Counteracting mycotoxin contamination
Counteracting mycotoxin contamination: the effectiveness of Saccharomyces cerevisiae cell wall glucans in Mycosorb® for sequestering mycotoxins
Published: June 21, 2007
Source : Alexandros Yiannikouris (Alltech), Jean-Pierre Jouany (INRA) and Bertin Gérard(Courtesy of Alltech Inc.)
Mycotoxins are secondary metabolites secreted by moulds, mostly belonging to the three genera Aspergillus, Penicillium and Fusarium produced in cereal grains as well as forages before, during and after harvest.
Forages and cereals naturally come into contact with fungal spores. The fungal contamination of plants and the biosynthesis of toxins depend on the state of health of the plant before harvest, meteorological conditions, harvesting techniques, delays and hydrothermal conditions before stabilisation for conservation and feed processing.
Depending on the fungus, fungal growth is then controlled by a number of physico-chemical parameters including the amount of free water (aw), temperature, presence of oxygen, nature of the substrate, and pH conditions.
Mycotoxins could be then formed pre-harvest as well as post-harvest in storage. Rodents, birds and insects may facilitate contamination by causing physical lesions on plants, providing a route of entry into the plant for fungal spores (CAST, 2003). Thus, mycotoxins could be present throughout food and feeds derived from plants and especially in processed feeds.
In this respect, distillers dry grains with solubles (DDGS) represent an important potential source of mycotoxins since mycotoxins are able to survive the ethanol production process and are concentrated three-fold in the residual matrix. Avoiding mycotoxin occurrence in the food chain involves a clear understanding of mycotoxin management strategies and effective quality control of food and feeds through adequate sampling, detection and quantification methodology.
Nevertheless, it is difficult to estimate precisely the mycotoxin concentration in a large bulk lot because of their uneven distribution in commodities and the large variability associated with the overall mycotoxin isolation procedure. Even when using accepted test procedures, there is variability associated with the selection of the sample, the grinding/mixing/ homogenization procedure and the extraction method used.
As an example, the distillers dried grains with solubles (DDGS) matrix gave strong background interference contributing variability by binding and/or masking the real mycotoxin level. Because of this variability, the true mycotoxin concentration in feeds cannot be determined with 100% certainty. Furthermore, significant total amounts may be present in feed at low concentrations.
Fungal contamination affects both the organoleptic characteristics and the alimentary value of feed, and entails a risk of toxicosis. The biological effects of mycotoxins depend on the ingested amounts, number of toxins, duration of exposure and animal sensitivity.
Mycotoxins can induce health troubles that are specific to each toxin, have synergistic toxic properties in the frequent case of multi-contamination, and/or affect the immune status of animals promoting infections and have a negative impact on livestock production. In addition, the possible presence of toxic residues in edible animal products when animals are fed contaminated feeds may have some detrimental effects on human health.
The control of mould growth involves maintaining the physical integrity of cereal grains with the aim of limiting the access of moulds to nutrients present in grains, and the strict control of environmental conditions. Chemical, physical and biological methods could be applied to limit both mould contamination and to decrease the concentration of mycotoxins present in food and feeds.
However, mycotoxin risk remains unavoidable and the lack of practical solutions to totally avoid mycotoxin contamination of feed explains why inorganic materials such as clays, bentonites and aluminosilicates, known for their adsorptive properties, were first proposed to reduce the toxic effect of aflatoxins (Grant and Phillips, 1998). However, these inorganic materials showed only limited efficacy against other mycotoxins. Furthermore, they reduced the biological value of certain nutrients and could contain dioxins and heavy metals.
Organic materials such as yeast cell walls (Devegowda et al., 1998) have been proposed as an alternative solution to complex several mycotoxins within the gastrointestinal tract without impairing nutrient bioavailability or inducing detrimental environmental effects.
It is important to understand the chemistry of the sequestration process involved in toxin clearance. More than simple ‘binding assays’ (Yiannikouris et al., 2003), interaction kinetic models based on overall capacity, standardized affinity rate and stereochemical views are becoming available. These models are required to complete and support in a meaningful way the advances in applied nutrition needed to define the beneficial role of organic sequestrants prepared from yeast cell wall.
Investigations
The chemical mechanisms involved in the sequestering activity of Saccharomyces cerevisiae cell wall components with several major mycotoxins were investigated. The in vitro methodology was based on the comparison of several sources of yeast cell wall (i.e., wt292, fks1, sc1026, mnn9) differing in their relative glucan/mannan/chitin content.
The evaluation of the affinity rate was achieved using Hill’s model given the equations in Table 1. Equation (1) was used to establish two sub-models: (i) Equation (2) took into account the amounts of total ß-D-glucans in the cell wall related to the sequestration properties for the mycotoxin (Yiannikouris et al., 2004a); (ii) Equation (3) took into account the respective roles and amounts of extracted fractions of the cell wall in the sequestration process (Yiannikouris et al., 2004b).
Each equation led to the calculation of the affinity rates A (Equation 4). ß-D-glucans, composed of linear chains of ß-(1 3)-D-glucans branched with ß-(16)-D-glucan side chains, are clearly important in defining the efficacy of cell wall materials against aflatoxin B1 (AFB1), deoxynivalenol (DON), zearalenone (ZEA), patulin (PAT) and ochratoxin A (OTA). A correlation between the ß-D-glucan amount and the efficacy of sequestering ZEA, AFB1, DON, PAT and, to a lesser extent OTA, was demonstrated (R² > 0.889; RSD < 0.855 μg/mL, Table 2).
3)-D-glucans branched with ß-(16)-D-glucan side chains, are clearly important in defining the efficacy of cell wall materials against aflatoxin B1 (AFB1), deoxynivalenol (DON), zearalenone (ZEA), patulin (PAT) and ochratoxin A (OTA). A correlation between the ß-D-glucan amount and the efficacy of sequestering ZEA, AFB1, DON, PAT and, to a lesser extent OTA, was demonstrated (R² > 0.889; RSD < 0.855 μg/mL, Table 2).
For all of these mycotoxins, cell walls of wt292 and mnn9 yeast strains with higher levels of ß-D-glucans are able to complex larger amounts of toxins with higher affinity rates than the fks1 and sc1026 strains. The higher chitin content in mnn9 and fks1 strains results in higher insolubility of ß-D-glucans, less flexibility of the overall structure and, therefore, a decrease in the access of mycotoxins to the chemical sites of the ß-D-glucans.
Thus, these strains exhibit a lower complex-forming capacity than expected from their ß-D-glucan contents (Yiannikouris et al., 2004a). Differences between mycotoxins were also recorded and could be undoubtedly assigned to the stereochemical, electrical charge, solubility, nature and size discrepancies between mycotoxins (Figure 1). Their affinities for the 3D-structure of the binding sites offered by ß-D-glucans are consequently variable and ranked from 95.0 to 12.2% affinity as follows: AFB1 > DON > ZEA > PAT > OTA.
Another study showed significant efficacy for sequestering T-2 toxin, as well as endophyte-associated toxins. The alkali extraction of the glucan fraction from the entire yeast cell wall contributed to increasing the potential of glucans to sequester mycotoxins.
Differences were then characterized in relation to the structure of the glucan fraction extracted. For example, the alkali-insoluble fraction had a greater affinity for ZEA sequestration (up to 50%) than the alkali-soluble fraction (about 16%). From the results obtained with the individual fractions and plotting our data with the Equation (3) (R² = 0.969; RSD = 0.296 μg/mL, Figure 2), the structure of glucans available in mnn9 and wt292 strains had the highest ZEA sequestration efficacy. Elimination of a part of the chitin content during alkali extraction of yeast cell wall improved the sequestrant activity 150% (Yiannikouris et al., 2004b).
Forages and cereals naturally come into contact with fungal spores. The fungal contamination of plants and the biosynthesis of toxins depend on the state of health of the plant before harvest, meteorological conditions, harvesting techniques, delays and hydrothermal conditions before stabilisation for conservation and feed processing.
Depending on the fungus, fungal growth is then controlled by a number of physico-chemical parameters including the amount of free water (aw), temperature, presence of oxygen, nature of the substrate, and pH conditions.
Mycotoxins could be then formed pre-harvest as well as post-harvest in storage. Rodents, birds and insects may facilitate contamination by causing physical lesions on plants, providing a route of entry into the plant for fungal spores (CAST, 2003). Thus, mycotoxins could be present throughout food and feeds derived from plants and especially in processed feeds.
In this respect, distillers dry grains with solubles (DDGS) represent an important potential source of mycotoxins since mycotoxins are able to survive the ethanol production process and are concentrated three-fold in the residual matrix. Avoiding mycotoxin occurrence in the food chain involves a clear understanding of mycotoxin management strategies and effective quality control of food and feeds through adequate sampling, detection and quantification methodology.
Nevertheless, it is difficult to estimate precisely the mycotoxin concentration in a large bulk lot because of their uneven distribution in commodities and the large variability associated with the overall mycotoxin isolation procedure. Even when using accepted test procedures, there is variability associated with the selection of the sample, the grinding/mixing/ homogenization procedure and the extraction method used.
As an example, the distillers dried grains with solubles (DDGS) matrix gave strong background interference contributing variability by binding and/or masking the real mycotoxin level. Because of this variability, the true mycotoxin concentration in feeds cannot be determined with 100% certainty. Furthermore, significant total amounts may be present in feed at low concentrations.
Fungal contamination affects both the organoleptic characteristics and the alimentary value of feed, and entails a risk of toxicosis. The biological effects of mycotoxins depend on the ingested amounts, number of toxins, duration of exposure and animal sensitivity.
Mycotoxins can induce health troubles that are specific to each toxin, have synergistic toxic properties in the frequent case of multi-contamination, and/or affect the immune status of animals promoting infections and have a negative impact on livestock production. In addition, the possible presence of toxic residues in edible animal products when animals are fed contaminated feeds may have some detrimental effects on human health.
The control of mould growth involves maintaining the physical integrity of cereal grains with the aim of limiting the access of moulds to nutrients present in grains, and the strict control of environmental conditions. Chemical, physical and biological methods could be applied to limit both mould contamination and to decrease the concentration of mycotoxins present in food and feeds.
However, mycotoxin risk remains unavoidable and the lack of practical solutions to totally avoid mycotoxin contamination of feed explains why inorganic materials such as clays, bentonites and aluminosilicates, known for their adsorptive properties, were first proposed to reduce the toxic effect of aflatoxins (Grant and Phillips, 1998). However, these inorganic materials showed only limited efficacy against other mycotoxins. Furthermore, they reduced the biological value of certain nutrients and could contain dioxins and heavy metals.
Organic materials such as yeast cell walls (Devegowda et al., 1998) have been proposed as an alternative solution to complex several mycotoxins within the gastrointestinal tract without impairing nutrient bioavailability or inducing detrimental environmental effects.
It is important to understand the chemistry of the sequestration process involved in toxin clearance. More than simple ‘binding assays’ (Yiannikouris et al., 2003), interaction kinetic models based on overall capacity, standardized affinity rate and stereochemical views are becoming available. These models are required to complete and support in a meaningful way the advances in applied nutrition needed to define the beneficial role of organic sequestrants prepared from yeast cell wall.
Investigations
The chemical mechanisms involved in the sequestering activity of Saccharomyces cerevisiae cell wall components with several major mycotoxins were investigated. The in vitro methodology was based on the comparison of several sources of yeast cell wall (i.e., wt292, fks1, sc1026, mnn9) differing in their relative glucan/mannan/chitin content.
The evaluation of the affinity rate was achieved using Hill’s model given the equations in Table 1. Equation (1) was used to establish two sub-models: (i) Equation (2) took into account the amounts of total ß-D-glucans in the cell wall related to the sequestration properties for the mycotoxin (Yiannikouris et al., 2004a); (ii) Equation (3) took into account the respective roles and amounts of extracted fractions of the cell wall in the sequestration process (Yiannikouris et al., 2004b).
Each equation led to the calculation of the affinity rates A (Equation 4). ß-D-glucans, composed of linear chains of ß-(1
For all of these mycotoxins, cell walls of wt292 and mnn9 yeast strains with higher levels of ß-D-glucans are able to complex larger amounts of toxins with higher affinity rates than the fks1 and sc1026 strains. The higher chitin content in mnn9 and fks1 strains results in higher insolubility of ß-D-glucans, less flexibility of the overall structure and, therefore, a decrease in the access of mycotoxins to the chemical sites of the ß-D-glucans.
Thus, these strains exhibit a lower complex-forming capacity than expected from their ß-D-glucan contents (Yiannikouris et al., 2004a). Differences between mycotoxins were also recorded and could be undoubtedly assigned to the stereochemical, electrical charge, solubility, nature and size discrepancies between mycotoxins (Figure 1). Their affinities for the 3D-structure of the binding sites offered by ß-D-glucans are consequently variable and ranked from 95.0 to 12.2% affinity as follows: AFB1 > DON > ZEA > PAT > OTA.
Another study showed significant efficacy for sequestering T-2 toxin, as well as endophyte-associated toxins. The alkali extraction of the glucan fraction from the entire yeast cell wall contributed to increasing the potential of glucans to sequester mycotoxins.
Differences were then characterized in relation to the structure of the glucan fraction extracted. For example, the alkali-insoluble fraction had a greater affinity for ZEA sequestration (up to 50%) than the alkali-soluble fraction (about 16%). From the results obtained with the individual fractions and plotting our data with the Equation (3) (R² = 0.969; RSD = 0.296 μg/mL, Figure 2), the structure of glucans available in mnn9 and wt292 strains had the highest ZEA sequestration efficacy. Elimination of a part of the chitin content during alkali extraction of yeast cell wall improved the sequestrant activity 150% (Yiannikouris et al., 2004b).
Table 1. Models used to plot and evaluate toxin sequestration on S. cerevisiae cell wall ß-D-glucans according to Hill’s equation. |
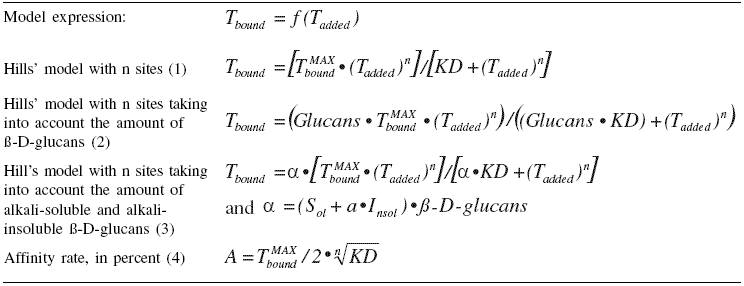 to enlarge the image, click here |
| a a = variable distinguishing the role played by alkali-insoluble compared with alkali-soluble ß-D-glucan; Insol = amount of alkali-insoluble ß-D-glucan (%); Sol = amount of alkali-soluble ß-D-glucans (%); KD = association constant compared with amount of toxins initially added; Tbound = amount of bound toxin (μg/mL) ; Tadded = amount of toxin added (μg/mL); T maxbound = maximal amount of bound toxin (μg/mL). |
The affirmation of the chemical interaction between mycotoxins and sequestrant was demonstrated using a molecular mechanics investigation, because the affinity was not only dependent on the quantity of ß-D-glucan present in the cell wall but also on ß-D-glucan structure and the network organization defined in the single and/or triple helix structure.
| Table 2. Evaluation of the biological parameters for the sequestration of AFB1, PAT, DON and OTA using equation (3) integrating the amount of soluble and insoluble alkali-fractions of ß-D-glucans extracted from four strains of S. cerevisiae. |
 |
| R2 = Coefficient of determination; RSD = Residual standard deviation; HMN = Hill’s model with n sites |
 |
| Figure 1. Computer-generated views of the energy-minimized conformations of ZEA, AFB1, PAT and DON molecules, respectively. |
ß-D-glucans were defined as complex three-dimensional structures with alternating regions of random coil, single helices and triple helices (Kogan, 2000). In contrast, alkali-soluble ß-D-glucans might make the yeast wall more flexible (Fleet, 1991).
  |
| Figure 2. Graphic representation of the relationship between ß-D-glucan content and the sequestration of ZEA. X1 = amount of total ZEA added to the medium (μg/mL); X2 = ß-D-glucan content of the cell walls of the four strains tested (%); Z = amount of ZEA sequestered by yeast cell walls (μg/mL). |
A powerful physical/chemical modeling technique, together with complementary nuclear magnetic resonance (NMR) and X-ray structural data analysis, was used to assess the overall stability of the modeled molecules in their most stable possible conformations and to allow calculation of the statistical probability of the existence of each conformation. ß-D-glucan spatial organization was calculated and evaluated as a cluster of four triplehelix chains of ß-(1
These helical chains were 1.56 nm apart, with 0.60 nm between helix 360° rotations, consequently defining six ß-D-glucopyranose units per turn of helix, as evaluated by X-ray diffraction. They had the potential to relax their structure and form single helices, depending on surrounding conditions, leading to dynamic behavior of the ß-D-glucans. Thus, environmental conditions such as pH and nature of the solvent can decrease the stability and alter the geometry of ß-D-glucans (Yiannikouris et al., 2004c).
The site-specific interactions between a mycotoxin and the sequestrant were then investigated by means of in silico molecular modeling. ZEA, AFB1, DON and PAT molecules were constructed using NMR and X-ray data (Panneerselvam et al., 1996; Cordier et al.,1990; Marchessault and Deslandes, 1979). A [ZEA + ß-D-glucan] complex was formed by manually positioning mycotoxin molecules in the cavity offered by the ß-(1
Translations plus rotations as well as up and down positioning of mycotoxins were carefully explored inside the ß-D-glucans to find all possible spatial orientations of the interaction (Yiannikouris et al., 2004d). It was concluded that the defining chemical interactions involve weak chemical linkages such as hydrogen bonds and van der Waals forces responsible for a stacking effect occurring between ß-D-glucans and the hydroxyl and cyclic groups of mycotoxins, if available.
The consequence was the production of several in silico models showing mycotoxin molecules caged inside the helix-shaped ß-(1
Different orientations of the docking of AFB1 were found with minor differences in stability. AFB1 seemed to be involved in a major stacking interaction due to van der Waals forces. This chemical interaction between the AFB1 core structure made of an aromatic and a pyran cycle and the residues of ß-D-glucopyranose ring, was responsible for a stabilizing effect due to the geometric symmetry and proximity between AFB1 and the cavities formed in the helical structure of ß-D-glucan (Figure 4). DON was able to create chemical interactions with the modeled ß-D-glucan despite a less stable stereochemistry than other mycotoxins because of its tetracyclic scirpenol ring common to all trichothecenes.
This weak stability of DON could be explained by the tetracyclic scirpenol ring and surrounding hydroxyl, epoxy and ketone groups. However, in spite of the lack of complementary geometry with ß-D-glucan, the overall structure of DON was able to enter the binding site of the helix with or without the presence of the ß- (1
 |
| Figure 3. Computer-generated view of the docking of ZEA into the single helix of ß-(1 |
  |
| Figure 4. Computer-generated views of the docking of mycotoxins into the single helix of ß-(1 |
| Conclusions Basic science in biochemistry is required to complete and support in a meaningful way the advances in applied nutrition needed to extend our understanding of the beneficial role of Saccharomyces cerevisiae cell wall in limiting the toxicological impact of mycotoxins. The plasticity of the structure of ß-D-glucan exhibiting diverse stereochemistry was undoubtedly responsible for the affinity of a large range of mycotoxins. Affinity rates varied widely among toxins due to their structural and physico-chemical disparities. Nevertheless, we concluded that ß-D-glucans may have strong affinities for mycotoxins exhibiting ‘aflatoxinlike,’ ‘deoxynivalenol-like’ or ‘zearalenone-like’ structures. The use of organic material from yeast cell walls such as Mycosorb® (Alltech Inc.) could therefore help sequester mycotoxins in the digestive tract when they are added to the diet and thus mitigate the harmful impact of mycotoxins on animals. This capability could have potentially large applications for future feed quality in view of the emerging ethanol industry by-products of wet or dry distillers grains with or without solubles. Furthermore, considering their structural conformation, glucans are not implicated in interactions with nutrients. Finally, as organic material, they are environmentally safe because of their biodegradable properties after excretion by the animal. |
References
CAST. 2003. Mycotoxins: Risk in plant, animal and human systems. Council for Agricultural Science and Technology. Ames, Iowa.
Chuah, C.T., A. Sarko, Y. Deslandes and R.H. Marchessault. 1983. Packing analysis of carbohydrates and polysaccharides. Part 14. Triple-helical crystalline structure of curdlan and paramylon hydrates. Macromolecules 16(8):1375-1382.
Cordier, C., M. Gruselle, G. Jaouen, D.W. Hughes and M.J. McGlinchey. 1990. Structures of zearalenone and zearalanone in solution: a high-field NMR and molecular modelling study. Magn. Reson. Chem. 28:835-845.
Devegowda, G., M.V.L.N. Raju, N. Afzali and H.V.L.N. Swamy. 1998. Mycotoxin picture world-wide: novel solutions for their counteraction. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 14th Annual Symposium (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, UK, pp. 241-256.
Fleet, G.H. 1991. Cell walls. In: The Yeasts, Vol. 4 (A.H. Rose and J.S. Harrison, eds). Academic Press, London, pp. 199-271.
Grant, P.G. and T.D. Phillips. 1998. Isothermal adsorption of aflatoxin B1 on HSCAS clay. J. Ag. Food Chem. 46(2):599-605.
Kogan, G. 2000. ß-(1,3)- and ß-(1,6)-D-glucans of yeasts and fungi and their biological activity. In: Studies in Natural Products Chemistry, Bioactive Natural Products (Part D), Vol. 23 (A. Rahman, ed). Elsevier Science B.V., The Netherlands, pp. 107-151.
Marchessault, R.H. and Y. Deslandes. 1979. Fine structure of ß-(1,3)-D-glucans: curdlan and paramylon. Carbohydr. Res. 75:231-242.
Panneerselvam, K., E. Rudino-Pinera and M. Soriano-Garcia. 1996. 3,4,5,6,9,10- Hexahydro-14,16-dihydroxy-3-methyl-1H-2-benzoxacyclotetradecin-1,7(8H)-dione (Zearalenone). Acta Crystallograph. Sect. C 52(12):3095-3097.
Yiannikouris, A., L. Poughon, X. Cameleyre, C.-G. Dussap, J. François, G. Bertin and J.-P. Jouany. 2003. A novel technique to evaluate interactions between S. cerevisiae cell wall and mycotoxins: application to zearalenone. Biotechnol. Lett. 25(10):783- 789.
Yiannikouris, A., J. François, L. Poughon, C.-G. Dussap, G. Bertin, G. Jeminet and J.- P. Jouany. 2004a. Adsorption of zearalenone by ß-D-glucans in the S. cerevisiae cell wall. J. Food Prot. 67(6):1195-1200.
Yiannikouris, A., J. François, L. Poughon, C.-G. Dussap, G. Bertin, G. Jeminet and J.- P. Jouany. 2004b. Alkali-extraction of ß-D-glucans from S. cerevisiae cell wall and study of their adsorptive properties toward zearalenone. J. Ag. Food Chem. 52(11):3666- 3673.
Yiannikouris, A., J. François, L. Poughon, C.-G. Dussap, G. Jeminet, G. Bertin and J.- P. Jouany. 2004c. Influence of pH on model ß-D-glucans complexation toward zearalenone. J. Food Prot. 67(12):2741-2746.
Yiannikouris, A., G. André, A. Buléon, G. Jeminet, I. Canet, J. François, G. Bertin and J.-P. Jouany. 2004d. Comprehensive conformational study of key interactions involved in zearalenone complexation with ß-D-glucans. Biomacromolecules 5(6):2176-2185.
Yiannikouris, A., G. André, L. Poughon, J. François, C.-G. Dussap, G. Jeminet and J.- P. Jouany. 2006. Chemical and conformational study of the interactions involved in mycotoxins complexation with ß-D-glucans. Biomacromolecules 7(4):1147-1155.
Related topics:
Authors:

Alltech

Recommend
Comment
Share

Would you like to discuss another topic? Create a new post to engage with experts in the community.



Featured users in Mycotoxins

