DON) wheat was incorporated into experimental diets to achieve desired DON concentrations. Wheat sources were hammermill ground to approximately 600µ, and each source was homogeneously blended to minimize any variation in DON concentration between diets. Diets were formulated to meet or exceed NRC (2012) requirements and to be identical in nutrient composition apart from DON concentration and the inclusion of detoxifying agents (Table 1). Diets for the growth performance and urinary balance experiment were manufactured simultaneously at the Kansas State University O. H. Kruse Feed Mill. The 5 experimental diets were: 1) positive control (PC; <0.5 ppm DON), 2) PC + 1.0% Product X, 3) negative control (NC; 4.0 ppm DON), 4) NC + 1.0% Product X, and 5) NC + 1.0% SMB (Na2 S2 O5; Samirian Chemicals, Campbell, CA). Two large batches using the low- or high-DON wheat were initially mixed to ensure consistency in DON concentrations. Each individual diet was then manufactured by subdividing the large batches and incorporating the appropriate detoxifying agent or sand at 1.0% of the final diet.
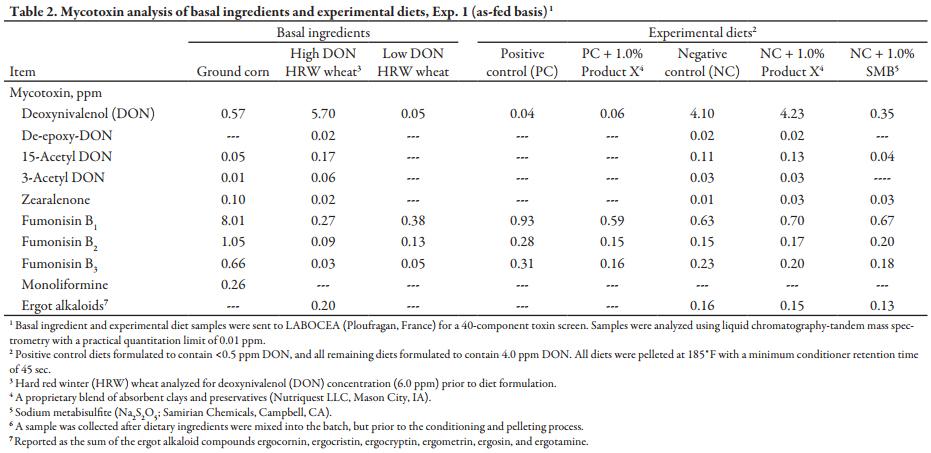
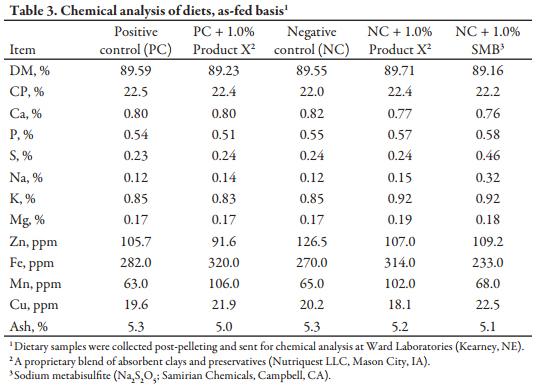
After mixing complete diets for 2 minutes in a double ribbon mixer, diets were pelleted (CPM Master Model 1000HD; Crawfordsville, IN) at a production rate of 1000 lb/h to maintain a minimum conditioner retention time and temperature of 45 seconds and 180°F, respectively. Diets were manufactured in numeric order to minimize carryover, with a flush between each diet. All personnel involved were required to wear respirators and safety goggles during the pelleting process as sodium metabisulfite releases sulfur dioxide gas in the presence of heat and moisture, which can irritate the eyes and respiratory tract. Samples of each diet were collected both pre- and post-pelleting. Diet samples were stored, frozen, and shipped along with basal ingredient samples to LABOCEA (Ploufragan, France) for a full mycotoxin screen (Table 2) and to Ward Laboratories (Kearney, NE) for chemical analysis (Table 3).
Growth experiment
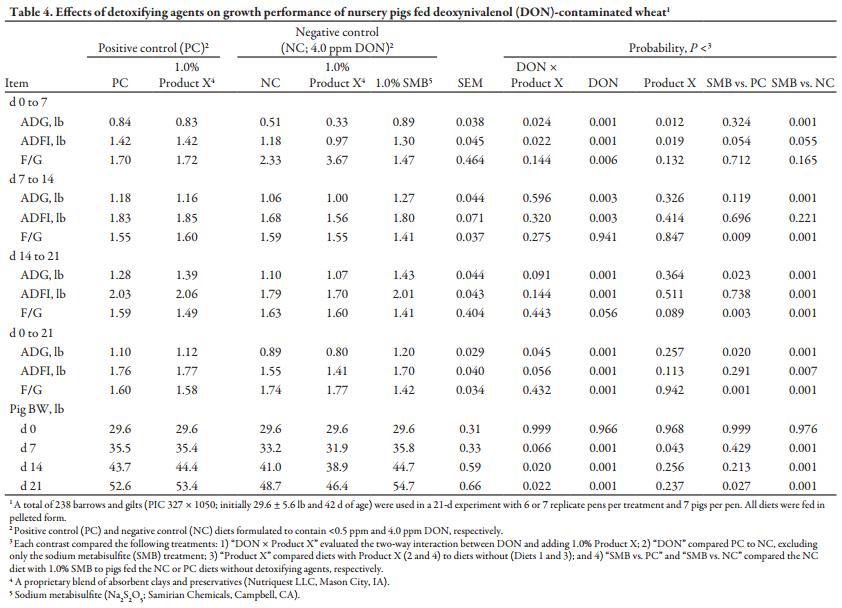
A total of 238 barrows and gilts (PIC 327 × 1050; initially 29.6 ± 5.6 lb and 40d of age) were used in a 21-d growth study with 7 replicate pens per treatment and 7 pigs per pen; however, based on limited pen availability, 1 treatment (PC) had 6 replicate pens. Pigs were allotted to pens by initial weight at weaning, and when pigs reached approximately 30 lb, they were reweighed and pen average pig weight was balanced across 1 of 5 treatments in a completely randomized design with a 2 × 2 + 1 factorial arrangement. Deoxynivalenol and Product X inclusion served as main effects with an additional treatment including SMB. Each pen contained a 4-hole, dry self-feeder and a nipple waterer to provide ad libitum access to feed and water. Experimental diets were fed for 21 d with ADG, ADFI, and F/G determined by weighing pigs and measuring feed disappearance on d 7, 14, and 21 (Table 4).
Urinary balance experiment
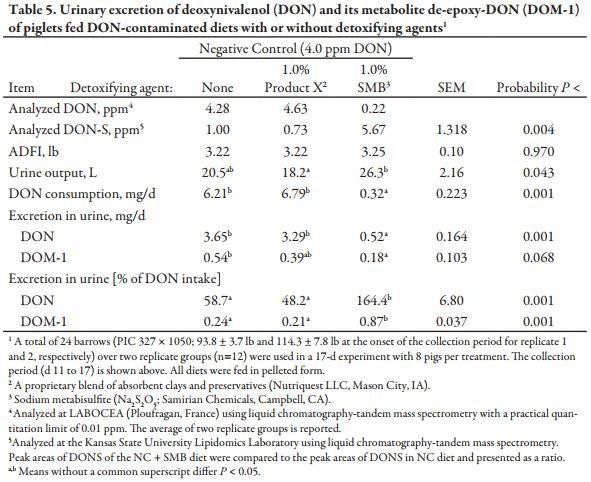
A balance study was also conducted involving pigs individually housed in stainless-steel metabolism cages (4.9 × 2.0 ft) in an environmentally controlled building. Each cage was equipped with a feeder and a nipple drinker for ad libitum access to water. To determine the effects of Product X and SMB on DON urinary excretion and metabolism, only the 3 NC diets from the growth experiment were included. A total of 24 barrows were used for two replicate groups (12 pigs per group), with 4 pigs per dietary treatment in each group. Pigs were allotted to treatments in a randomized complete block design based on initial BW and location within the experimental room. Pigs were adapted to the diets and to an amount of feed taken up completely by all pigs (3.0 and 3.5 lb for groups 1 and 2, respectively) and to the metabolism cages during a 10-d period. Then, during a 7-d collection period, daily feed intake and urinary output were recorded quantitatively (Table 5). The mean initial BW at the start of the collection period was 93.8 ± 3.7 lb and 114.3 ± 7.8 lb for groups 1 and 2, respectively. Feed allocation was divided into two equal amounts and given twice daily at 0700 and 1500. Due to the low recovery of DON and its primary metabolite de-epoxy-DON (DOM-1) in feces (0.1 to 1.7% of DON intake) in similar studies (Danicke et al., 2007, Danicke et al., 2012), fecal DON and fecal DOM-1 were not analyzed in the present experiment. The separation of feces from urine was achieved by using differently sized screens located beneath the slatted floor of the cage and connected to a funnel and urine collection bottle. Each pig’s total daily urine output was frozen and then thawed and homogeneously mixed at the end of the collection period. A representative aliquot sample was collected and then frozen before being sent for a full mycotoxin screen at LABOCEA (Ploufragan, France) using liquid chromatography-tandem mass spectrometry with a practical quantitation limit of 0.01 ppm.
DON-sulfonate quantification
The primary objective of DONS analysis was to confirm that the decreased analyzed DON in the pelleted NC + SMB diet was due to DON structural modification to form DONS, as demonstrated in prior research. An automated electrospray ionization-tandem mass spectrometry (ESI-MS/MS) approach was used, and data acquisition and analysis were carried out as in Beyer et al. (2010). Unfractionated DONS extracts were introduced by continuous infusion into the ESI source on a triple quadrupole MS/MS (4000QTrap, Applied Biosystems, Foster City, CA). An aliquot of 75 µl of extract in methanol/water (3/1 vol/vol) was introduced using an autosampler (LC Mini PAL, CTC Analytics AG, Zwingen, Switzerland) fitted with the required injection loop for the acquisition time and presented to the ESI needle at 30 µl/min. A negative neutral loss scan of 80.9 was used to detect the DON-S molecular ion 377 [M-H]-. The ESIMS/MS parameters used were: DP -80, EP -10, CE-36, CXP -15, electrospray capillary voltage -4500, collision gas pressure 2 (arbitrary units), interface heater on, source temperature (heated nebulizer) 572°F, curtain gas 20 and both ion source gases 45 (arbitrary units). Seventy-five continuum scans were averaged in multiple channel analyzer mode (MCA).
The background of each spectrum was subtracted, the data were smoothed, and peak areas were integrated using Applied Biosystems Analyst software. For both replicate groups of the urinary balance experiment, samples of each diet (n = 3) were analyzed in triplicate. Peak areas of DONS of the NC+SMB diet were compared to the peak areas of DONS in the NC diet and presented as a ratio.
Statistical analysis
Data collected from both experiments were analyzed using analysis of variance in the MIXED procedure of SAS, version 9.1 (SAS Institute Inc., Cary, NC). In the growth experiment, treatment effects were assessed within each experimental period using pen as the experimental unit. The fixed factors in the model were DON level and the presence or absence of Product X. The pre-planned contrasts in the growth experiment included: 1) interactions between DON and Product X, 2) DON vs. noncontaminated, and 3) the absence or presence of Product X in diets. Finally, two pairwise comparison contrasts were used to evaluate the effects of 1) adding SMB to DON-contaminated diets and 2) NC + SMB versus non-contaminated diets with no detoxifying agents present.
The urinary balance experiment was analyzed as a randomized complete block design with individual pig as the experimental unit. Data from the two replicates were combined and analyzed for replicate × treatment interactions. Due to a lack of significant interactions, the data were combined and analyzed with replicate and block included in the model as random effects. For all data analysis, when a significant overall treatment difference was found, differences among treatments were determined using the PDIFF statement in SAS. Overall significant differences were set at P < 0.10 and differences among treatments declared at P < 0.05, with tendencies reported when 0.05 ≤ P ≤ 0.10.
Results and Discussion
Mycotoxin analyses of the high-DON and low-DON wheat at LABOCEA generally matched initial analyses from NDSU showing only minimal co-contamination from other mycotoxins. However, the ground corn used across all diets contained a low level of DON (0.57 ppm) and a high level of fumonisin B1 (FUM; 8.01 ppm), which is above cautionary levels for swine. Interactive effects between DON and FUM are well documented (Grenier et al., 2011) and cannot be ruled out completely, but the low inclusion rate (4%) of FUM-contaminated corn in experimental diets makes the impact of any interactive effects likely minimal on experimental outcomes. The analyzed concentration of DON in final diets, in general, matched anticipated levels, with the NC + SMB diet the only exception (0.35 ppm). To reiterate, all 3 NC diets were initially prepared as a single, large batch to ensure consistent DON levels. That large batch was then split and the appropriate detoxifying agent or sand was incorporated prior to pelleting. The decrease in analyzed DON is likely attributed to the formation of 5-fold greater (P < 0.01) ratio of DONS present in the NC + SMB diet compared to the NC alone. DONS is a non-toxic product formed by the reaction between SMB and DON, which is amplified by hydrothermal environmental conditions (Danicke et al. 2005), such as those present in the pelleting conditioner in this study. The presence of low levels of other toxins in experimental diets is most likely inconsequential, as concentrations were all well below cautionary limits for growing swine. Nutrient analyses for CP, Ca, P, and ash content were consistent across experimental diets. Adding 1.0% Product X increased Fe and Mn levels in the diet by approximately 15 and 60% compared to those without. Furthermore, the addition of 1.0% SMB increased dietary S and Na concentrations approximately 2-fold versus other diets.
Growth experiment
From d 0 to 7, a two-way interaction for ADFI was detected in which adding Product X worsened ADG and ADFI (P < 0.05) by a greater magnitude in DON-contaminated diets than PC diets. The presence of DON in diets decreased ADG by 52% (P < 0.001), driven by 24% lower ADFI (P < 0.001) and 56% poorer F/G (P < 0.01). However, the addition of SMB to the NC diet markedly improved ADG (P < 0.001) and tended to improve ADFI (P < 0.10) versus the NC alone. Nevertheless, from d 0 to 7, pigs fed the NC + SMB diet still tended to have decreased ADFI (P < 0.10) versus pigs fed the PC.
From d 7 to 14, no DON × Product X interactions were present. The previously observed worsening of F/G for NC-fed pigs was not present and DON’s impact on ADFI was less marked than in the initial period. However, pigs fed NC diets still experienced the anorexic effects typically associated with DON, which reduced ADFI (P < 0.01) and decreased ADG (P < 0.01) relative to pigs fed the PC. Adding Product X to diets had no effect on ADG, ADFI or F/G, but the addition of SMB improved ADG (P < 0.001) by 20% compared to the NC, driven primarily by an improvement (P < 0.001) in F/G. Pigs fed the NC + SMB diet also exhibited 11% greater F/G (P < 0.01) than pigs fed PC diets during the second period.
From d 14 to 21, a tendency for a two-way interaction was detected (P < 0.10) for ADG in which Product X inclusion increased ADG in PC diets, but worsened ADG in NC diets. Average daily gain was decreased (P < 0.001) for pigs fed the NC, again driven by reduced ADFI (P < 0.001) but also by poorer F/G (P < 0.05). Product X addition tended to worsen F/G (P < 0.10), while ADG and ADFI were not affected. SMB supplementation in NC diets improved ADG, ADFI, and F/G (P < 0.001) by the greatest magnitude during the third period. Pigs fed the NC + SMB also had increased ADG (P < 0.05) compared to pigs fed the PC, driven by an 11% improvement in F/G (P < 0.01).
Overall, a two-way interaction was observed for ADG and final BW in which Product X supplementation worsened ADG and final BW (P < 0.05) and tended to worsen ADFI (P < 0.10) in NC diets but did not affect performance in PC diets. Feeding 4 ppm DON in NC diets decreased ADG (24%; P < 0.001) and final BW (P < 0.001) during the experimental period, reducing ADFI by 16% and worsening F/G (P < 0.001) by 10%. Supplementing 1.0% SMB in the NC diet improved ADG, ADFI, and F/G (P < 0.01) over NC alone by 35, 10, and 19%, respectively, resulting in a 12% improvement (P < 0.001) in final BW. Unexpectedly, ADG and final BW of pigs fed the NC + SMB diet surpassed even those of pigs fed the noncontaminated PC diet (P < 0.05), primarily driven by an 11% improvement in F/G (P < 0.001).
These results reiterate the extent to which high-DON diets can negatively impact nursery pig growth performance. The present data agree with findings of Etienne and Waché (2008), who cited a 4.6% decrease in ADFI for every 1 ppm of DON in the diet, and Frobose et al. (2015), who described the feed intake suppression pattern as being the most marked during the initial exposure period and lessening over time. These anorexic effects of DON are most frequently attributed to changes in: metabolism and concentration of brain transmitters such as serotonin in cerebrospinal fluid (Prelusky and Trenholm, 1993), inhibition of small-intestinal motility (Rotter et al., 1996), and development of conditioned taste aversion to DON (Ossenkopp et al., 1994). The effects are more severe in pigs than other species due to a more rapid absorption and tissue distribution of DON (15 to 30 m) coupled with delayed clearance of DON from cerebrospinal fluid (Prelusky et al., 1990). Previous reports of the impact of DON on F/G have been more variable (Rotter et al., 1996), but a series of four growth experiments (Frobose et al., 2015) consistently observed depressed F/G during the initial period, which is consistent with the reduction in F/G seen from d 0 to 7 in the growth experiment. This may be associated with wasted feed from pigs sorting feed due to taste aversion and may also be connected with immune system stimulation during the initial exposure, which could increase maintenance requirements. After this initial decrease, F/G of pigs fed DON-contaminated diets was generally similar to those fed the PC diet.
In this study, the addition of Product X at 1.0% in DON-contaminated diets did not alleviate DON’s negative effects on nursery pig growth. While Product X did not affect growth when added to the PC diet, intriguingly, when Product X was added to highDON diets, ADG was suppressed by an additional 11%, mainly driven by 9% lower ADFI. Although the negative DON × Product X interaction was unexpected, adsorbent clays have shown little efficacy in previous research on DON and some adsorbing agents have been reported to be nonselective in that they may affect the utilization of essential nutrients, such as vitamins and minerals (EFSA, 2009).
Pelleting NC diets with 1.0% SMB restored the DON-associated reduction in ADFI, which agrees with previous research (Frobose et al., 2011) and is most likely associated with the greater than ten-fold reduction in analyzed DON levels and the probable conversion to the nontoxic DONS. However, pelleting NC diets with SMB also resulted in consistent improvement in F/G throughout the duration of the experiment versus not only the NC (18%) but also compared to pigs fed the noncontaminated PC diet (11%), which suggests that the benefit may be independent of the DON-contamination of the diet. While the biological mechanism remains unclear, the efficiency benefit is consistent with Danicke et al. (2005) and implies that the hydrothermal treatment of SMB improved the nutrient availability for the animal.
Despite SMB being generally recognized as safe (GRAS status) by the FDA, the release of sulfur dioxide when pelleting diets containing SMB is an additional concern for feed mill employees. It can be irritating to the eyes and respiratory tract and may require the use of protective equipment. Moreover, the addition of 1.0% SMB increased the S and Na concentrations in the diet, and based on results of Til et al. (1972), SMB destroys thiamine and long-term supplementation may result in reductions in growth performance and potentially sulfur-induced thiamine deficiency. As a result, additional research is necessary to determine the minimum SMB level necessary and acceptable feeding duration to minimize feed processing and thiamine deficiency concerns.
Urinary balance experiment
The experimental diets used in the urinary balance experiment were sampled daily within each replicate, and a subsample of each was sent for mycotoxin analysis at LABOCEA. Analyzed DON concentrations were generally similar to those used in the growth study. Daily feed intake was set by the amount of feed consumed daily by NC-fed pigs during the 10-d adaptation period, and no differences in feed disappearance were observed between treatments during the collection period. Pigs fed the NC + SMB diet had the greatest urine output during the collection period, being significantly greater (P < 0.05) than pigs fed NC + Product X, with NC pigs intermediate. This additional urinary excretion is likely due to increased water intake due to the elevated dietary Na level when 1.0% SMB was incorporated into the diet.
As calculated from analyzed DON levels, pigs fed NC and NC + Product X treatments consumed a greater amount of DON per d (P < 0.001) than pigs fed the NC + SMB diet, since DON conversion to DONS occurred during feed manufacturing when SMB was added before pelleting. The DONS analysis confirmed that DON to DONS conversion was over 5-fold greater (P < 0.01) when 1.0% SMB was added to NC diets prior to pelleting versus the NC alone and NC + Product X. Although DONS is known to lack the emetic activity of DON (Young et al., 1987), interestingly, the addition of SMB to NC diets did not reduce the incidence of vomiting. In fact, NC + SMB pigs vomited on 10 occasions as compared to 7 and 3 for the NC and NC + Product X treatments, respectively (data not shown). Still, the daily DON urinary excretion was reduced (P < 0.001) for NC + SMB fed pigs versus the NC and NC + Product X, and the excretion of the primary metabolite DOM-1 was also less (P < 0.05) in the NC + SMB pigs. However, when expressed as a percentage of daily DON intake, pigs fed the NC + SMB diet excreted more DON than they consumed (164%), which was greater (P < 0.001) than pigs fed NC (59%) or the NC + Product X (48%) diet.
For pigs fed the NC + SMB treatment, DON recovery greater than 100% appears to indicate that some of the DONS was degraded to the parent DON. Recent work by Schwartz et al. (2013) revealed that 3 structurally unique forms of DONS can be formed by the reaction of DON with SMB, dependent on the sulfiting agent and processing conditions present. While DONS-1 and DONS-2 are stable across a broad pH range, DONS-3 can decompose to DON at alkaline pH, such as those in the proximal small intestine. Schwartz-Zimmerman et al. (2014) compared sulfiting agents in a follow-up study and found the predominant form produced by the reaction between DON and SMB to be DONS-3. If the sulfonate formation profile was similar in the present study, this would explain the degradation of a portion of DONS-3 back into DON, which would then be detected as additional DON in the urine. Since the gross DON urine recovery remained only 15% of the DON consumed by pigs fed the NC or NC + Product X diets, the physiological impact from the degradation of DONS back to DON in the digestive tract was likely minimal in this study. Nevertheless, the degradation pattern is important to consider for future research to potentially enhance the efficacy of the reaction with SMB and lower the dietary concentration of SMB needed to alleviate the effects of DON.
The recovery of DON from pigs fed the NC and NC + Product X matches urinary DON recovery rates in previous work. Since urine is the main DON absorption and excretion route, if Product X was able to decrease the uptake of DON, urinary DON excretion would also be decreased. However, in the present study, DON recovery was similar between pigs fed NC or NC + Product X diets, and the lack of a Product X response in urinary metabolism is congruent with the lack of the growth benefit to Product X. Since pigs have limited ability to de-epoxidate DON other than via microbial fermentation in the hindgut, recovery of urinary DOM-1 was minimal (0.2 to 0.9% of DON intake) but consistent with previous work (0 to 1.1%).
In summary, feeding diets contaminated with 4 ppm DON to nursery pigs reduces their growth most severely during the initial exposure period and is primarily associated with feed intake suppression. The addition of Product X neither alleviated the DON-associated effects on pig growth nor reduced DON absorption and urinary excretion compared to pigs fed DON-contaminated diets alone. However, hydrothermally treating DON-contaminated diets with 1.0% SMB restored feed intake and improved F/G markedly. Even so, the urinary balance experiment revealed that a portion of the converted DONS can be degraded back to DON under physiological conditions. Questions remain surrounding processing methods and long-term supplementation effects of SMB, but this research demonstrates that pelleting DON-contaminated diets with SMB can alleviate DON effects on growth. Additional research is also needed to evaluate the effect of sodium metabisulfite on F/G in noncontaminated diets.
This article was originally published in Kansas Agricultural Experiment Station Research Reports: Vol. 1: Iss. 7. https://doi.org/10.4148/2378-5977.1141. This is an Open Access article licensed under a Creative Commons Attribution 4.0 License.