INTRODUCTION
Zearalenone (ZEA) is produced by Fusarium fungi growing on grains worldwide and is mostly found in its derived products. Concentrations depend on climatic conditions, as well as on the conditions of feed storage and production (Rotter et al., 1996; Voss et al., 2007). With a similar structure to estrogen, zearalenone has been demonstrated to affect swine reproduction (Etienne and Jemmali, 1982; Young et al., 1990). Besides its estrogenic effects, zearalenone has also been shown to be hepatotoxic (Maaroufi et al., 1996), immunotoxic (Abbès et al., 2006), and genotoxic (Abbès et al., 2007) under high dosages in different species. In swine, zearalenone research has mainly focused on breeding performance and the reproductive system (Long and Diekman, 1984) with little information of the effect of ZEA on nutrient availability. In addition, most toxicological data on animals has been obtained using medium to high doses of ZEA (2 to 90 mg/kg of feed). Such high dosages are not commonly found in animal feeds; therefore, one of the objectives of the present study was to investigate the effects of ZEA in pigs under low dosage.
Clay enterosorbent has been used in feeds for counteracting mycotoxicoses in farm animals for several decades (Mayura et al., 1998; Abbès et al., 2007). Although in vitro data have shown that clay enterosorbent could effectively bind ZEA (Sabater-V et al., 2007; Feng et al., 2008), its in vivo effectiveness has not been thoroughly examined. Information on the effects of different levels of natural clay enterosorbent on the growth, nutrient availability, and genital organs of post-weaning female pigs fed low levels of ZEA is only fragmentary and conflicting.
Therefore, the purpose of the present study was to investigate the effects of low ZEA dosage (1 mg/kg) on growth performance, nutrient availability, vulva size, relative weights of genital organs and histological damage of the ovary in post-weaning female pigs; efficacy of a clay enterosorbent in preventing ZEA-induced adverse effects was also evaluated.
MATERIALS AND METHODS
Preparation of zearalenone contaminated diet
Purified crystalline powder (600 mg) of ZEA (Fermenteck, Israel) was dissolved in 600 ml acetic ether and then poured onto 600 g of talcum powder. The spiked material was left overnight to allow evaporation of acetic ether. A mix of 10 mg ZEA /kg premix was prepared by blending 600 g ZEA-contaminated talcum powder with 59.4 kg of ground ZEA-free corn, and was subsequently used at the appropriate levels with a corn-soybean meal diet to make the experimental diets. A composite sample of each experimental diet was sampled for assay of ZEA and other mycotoxins. The content of mycotoxins in the test diets was analyzed by Asia Mycotoxin Analysis Center (Chaoyang University of Technology, Taiwan) before and at the end of the feeding experiment. Actual ZEA contents were 0.1 and 1.3 mg/kg in the control and experimental treatments, respectively. Aflatoxin and fumonisin detected in all treatments were 12.9±1.2 μg/kg and 4.73±0.15 mg/kg, respectively. Deoxynivalenol was not detected in any of the diets.
Experimental design, animals and management
A total of thirty-five post-weaning female piglets (Landrace×Yorkshire×Duroc) with an average body weight of 12.36±1.46 kg were used in the study. Piglets were randomly allocated to 7 experimental groups according to BW after 7 days adaptation. The gilts were fed a corn-soybean meal-whey based diet with an addition of 0 or 1 mg/kg of ZEA for 24 days with four levels of natural clay enterosorbent (Amlan International, United States) added in the feeds. The treatments were: i) control; ii) control+2.5 g/kg clay; iii) control+1 mg/kg ZEA; iv) control+1 mg/kg ZEA+1.25 g/kg clay; v) control+1 mg/kg ZEA+2.5 g/kg clay; vi) control+1 mg/kg ZEA+5.0 g/kg clay; vii) control+ 1 mg/kg ZEA+10 g/kg clay. All gilts were housed individually in metabolism cages in a temperature controlled room at Jinzhuyuan Farm (Yinan, Shandong, China). Animals were provided 0.48 m2 of floor space with one nipple drinker and one hopper type feeder. During the experimental period, the temperature in pigsties was 28 to 26°C. The mean relative humidity was approximately 65%. Animals were cared for in accordance with guidelines of the National Institutes of Health Guide and the China Ministry of Agriculture for the care and use of laboratory animals.
All diets (Table 1) used in the study were isocaloric and isonitrogenous with only treatment difference in ZEA and clay enterosorbent. Nutrient concentrations met or exceeded minimal requirements according to the NRC (1998). Representative feed samples were taken at the beginning and end of feeding for nutrient and mycotoxin analyses. Gross energy (GE), crude protein (CP), calcium, phosphorus, crude fiber, ether extract, crude ash and amino acid contents of the diets were analyzed according to the AOAC (1990).
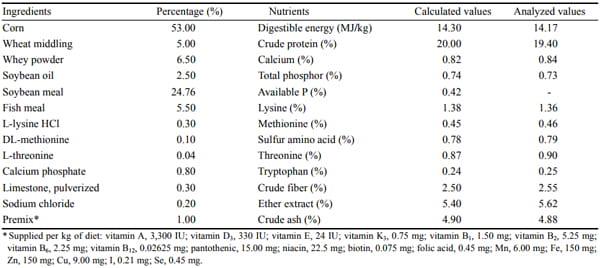
Table 1. Ingredients and composition of the basal diet
Body weights were measured weekly and at the end of the test. Feed intakes and refusals of the piglets were recorded daily. Average daily gain (ADG), average daily feed intake (ADFI) and feed efficiency (FE, g gain/g feed) were calculated based on these data.
Sample collections for analyses of nutrient availability
Three gilts from each treatment were randomly selected for determination of nutrient digestibility. Total quantity of feces and urine excreted by each pig was collected daily. Feces and urine were weighed, thoroughly mixed separately, and then sampled daily. Total fecal and urine samples were pooled by pig and analyzed on a weekly basis. Representative samples were stored at -20°C until analysis. For crude protein analysis, 2.5% of daily feces and 1% of daily urine excreted were preserved in 1:3 diluted sulfuric acid (250 ml concentrated sulfuric acid per liter solution). Subsamples of feeds and feces were dried in a 60°C oven for 48 h and the dried samples were finely ground by mortar and pestle to pass through a 1 mm screen and then stored in sealed containers until analysis of gross energy and nitrogen content. Feed intakes were recorded daily. Digestibility of crude protein (DCP) and net protein utilization (NPU) were calculated as DCP = (DN/IN)×100 and NPU = (RN/IN)× 100, respectively, where DN (digested N) = ingested Nfecal N, RN (retained N) = ingested N-fecal N-urinary N. Digestibility of gross energy (DGE) and metabolic rate of gross energy (MRGE) were calculated as DGE = (DE/GE) ×100 and MRGE = (ME/GE)×100, respectively, where DE = GE-fecal energy, ME = GE-fecal energy-urinary energy.
Vulva and organ weights
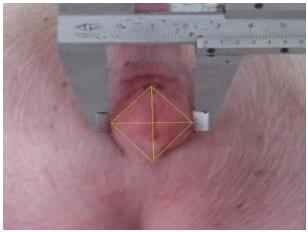
Vulva length and width were measured at 4 d intervals, and vulva area was calculated approximately as a diamond shape ((vulva length×vulva width)/2) as shown in Figure 1.
Figure 1. Measurement and calculation of vulva size.
Upon completion of the experiment, the piglets were killed by euthanasia, necropsied and examined for gross lesions of the reproductive organs. The genital organs (ovary+cornu uteri+vagina-vestibule) were isolated and weighed. The organ weights were expressed on a relative body weight basis (g/kg).
Pathohistological tests
Ovaries were extirpated and fixed promptly in 10% buffered formalin upon weighing. After routine processing, the tissues were embedded in paraffin. For general orientation, the tissues were sectioned in pieces of 5 μm thickness and stained with hematoxylin and eosin (H&E) for microscopic examination. The slides were examined under 100× or 200× magnification using an optical microscope (Carl Zeiss, Germany).
Statistical analysis
All data were subjected to analysis of variance using the GLM procedure of SAS (SAS 9.1, 2003). The data were first analyzed as a completely randomized design with individual piglet as random factor to examine the overall effect of treatments. The efficacy of supplemental clay was determined by using a contrast between ZEA-contaminated diets and ZEA-contaminated diets with clay enterosorbent. Orthogonal polynomial contrasts were then used to determine linear and quadratic responses to clay levels with the addition of purified ZEA treatments. The significance of differences among treatments was tested using Duncan’s multiple range test. All statements of significance were based on the probability of p<0.05.
RESULTS
Growth performance
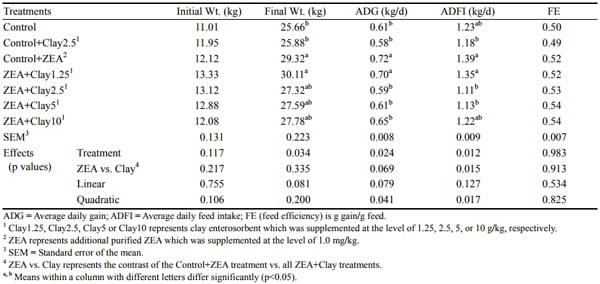
There were no differences (p>0.05) in final weight, ADG, ADFI, and FE as compared between pigs fed control and control+2.5 g/kg clay diets (Table 2); implying no negative interaction of feeding the clay enterosorbent on normal pig growth after 24 days feeding. Pigs fed a diet containing 1 mg/kg purified ZEA showed increases (p< 0.05) in final weight, ADG, and ADFI compared with pigs fed the control diet, indicating that increased weight gain was mainly due to increased feed consumption. Final weight, ADG, ADFI, and FE of pigs fed the ZEAcontaminated diet or the ZEA-contaminated diet with addition of 1.25 g/kg clay were not different (p>0.05). Although no difference was obtained in final weights of pigs fed ZEA diets with different levels of clay enterosorbent supplement, the ADG and ADFI were significantly different (p<0.05) when higher inclusion rate of clay enterosorbent was used in the diet. No difference (p>0.05) of FE was obtained among treatments suggesting that increased weight gain in some treatments was due to increased feed intake. Addition of clay enterosorbent at increasing levels to the ZEA-contaminated diet had a quadratic effect (p<0.05) on reduced ADG and ADFI as compared with the single ZEA-contaminated treatment.
Table 2. Growth performance of pigs fed zearalenone contaminated feeds with or without clay enterosorbent for 24 days
Nutrient availability
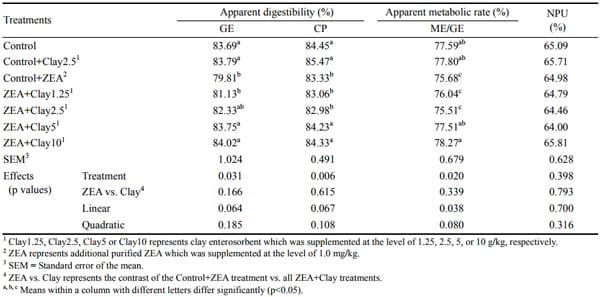
Pigs fed control+2.5 g/kg clay showed no differences in apparent digestibility of nutrients (CP, GE), metabolic rate of GE (ME/GE) and net protein utilization (NPU) as compared to pigs fed the control diet (p>0.05, Table 3). However, pigs fed the control+ZEA diet had significant reductions in apparent digestibility of nutrients (CP, GE) and ME/GE as compared to pigs fed the control diet (p< 0.05). Pigs fed the ZEA+1.25 g/kg clay diet showed similar nutrient digestibility as compared to pigs fed the control+ ZEA diet; indicating little effect of low level of dietary clay enterosorbent under 1 mg/kg of purified ZEA challenge. However, the reduced apparent digestibility of nutrients (CP, GE) and ME/GE were significantly improved (p<0.05) when diets were supplemented with 5 or 10 g/kg of the clay enterosorbent compared with control+ZEA treatment. Dietary addition of clay enterosorbent in the ZEAcontaminated diet linearly (p<0.10) improved the apparent digestibility of nutrients (CP, GE) and ME/GE as the clay concentration increased. Net protein utilization was not different among treatments (p>0.05).
Table 3. Apparent digestibility of nutrients (CP, GE), metabolic rate of GE (ME/GE) and net protein utilization (NPU) of pigs fed zearalenone contaminated feeds with or without clay enterosorbent for 24 days (CP on wet-basis and GE on dry basis)
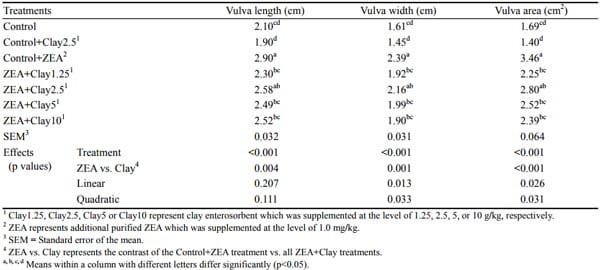
Table 4. Vulva size of pigs fed zearalenone contaminated feeds with or without clay enterosorbent for 24 days
Vulva size and organ weight
Gilts fed diets containing 1.3 mg/kg ZEA without clay had significantly increased vulva size as compared to other treatments except for the ZEA+2.5 g/kg clay treatment (p< 0.001, Table 4). Simultaneous addition of a higher level of clay enterosorbent at 5 and 10 g/kg levels resulted in a significant reduction in vulva length, vulva width and vulva area as compared to the control+ZEA treatment (p<0.05). Pigs which received the control+2.5 g/kg clay diet had the smallest vulva length, width, and area although the values were not different to the control group. Addition of clay enterosorbent at increasing levels to the ZEA-contaminated diet quadratically reduced (p<0.05) the enlarged vulva width and vulva area when pigs were fed diets contaminated with purified 1 mg/kg ZEA.
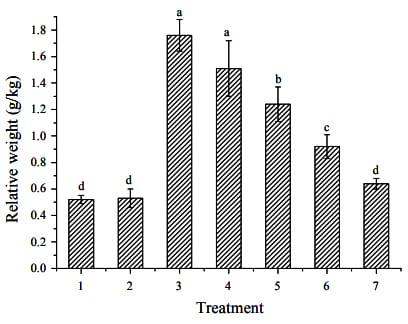
Gilts fed the test diet (control+ZEA) had significantly increased relative weight of reproductive organs as compared to the control (p<0.001) (Figure 2). Relative weight of genital organs in pigs fed ZEA contaminated feeds decreased linearly as dietary natural clay enterosorbent increased (p<0.05).
Histopathological examination
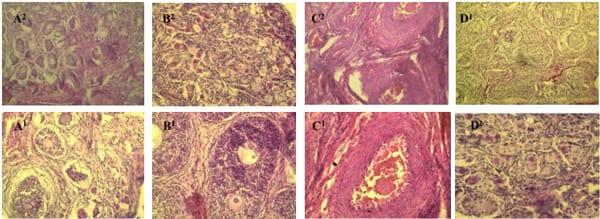
No macroscopic pathological changes of ovaries among treatments were observed by the veterinarian through clinical examination. Ovarian sections from piglets treated with ZEA alone showed numerous primordial follicles in the cortex (B1), hyperplasia of vessel wall in the medulla and hyperemia in blood vessels (C1), two primary oocyte primary oocytes in one primary follicle (B2), numerous erythrocytes and leukocytes filled with lumina (C2) (Figure 3). However, moderate primordial follicles in different stages of development in the cortex of the ovarian sections were found in ZEA and clay (10 g/kg) groups, and only individual primordial follicles developed to primary follicles (D1 and D2). Normal histological pictures were observed in control groups (A1 and A2).
DISCUSSION
In the present study, ADG and ADFI of pigs fed the control+ZEA diet over 24 days were increased by 17.2% and 13.3%, respectively, compared with pigs fed the control diet. These findings are contrary to reports of pigs fed with increasing dietary levels of ZEA (Kalliamurthy et al., 1997). Šperanda et al. (2006) reported that weaned pigs fed diets containing addition of 3 mg/kg ZEA showed lower body weights, but had no statistical significance. Other studies found no effect of ZEA on growth performance of gilts (Etienne et al., 1982; James et al., 1982; Rainey et al., 1990). However, Pfaffl et al. (2001) reported ZEA as a strong anabolic compound in a cell culture model which supports the present results. We previously conducted a dose response feeding trial (data presented in ASAS annual meeting, 2008) adding 0, 1, 2, and 3 mg/kg of purified ZEA to a similar basal diet, and fed the test diets to female pigs of similar age. Results showed that liver, kidney, and reproductive organ weights were significantly increased as dietary ZEA increased; however, dressing percentage was not different between treatments. Therefore, the increased ADG in the present study may be partially due to increased target organ weights.
Figure 2. Effects of zearalenone and different levels of clay on relative weight of genital organ of piglets. i) control; ii) control+2.5 g/kg clay; iii) control+1 mg/kg ZEA; iv) control+1 mg/kg ZEA+1.25 g/kg clay; v) control+1 mg/kg ZEA+2.5 g/kg clay; vi) control+1 mg/kg ZEA+5.0 g/kg clay; vii) control+1 mg/kg ZEA+10 g/kg clay.
Figure 3. Photomicrographs of hematoxylin and eosin stained ovary sections of piglets after feeding ZEA contaminated feeds for 24 days. Bars equal 5 μm (1 100×original magnification or 2 200×original magnification). A = Control; B, C = Control+1 mg/kg ZEA; D = control+1 mg/kg ZEA+10 g/kg clay.
Clay enterosorbent has been recognized as a binder for mycotoxins in feeds for different species; however, controversial results were reported from different studies. For instance, Papaiounnou et al. (2002) reported that organozeolites were beneficial for sows, gilts and the performance of their litters. Positive effects of clinoptilolite on growing swine performance were also found by Ward et al. (1991) and Pond et al. (1988). On the other hand, in the present study, we demonstrated no improvement in growth performance on adding clay enterosorbent, although adding clay enterosorbent into ZEA contaminated feed showed, numerically, improvement in feed efficiency as compared to the control.
Only limited data are available regarding the effect of ZEA on nutrient digestibility. Hauschild et al. (2007) reported that feeding 2 mg/kg ZEA to pigs did not affect digestibility of dry matter and gross energy, metabolizable energy, digestible energy as well as digestible protein, and addition of 0.3% organo-aluminosilicate in the ZEAcontaminated feed had no positive or negative impact on nutrient digestibility. However, our previous study concluded that the apparent dry matter and organic matter digestibility of nutrient substance (CP, GE) and NPU were all decreased linearly as dietary ZEA concentrations increased (0, 1, 2, or 3 mg/kg purified ZEA in diets) (unpublished data), which was consistent with the present results. However, Fokas et al. (2004) determined no effect of organozeolites on nutrient utilization in growing pigs. Aflatoxin and fumonisin were detected in the basal diet used in the present study; the interaction of multiple mycotoxins may result in reductions of apparent digestibility of nutrients (CP, GE) and ME/GE. Further studies will be needed to elucidate the potential interaction effects of multiple mycotoxins. Nevertheless, the present study showed that gilts fed ZEA-contaminated diets supplemented with clay enterosorbent had ameliorated effects on apparent digestibility of nutrients (CP, GE) and ME/GE.
ZEA induces hyperestrogenosis in pigs; typical clinical symptoms are swollen vulva, prolapse of the vagina and rectum, and enlargement of the mammary glands (Stob et al., 1962). After 7 days of feeding in the study (data not shown), a swollen vulva was visually observed in those female pigs fed the control+ZEA diet, and at 24 days the pigs showed a 38.1%, 48.4% and 104.7% increase in vulva length, vulva width, and vulva area, respectively, as compared to the control group. The detrimental effects on vulva swelling by ZEA feeding in the present study were linearly decreased by the addition of clay enterosorbent. This implies that clay enterosorbent at the levels of 5 or 10 g/kg could effectively spare the estrogenic effect of ZEA on vulva swelling in post-weaning female pigs. Papaioannou et al. (2002) reported that reproductive performance of sows was improved by the inclusion of zeolite in ZEAcontaminated diets (0.16 to 1.55 mg/kg). A previous study revealed that continuous exposure of 0.25 mg/kg dietary ZEA in young pigs resulted in a hyper-estrogenism (Cheng et al., 2006). Etienne et al. (1982) reported that uterus weight was nearly doubled after gilts ingested 3.61 and 4.33 mg/kg ZEA. Vulva vaginitis and enlarged reproductive tracts were observed in all female pigs after feeding zearalenone orally in gelatine capsules at 3.5, 7.5 or 11.5 mg/kg bodyweight (Farnworth and Trenholm, 1981). Vulva swelling and reddening were seen within seven days of ZEA (1.5 mg/kg or 2 mg/kg) exposure (Rainey et al., 1990). In the present study, gilts fed ZEA-contaminated diets increased both vulva size and ovary-uterus weights, and supplementation with clay enterosorbent showed positive effects on the reduction of vulva size and relative weight of genital organs, which was confirmed by changes in histopathology of the ovary.
The results of histopathological changes in the ZEAtreated piglets suggested that the ovaries were inflamed and hyperestrogenized by dietary ZEA, which was supported by the hyperplasia of vessel wall in the medulla and hyperemia in blood vessels of the ovary, numerous erythrocytes and leukocytes filled with lumina, two primary oocytes in one primary follicle, and numerous primordial follicles in the cortex. Diekman and Long (1989) found that blastocyst development was influenced in ZEA-treated sows that received contaminated feed 7 days after breeding. Previous studies showed that ZEA had proliferation effects on uteral cells in gilts (Obremski et al., 2003; Zwierzchowski et al., 2005). Moreover, Wasowicz et al. (2005) reported that ZEA (20 or 40 μg/kg bodyweight) did not induce apoptosis in porcine ovaries. The present study showed a clear protective effect of the clay enterosorbent against ovarian tissue changes induced by dietary ZEA. The findings suggested that clay enterosorbent may sequester ZEA in the digestive tract which lead to decreased ZEA absorption in the small intestine.
IMPLICATIONS
The study demonstrated that young gilts which received 1 mg/kg purified ZEA for 24 days had reduced apparent digestibility of nutrients (CP, GE) and ME/GE, and increased vulva size and reproductive organ weights as compared to pigs fed the control diet. Addition of natural clay enterosorbent to ZEA contaminated feeds showed amelioration effects on nutrient digestibility, vulva size, genital organ weights, and histopathological changes of ovaries, and the protection increased linearly as clay enterosorbent supplement increased. This implies that the clay enterosorbent used in the study may apply in the field for protection against the detrimental effects of ZEA in swine feeding.
This article was originally published in Asian-Australasian Journal of Animal Sciences 2010;23(1): 74-81. DOI: https://doi.org/10.5713/ajas.2010.90242. This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) REFERENCES
Abbès, S., Z. Ouanes, J. B. Salah-Abbes, M. A. Abdel-Wahhab, R. Oueslati and H. Bacha. 2007. Preventive roles of aluminosilicate clay against induction of micronuclei and chromosome aberrations in bone-marrow cells of Balb/c mice treated with Zearalenone. Mutat. Res. 631:85-92.
Abbès, S., Z. Ouanes, J. B. Salah-Abbès, Z. Houas, R. Oueslati, H. Bacha and O. Othman. 2006. The protective effect of hydrated sodium calcium aluminosilicate against haematological, biochemical and pathological changes induced by Zearalenone in mice. Toxicon. 47:567-574.
AOAC. 1990. Official methods of analysis. 15th edn. Association of Official Analytical Chemists, Arlington, Virginia.
Cheng, Y. H., C. F. Weng, B. J. Chen and M. H. Chang. 2006. Toxicity of different Fusarium mycotoxins on growth performance, immune responses and efficacy of a mycotoxin degrading enzyme in pigs. Anim. Res. 55:579-590.
Diekman, M. A. and G. G. Long. 1989. Blastocyst development on days 10 or 14 after consumption of zearalenone by sows on days 7 to 10 after breeding. Am. J. Vet. Res. 50:1224-1227.
Etienne, M. and M. Jemmali. 1982. Effects of Zearalenone (F2) on Estrous Activity and Reproduction in gilts. J. Anim. Sci. 55:1- 10.
Farnworth, E. R. and H. L. Trenholm. 1981. The effect of acute administration of the mycotoxin zearalenone to female pigs. J. Environ. Sci. Health B, 16:239-252.
Feng, J., M. Shan, H. Du, X. Han and Z. Xu. 2008. In vitro adsorption of zearalenone by cetyltrimethyl ammonium bromide-natural montmorillonite nanocomposites. Micropor. Mesopor. Mat. 113:99-105.
Fokas, P., G. Zervas, K. Fegeros and P. Zoipoulos. 2004. Assessment of Pb retention coefficient and nutrient utilization in growing pigs diets with added clinoptilolite. Anim. Feed Sci. Technol. 117:121-129.
Hauschild, L., P. A. Lovatto, C. R. Lehnen, A. Á. Carvalho, G. G. Garciae and C. A. Mallmann. 2007. Digestibility and metabolism of piglet diets containing zearalenone with addition of organoaluminosilicate. Pesqui. Agropecu. Bras. 42:219-224.
James, L. J. and T. K. Smith. 1982. Effect of dietary alfalfa on zearalenone toxicity and metabolism in rats and swine. J. Anim. Sci. 55:110-118.
Kalliamurthy, J., P. Geraldine and P. A. Thomas. 1997. Effects of Zearalenone on food consumption, growing rate, organ weight and serum testosterone level in male rats. J. Environ. Biol. 18:115-120.
Long, G. G. and M. A. Diekman. 1984. Effect of purified Zearalenone on early gestation in gilts. J. Anim. Sci. 59:1662- 1670.
Maaroufi, K., L. Chekir, E. E. Creppy, F. Ellouz and H. Bacha. 1996. Zearalenone induces modifications of haematological and biochemical parameters in rats. Toxicon. 34:535-540.
Mayura, K., M. A. Abdel-Wahhab, K. S. Mckenzie, A. B. Sarr, J. F. Edwards, K. Naguib and T. D. Philips. 1998. Prevention of maternal and developmental toxicity in rats via dietary inclusion of common aflatoxin sorbents: potential for hidden risks. Toxicol. Sci. 41:165-167.
National Research Council. 1998. Nutrient Requirements of Swine. 10th Ed. National Academy Press, Washington, DC.
Obremski, K., M. Gajecki, W. Zwierzchowski, L. Zielonka, I. Otrocka-Domagala, T. Rotkiewicz, A. Mikolajczyk, M. Gajecka and M. Polak. 2003. Influence of zearalenone on reproductive system cell proliferation in gilts. Pol. J. Vet. Sci. 6:239-245.
Papaioannou, D. S., S. C. Kyriakis, A. Papasteriadis, N. Roumbies, A. Yannakopoulos and C. Alexopoulos. 2002. A field study on the effect of in-feed inclusion of a natural zeolite (clinoptilolite) on health status and performance of sows/gilts and their litters. Res. Vet. Sci. 72:51-59.
Pfaffl, M. W., I. G. Lange, A. Daxenberger and H. H. D. Meyer. 2001. Tissue-specific expression pattern of estrogen receptors (ER): Quantification of ERá and ERâ mRNA with real-time RT-PCR. APMIS. 109:345-355.
Pond, W. G., J. T. Yen and V. H.Varel. 1988. Response of growing swine to dietary copper and clinoptilolite supplementation. Nutr. Rep. Int. 37:797-803.
Rotter, B. A., D. B. Prelusky and J. J. Pestka. 1996. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health. 48:1- 34.
Rainey, M. R., R. C. Tubbs, L. W. Bennet and N. M. Cox. 1990. Prepubertal exposure to dietary zearalenone alters hypothalamo-hypophyseal function but does not impair postpubertal reproductive functions in gilts. J. Anim. Sci. 68:2015-2022.
Sabater-Vilar1, M., H. Malekinejad, M. H. J. Selman1, M. A. M. Doelen and J. Fink-Gremmel. 2007. In vitro assessment of adsorbents aiming to prevent deoxynivalenol and zearalenone mycotoxicoses. Mycopathologia 163:81-90.
SAS Institute. 2003. SAS/STAT User's Guide: Version 9.1th edn. SAS Institute Inc., Cary, North Carolina.
Šperanda, M., B. Liker, T. Šperanda, V. Šeric, Z. Antunovic, Ž. Grabarevic, D. Sencic and Z. Steiner. 2006. Haematological and biochemical parameters of weaned piglets fed on fodder mixture contaminated by zearalenone with addition of clinoptilolite. Acta. Vet. (Beogr.) 56:121-136.
Stob, M., R. S. Baldwin, J. Tuite, F. H. Andrews and K. G. Gillette. 1962. Isolation of an anabolic, uterotrophic compound from corn infected with Gibberella Zeae. Nature 196:1318.
Voss, K. A. and W. M. Haschek. 2007. Fumonisins: toxicokinetics, mechanism of action and toxicity. In: Fusarium and their toxins: Mycology, occurrence, toxicity, control and economic impact (D. P. Morgavi and R. T. Riley). Anim. Feed Sci. Technol. 137:299-325.
Ward, T. L., K. L. Watkins, L. L.Southern, P. G. Hoyl and D. D. French. 1991. Interactive effects of sodium zeolite-A and copper in growing swine: growth, and bone and tissue mineral concentrations. J. Anim. Sci. 69:726-733.
Wasowicz, K., M. Gajecka, J. Calka, E. Jakimiuk and M. Gajecki. 2005. Influence of chronic administration of zearalenone on the processes of apoptosis in the porcine ovary. Vet. Med. (Czech) 50:531-536.
Yang, Z. B., H. Zao, C. C. Chen and F. Chi. 2008. Feeding different levels of zearalenone on growth, vulva size, and organ weight in postweanling female pig. Proc. 2008. Joint Meeting. J. Anim. Sci. 86(Suppl. 2):45(Abstr.).
Young, L. G., H. Ping and G. J. King. 1990. Effects of feeding zearalenone to sows on rebreeding and pregnancy. J. Anim. Sci. 68:15-20.
Zwierzchowski, W., M. Przybylowicz, K. Obremski, L. Zielonka, E. Skorska-Wyszynska, M. Gajecka, M. Polak, E. Jakimiuk, B. Jana, L. Rybarczyk and M. Gajecki. 2005. Level of zearalenone in blood serum and lesions in ovarian follicles of sexually immature gilts in the course of zearalenone mycotoxicosis. Pol. J. Vet. Sci. 8:209-218.







.jpg&w=3840&q=75)