Dietary cation-anion balance
Dietary cation-anion balance: a review of definitions and responses in prepartum and lactating cows
As a result of improved nutritional and management practices, milk yields in well managed herds have doubled in less than 30 years. As the genetic potential of our dairy cattle population continues to increase, designing diets with the optimum concentration of minerals becomes very critical. Interrelated effects among dietary concentrations of macrominerals such as Na, K, Cl, and S have been suspected for some time, but little information is available that explains the mechanism of these effects on lactation al performance of the dairy cow.
DCAD and acid-base status
Shohl and Sato (1922) were the first to propose that mineral interrelationships were related to acidbase status. Shohl (1939) proposed that maintenance of normal acid-base equilibrium required excretion of excess dietary cations and anions. He hypothesized that consumption of either excess mineral cations relative to anions or excess anions relative to cations, resulted in acid-base disturbances.
Once animal nutritionists began to test this hypothesis, mineral interrelationships were found to affect numerous metabolic processes. Leach (1979) and Mongin (1980) reviewed related literature and concluded that mineral interrelationships had profound influences. They theorized that for an animal to maintain its acid-base homeostasis, input and output of acidity had to be maintained. It was shown that net acid intake was related to the difference between dietary cations and anions. The monovalent macromineral ions sodium (Na+), potassium (K+) and chloride (Cl-) were found to be the most influential elements in the expression (Mongin, 1980).
SPECIFIC EFFECTS OF MACROMINERAL SALTS ON ACID-BASE STATUS
Nutrient metabolism results in the degradation of nutrient precursors into strong acids and bases.
During normal metabolism, the flux of hydrogen ions (H+) is great. In typical rations fed to dairy cattle, inorganic cations exceed dietary inorganic anions by several millequivalents (meq) per day. Carried with excess dietary inorganic cations are organic anions, which can be combusted to bicarbonate ion (HCO3 -). Therefore, a diet with excess inorganic cations relative to inorganic anions is alkaline and a diet with excess inorganic anions relative to cations is acidogenic.
NUTRITIONAL FACTORS RELATED TO CATIONANION INTERRELATIONSHIPS
Leach (1979) and Mongin (1980) reviewed nutritional concepts related to cation-anion interrelationships. Historically, nutritionists intuitively knew it was difficult to evaluate the effect of one macromineral without considering the influences of others. Early concepts evaluated total ash, mineral ratios, and differences between two or more of the macrominerals.
Acid or alkaline ash
Nutritionists first investigated the alkalinity and acidity of the diet under the acid or alkaline ash concept (Shohl, 1939). It was recognized that human food had either an acid or alkaline ash. When food is metabolized in the body, organic anions such as acetate, citrate, malate, etc., are oxidized.
Inorganic cations originally associated with these organic anions remain. Because organic anions can buffer H+ ions generated through metabolism, a food with a large amount of organic anions (and thus inorganic cations) was considered alkaline. The pH of the ash represented the acid or alkaline nature of human food.
DIETARY CATION-ANION DIFFERENCE (DCAD)
Blood pH is ultimately determined by the number of cation and anion charges absorbed into the blood.
If more anions than cations enter the blood from the digestive tract, blood pH will decrease. Mongin (1980) was one of the first to propose a three-way interrelationship among dietary Na, K and Cl. He proposed that the sum of Na and K minus Cl (in meq per 100 g diet DM) could be used to predict net acid intake. This sum is commonly called the dietary cation-anion balance (Tucker et al., 1988) or dietary electrolyte balance (West et al., 1991).
However, Sanchez and Beede (1991) coined the term cation-anion difference to represent, more precisely, the mathematical calculation used and to avoid the erroneous connotation that mineral cations truly are balanced with mineral anions in the diet.
Expressed in its fullest form, DCAD would be written as:
| Equation 1 | |
| DCAD = meq [(Na + K + Ca + Mg) - (Cl + S + P)] | |
| 100 g of dietary DM |
A problem with including the multivalent macrominerals (calcium (Ca), magnesium (Mg), phosphorus (P), and sulfur (S)) in the DCAD expression for ruminants relates to the variable and incomplete bioavailability of these ions compared to Na, K and Cl. The expression that has been used most often in non-ruminant nutrition is the monovalent cation-anion difference (Equation 2).
This expression was considered superior for nonruminant nutritionists because it comes closest to representing feed ions that are completely dissociated and solubilized from their respective salts and absorbed into the body.
| Equation 2 | |
| DCAD = meq (Na + K - Cl)/100 g dietary DM |
Because of the additional use of sulfate salts in prepartum dairy rations, the expression that has gained the most acceptance in ruminant nutrition, and is the most common expression used in ration software, is:
| Equation 3 | |
| DCAD = meq [(Na + K) - (Cl + S)]/100 g dietary DM |
CALCULATING DCAD
To actually calculate DCAD using Equation 3, mineral concentrations are first converted to meq as follows:
| meq/100 g = | (mgs)(valence) |
| (atomic weight in g) |
As an example, the meq (Na + K) – (Cl + S) value of a diet with 0.1% Na, 0.65% K, 0.2% Cl and 0.16% S (minimum recommendation for dry cows; NRC, 1989) will be calculated. There are 100 mg Na (0.10% = 0.10 g/100 g or 100 mg/100 g), 650 mg K (0.65% K), 200 mg Cl (0.2% Cl), and 160 mg S (0.16% S) per 100 g diet DM. Therefore, this diet contains:
| meq Na = | (100 mg)(1 = valence) | = 4.3 meq Na |
| (23 g = atomic weight) |
| meq K = | (650 mg)(1 = valence) | = 16.7 meq K |
| (39 g = atomic weight) |
| meq Cl = | (200 mg)(1 = valence) | = 5.6 meq Cl |
| (35.5 g = atomic weight) |
| meq S = | (160 mg)(2 = valence) | = 10.0 meq S |
| (32 g = atomic weight) |
The next step is to add the meq from the cations and subtract the meq of the anions:
meq (Na + K) - (Cl + S) = |
(4.3 + 16.7) – (5.6 + 10.0) = + 5.4 meq/100 g diet DM |
Another simpler way to calculate DCAD is:
| DCAD = |
[(%Na in DM/0.023)+(%K in DM/0.039)]-[(%Cl in DM/0.0355)+(%S in M/0.016)] |
For example, using the same numbers as above, the calculated DCAD equals (0.10% Na/0.023) + (0.65% K/0.039) - (0.2% Cl/0.0355) - (0.16% S/ 0.016) = +5.4 meq/100g diet DM.
Note that values calculated per 100 g are 10 times less than on a per kg basis (100 g = kg/10). Note also that the DCAD equation with only Na, K, and Cl in it yields a value approximately 10 DCAD units higher than with the equation with Na, K, Cl, and S in it (assuming S is equal to 0.16%).
Three sources of error in calculating DCAD Units
Some papers report the units of DCAD per 100 g DM and some report DCAD per kg DM. This can cause a 10 fold calculation error, which can cause a serious formulation error if not corrected before balancing the diet.
Sulfur in the DCAD equation
For reasons mentioned above, the sulfate ions were not initially included in the DCAD equation.
However, because of the extensive use of sulfate salts in dry cow rations, the DCAD equation [(Na + K) - (Cl + S)] has become more common in ration formulation programs. Because S changes the DCAD calculation drastically, this has led to errors in calculating DCAD and comparing information from the literature.
The Spartan Dairy Ration Program calculation The popular Spartan Dairy Program took a novel approach to calculating DCAD. Because of the problems mentioned above relative to divalent ions, it considered only the inorganic form of S in the calculation. Therefore, depending on the amount of organic S sources, the calculation of DCAD could be off by 10 to 20 meq/100 g of DM. One way to use the program to calculate the DCAD in the same way that has been done in research studies (using Equation 3) is to categorize all ingredients as minerals before checking the DCAD calculation.
However qualifying the amount of organic vs. inorganic S may be a more appropriate method to account for the effect of DCAD on acid-base status because the impact of S on acid-base status is much less than earlier projected (Goff et al., 1997a).
DCAD and the prepartum cow
The period from three weeks prepartum to parturition is an extremely critical time for the dairy cow; possibly defining her entire lactational performance (Wang, 1990).
Many years ago, researchers discovered that an acidic diet caused the concentration of blood Ca to increase. This led to the practice of feeding a diet with more anions relative to cations to help reduce milk fever problems. The increased blood Ca in these cows not only prevented milk fever, it also reduced problems like retained placenta and displaced abomasum. These problems were tied to a Ca deficiency that prevented the muscles from contracting. Therefore, nutritionists began feeding prepartum cows diets with less cations than anions to help increase blood Ca around the time when it was very deficient at calving. These diets are described as anionic diets or diets with a low or negative cation-anion difference.
Milk fever may affect 5-7% of all high producing adult dairy cows in the United States (Jordan and Fourdraine, 1993). In addition, the prevalence of subclinical hypocalcemia may be as high as 66% for multiparous dairy cows following calving (Beede et al., 1992). Research indicates that cows with clinical milk fever produce 14% less milk in the subsequent lactation and their productive life is reduced approximately 3.4 years when compared to non-milk fever cows (Block, 1984; Curtis et al., 1984). Furthermore, cows that recover from milk fever have an increased risk of ketosis, mastitis (especially coliform mastitis), dystocia, left displaced abomasum, retained placenta and milk fever in the subsequent lactation (Curtis et al., 1984; Wang, 1990; Oetzel, et al. 1988). Guard (1996) estimated that the cost associated with a single case of milk fever is approximately $334, when considering lost production and income, veterinary costs and treatment costs.
FEEDING ANIONIC SALTS
Feeding anionic salts or manipulating the dietary cation-anion difference of the diet has become a common approach on dairies that can accommodate multiple dry cow groups. Feeding prepartum dry cows less Na and K relative to Cl and S (i.e., a negative DCAD diet) increases blood Ca at calving, presumably by increasing bone mobilization and(or) absorption of Ca in response to changes in acidbase status. Studies have shown that when dry cows were fed diets with negative DCAD, milk fever incidence was reduced drastically, and in some cases eliminated.
It is important to emphasize that for the trials in which negative DCAD aided in prevention of milk fever, dietary concentrations of Ca were high (approximately 1.5% Ca). Negative DCAD increases urinary excretion of Ca; therefore, if dietary Ca were low with a negative DCAD, hypocalcemia may occur, regardless of and separately from milk fever. Conversely, high dietary Ca with low DCAD may be necessary for this method to be successful.
However, the optimal dietary Ca content has not been established.
RECENT RESEARCH
Research on the DCAD expression
Because of differences in bioavailability of each mineral element in the DCAD expression, the functional equation most applicable in practical situations differs. Based on the bioavailability figures for Ca, Mg and P from NRC (1989), Goff et al. (1997a,b) suggested that Equation 4 was more appropriate. In this calculation Na, K and Cl were considered 100% bioavailable; and the bioavailability of 60% for S was based on work of Tucker et al. (1991).
| Equation 4 | |
| meq([Na + K + 0.38Ca + 0.30Mg) - (Cl + 0.60S +0.5P)]/ 100g of dietary DM |
Goff and Horst (1997) then compared the acidifying effects of dietary hydrochloric acid or sulfuric acid on urine pH of nonlactating Jersey cows. Sulfuric acid exhibited about one-third of the acidifying power (e.g., change in urine pH) of hydrochloric acid.
Sulfuric acid would be considered the most bioavailable chemical form of the sulfate anion (SO4 -2) compared with other mineral sources of sulfate, such as magnesium sulfate, calcium sulfate, and ammonium sulfate.
Goff et al. (1997a) compared the relative acidifying strengths of six anion sources with a similar animal model. Urine samples were taken 4 hr after feeding on day 3, 4 and 5 of each experimental period in which a different anion source was fed. Urine pH values of multiparous non-lactating Jersey cows fed hydrochloric acid, calcium chloride, ammonium chloride, calcium sulfate, magnesium sulfate, and elemental S were 6.2, 7.1, 7.0, 7.6, 7.9 and 8.2, respectively. The order of strength from strongest to weakest acidifiers was hydrochloric acid, ammonium chloride, calcium chloride, calcium sulfate, magnesium sulfate, and elemental sulfur.
Overall, the chloride salts were more acidogenic than the sulfates. Elemental S had no effect on acidbase status, as one should expect; although occasionally elemental S is found as a source of anion in mineral supplements for close-up diets.
These new data cause us to question what the most appropriate DCAD equation should be and what anion sources are most appropriate for supplementation. Based on results of these two experiments, Goff et al. (1997b) suggested that a more biologically or functionally correct DCAD equation might be:
| Equation 5 | |
| meq [(Na + K + 0.15Ca + 0.15Mg) - (Cl + 0.20S + 0.3P)]/ 100g of dietary DM |
Recently, Rodriguez et al. (1997) found no difference in urine or blood plasma pH when non-lactating, non-pregnant Holstein cows were fed diets with either 0.5 or 2.0% Ca (from calcium car bonate) across diets with DCAD set at about -10 (meq[(Na + K) - (Cl + S)]/ 100g of dietary DM).
In both trials the high Ca diets reduced DM intake when anionic salts were fed. Therefore, excessively high dietary Ca levels are not recommended. We continue to recommend 120 to 150 g of Ca/day for cows fed anionic salts. When more positive DCAD concentrations are used, feed a dietary Ca concentration at the lower end of this recommendation.
When more negative concentrations are fed, bring dietary Ca toward the upper end of this recommendation.
Remember that the source of Ca influences its overall effect. For example, Ca from alfalfa forage is less bioavailable than calcium carbonate, and calcium carbonate is less bioavailable than calcium chloride.
Results of these experiments have stimulated considerable discussion of DCAD equations and supplementation of anions. There is no consensus on which equation to use. If a more precise estimate is required, the weighted equations are probably most effective. However, the simplest and most practical approach may be to use the three-element equation. Until additional data are available, most researchers continue to recommend the fo urelement expression (Equation 3).
Effects of DCAD on lactational performance
STUDIES USING THE (Na + K – Cl) DCAD EXPRESSION
Coppock (1986) reviewed the influence of dietary DCAD as (Na + K – Cl) on lactational performance of dairy cattle. In general, it was noted that ruminants could withstand higher DCAD than poultry or pigs.
In summarizing trials after the fact, he found that DCAD had no influence between +10 to +40.
Escobosa et al. (1984) studied effects of feeding either 0.23% sodium chloride (NaCl), 0.23% NaCl plus 2.28% calcium chloride (CaCl2), or 0.23% NaCl plus 1.7% sodium bicarbonate (NaHCO3). Diets had -14, +20 or +35 DCAD. It was found that the excess Cl (-14 DCAD) depressed feed intake and resulted in acidosis.
Kentucky researchers (Tucker et al., 1988) appear to have been the first to conduct a study specifically designed to evaluate the effect of DCAD on acid-base status and lactational performance of dairy cattle. They compared diets formulated with -10, 0, +10 or +20 DCAD. A diet with a +20 DCAD improved dry matter (DM) intake by 11% and milk yield by 9% compared with a diet with -10 DCAD. Blood bicarbonate (HCO3 -) increased linearly with increasing DCAD, which indicated an improvement in acid-base status with high DCAD compared with low DCAD. They concluded that responses to increasing DCAD were independent of specific Na, K or Cl effects.
Because lactation diets typically contain greater DCAD than +20, these results were initially more theoretical than practical. For example, the NRC (1989) minimum Na, C1, and S requirements indicate that DCAD should be greater than about +25.
The next question that had to be answered was whether responses would continue to increase with diets above +20 DCAD. West et al. (1991) in Georgia answered part of this question when they evaluated diets with up to +40 meq/100 g of diet DM. Their study used two 4 x 4 Latin squares blocked by environmental temperature (cool vs. hot). Separate squares included four Holstein and four Jersey cows. Diets contained +2.5, +15, +27.5 or +40 DCAD. No effect of environment was reported, but increasing DCAD from +2.5 to +27.5 increased DM intake, milk yield and blood bicarbonate (HCO3 -). These findings suggested that performance was depressed with lower DCAD.
At +27.5 DCAD, negative effects were overcome.
Above +27.5 DCAD no additional improvement was attained.
In another study by this group (West et al., 1992), diets with even higher DCAD (+10, +21.7, +33.4 and +45.1) were fed to a total of 16 lactating dairy cows during hot weather. Source of cation (Na or K) used to manipulate DCAD also was compared.
Increasing DCAD increased DM intake linearly, independent of Na or K source. Yield of 3.5% fatcorrected milk (FCM) was not affected by DCAD or cation source. Milk fat concentration was greater with Na- compared with K-manipulated diets (3.92 vs. 3.62%). Blood pH increased linearly; whereas blood HCO3 - increased curvilinearly; and there was no effect due to cation source on acid-base status.
Their results indicated that increasing DCAD improved DM intake and acid-base status in a manner consistent with other studies. In general, DCAD was independent of a specific Na or K effect.
The influence of Na, K and Cl at constant DCAD was evaluated by Tucker and Hogue (1990). Diets were formulated to provide +32 DCAD in either: a basal diet (adequate in dietary Na, K, and Cl), a
basal diet containing an additional 1.17% NaCl, or a basal diet containing an additional 1.56% potassium chloride (KCl). Fifteen mid lactation cows were assigned to replicated 3 x 3 Latin squares.
The KCl-fed cows consumed more DM and had lower milk fat percentage than NaCl-fed cows, but there were no differences in milk production. It was concluded that dietary DCAD was a more important determinant of dietary impact on systemic acid-base status than actual dietary concentrations of Na, K, and Cl.
To model the effect of multiple concentrations of DCAD across a variety of diets and management conditions, Sanchez et al. (1994b) assembled a large database from 10 years of studies with mid-lactation cows in Florida. Combining data from many studies into one analysis verified the curvilinear response to DCAD. Dry matter intake, milk yield and FCM yield were all maximized at +38 DCAD (Na + K – Cl)/100 g. These models were validated by comparing them to independent data of Tucker et al. (1988), West et al. (1991) and West et al.
(1992). The DCAD models developed from the data base predicted results very well. For DMI, the DCAD model and independent data set predictions differed only by an average of 2.87% (range 0.19 to 12.27%). For milk yield, they differed only by an average of 2% (range 0.13 to 7.94%). Absolute deviations between the data base DCAD models and independent data set predictions (corrected for experiment effects) ranged from 0.24 to 1.22 kg/d for DM intake and 0.07 to 0.60 kg/d for milk yield.
A summary of studies that used the Na + K - Cl expression is presented in Table 1.
Research with the DCAD = meq[(Na + K) – (Cl + S)] equation A large study with 48 cows and 15 dietary treatments was conducted by Sanchez et al. (1994a) to investigate lactational and acid-base responses to DCAD as [(Na + K) – (Cl + S)]. Treatments consisted of combinations of Na, K and Cl, so that DCAD ranged from 0 to +50 [(Na + K) – (Cl + S)]/100g DM. The basal diet was 54.5% concentrate, 5.5% cottonseed hulls and 40% corn silage (DM basis). Dry matter intake and milk yield was highest when DCAD was between +17 to +38 and +25 to +40, respectively (Figure 1). There was one odd treatment (a low Cl, high K, and high Na combination) that may have caused a Cl deficiency.
Had that treatment not been included, the regression line would have shifted to the right. Blood HCO3 - (P = 0.09) also responded quadratically to increasing DCAD. Blood HCO3 - was maximized with +38 DCAD. In support of conclusions of Tucker et al.
(1988) and West et al. (1991; 1992) results of this study indicated that diets with less than +20 DCAD depressed blood HCO3 - and should not be fed. Note that with most DCAD studies (this one included), dietary carbonate and bicarbonate concentrations are confounded with DCAD. Because these salts are used to elevate DCAD, DCAD effects cannot be separated from well-known ruminal and systemic buffering effects of carbonate and bicarbonate salts.
Research with the DCAD [(Na + K) – (Cl + S)] equation throughout different phases of lactation
Three switchback experiments (Delaquis and Block, 1995) were conducted with 12 cows each in early, mid, and late lactation. Each experiment compared two DCAD levels calculated as [(Na + K) – (Cl + S)]/100g DM. Increasing DCAD from +5.5 to + 25.8 in early lactation and from +14.0 to + 37.3 in midlactation increased DM intake and milk production.
These effects were not observed in late lactation (with either +20.0 or +37.5 DCAD). Concentration of blood HCO3 - was decreased in early lactation and excretion of carbonate ions in urine was reduced by a lower DCAD at all stages of lactation.
Responses in this study are consistent with the effect of DCAD on acid-base status observed in other studies; however, this study supports the concept that response to DCAD is affected by stages of lactation.
| Table 1. Summary of the effect of DCAD on milk, intake and blood bicarbonate concentrations in studies using the (Na + K) – Cl) DCAD expression. |
 |
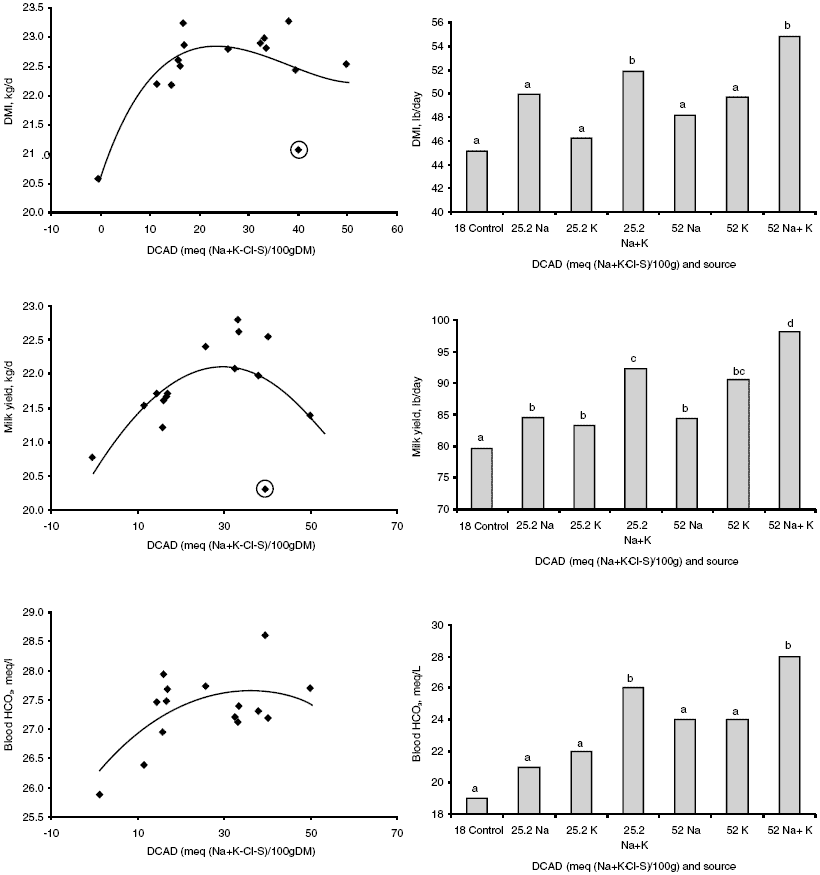 |
| Figure 1.Dry matter intake, milk yield, and blood bicarbonate response to DCAD in mid lactation (left panel) and early lactation (right panel) cows. Data in left panel are from Sanchez et al. (1994a) (treatments circled are from a low Cl, high K, and high Na treatment combination that may have caused a Cl deficiency). Data in right panel are from Block, McGill University 1999 (unpublished data). Block evaluated both DCAD concentration and source (either Na, K or a proprietary combination of both) in ten early lactation cows (weeks 1 – 10 in milk) per treatment. Different superscripts indicate a statistical difference between treatment means. |
Perhaps the most complete study on the effects of DCAD on early lactation dairy cows conducted to date is an unpublished trial at McGill University in 1999. The trial involved a control diet with no added Na or K (+18 DCAD) and two higher (+25 and +52) DCAD diets fed to early lactation (0 – 10 weeks in milk) Holstein cows. In order to determine the individual or combined effects of Na and K, within the higher DCAD diets sources of DCAD were either sodium bicarbonate or potassium carbonate alone or a proprietary combination of both. The combination of Na and K yielded the best response in DM intake and milk production, and the +52 DCAD diet yielded the highest milk production response (Figure 1). The combinations of Na and K also resulted in the highest blood bicarbonate concentrations. A summary of studies that used the (Na + K) - (C1 + S) expression is presented in Table 2.
Potassium as the source of increased DCAD
The above positive responses observed with combinations of Na and K point to the unique role of dietary K, which has been particularly evident during heat stress. Heat-stressed cows lose K via sweat; and milk is actually higher in K than Ca.
Thus, the heat-stressed dairy cow is often Kdeficient.
Research conducted by Joe West in Texas, where potassium carbonate was the source of dietary K, indicated that there is a linear response to dietary K in heat-stressed dairy cows. (West et al., 1986; West et al., 1987a,b). Figure 2 shows the FCM responses from three studies and an overall regression response to varying dietary K via potassium carbonate. The regression line indicates that every 1% increase in K can raise FCM by 8.9 lb/day.
DCAD feeding strategy
Detailed feeding strategies for using anionic salts including recommended salts to use, length of feeding interval, and specific precautions can be found in several excellent reviews (Beede 1995; Horst et al., 1997). The most common recommendation is to add sulfate salts until Mg and S are maximized at 0.4 to 0.45% and then chloride salts until DCAD is lowered to -10 to -15 meq/100g dietary DM. Calcium intake should be increased to approximately 120 to 150 g per cow/day. New information on this concept is outlined below.
| Table 2.Summary of the effect of DCAD on milk, intake and blood bicarbonate concentrations in studies using the (Na + K) – (Cl + S) DCAD expression. |
 |
 |
| Figure 2. Fat-corrected milk (3.5%) response and overall regression response to feeding various potassium concentrations (as potassium carbonate) to heat stressed mid-lactation dairy cows. (West et al., 1986; West et al., 1987a,b). |
OPTIMAL DCAD AND DOSE OF ANIONIC SALTS
Controlled experiments have not yet determined the optimal DCAD. The recommended target DCAD of -10 to -15 meq/100g dietary DM may be lower than needed to achieve the desired changes in acidbase status and subsequent increases in blood Ca.
However, this range of DCAD provides a margin of safety to account for varying K concentrations in feeds and K consumed from pasture or freechoice hay. Research from the University of Idaho addressed this question further and established a numerical relationship between DCAD and blood calcium (Giesy et al., 1997). The study also showed a tight relationship between DCAD and urine pH.
Urine pH can be monitored on farm to make sure diets are formulated correctly.
WHAT IF THE BASAL DIETARY DCAD IS TOO HIGH?
Horst et al. (1997) suggested the maximum amount of anions that can be added before intake declines is about 30 meq. This means if the basal DCAD is about +20, then the DCAD can be lowered to -10 with 30 meq of anions. However, when the basal DCAD is greater than +30 or +40, what then should be the strategy? The first priority would be to remove as much high K feedstuff as possible (some hay samples contain more than 4.0% K). Once that is done, if the basal DCAD is still +30 or +40, there are two options. First, add more than 30 meq of anions and lower the DCAD to -10. This potentially could lower intake by the prepartum cow and lead to other metabolic problems (Bertics et al., 1992).
However, research conducted by Joyce et al. (1997) at the University of Idaho demonstrated that reducing intake by feeding anionic salts prepartum is not always detrimental. In that study (Figure 3) prepartum Holstein cows were fed either a grassbased (+30 DCAD), an alfalfa-based (+35 DCAD), or an alfalfa plus anionic salts diet (-7 DCAD). The cows fed the -7 DCAD diet had lower intakes prepartum but greater intakes postpartum compared with cows fed the other treatments. The -7 DCAD diet did increase blood Ca, which apparently overcame any negative effect of reducing intake.
The relationship between DCAD, intake and postpartum performance was similar to that found in a recent experiment with periparturient cows (Moore et al., 1997).
Vagnoni and Oetzel (1997) studied the effects of DCAD on DM intake and acid-base status. Four diets were evaluated: (1) control, (2) Biochlor (Biovance Technologies, Omaha NB), (3) magnesium sulfate and ammonium chloride, and (4) magnesium sulfate, calcium sulfate and calcium chloride. Urine pH was reduced by feeding anionicsalts and Biochlor. Biochlor was the most effective treatment in reducing urine pH, followed by treatment 2 then 3.
 |
| Figure 3. Intake response to varying diets [a grass-based (+30 DCAD), an alfalfa-based (+35 DCAD), or an alfalfa plus anionic salts diet (-7 DCAD)] fed prepartum to Holstein cows. After parturition, all cows were fed a similar alfalfa-based lactation ration (from Joyce et al., 1997). |
The second option to try if the basal DCAD is too high would be to reduce the DCAD as much as possible, for example to +10 or 0. A potential problem with this option is that the DCAD concentration may not be low enough to control hypocalcemia.
When this is done, it is recommended to only increase dietary Ca partially (Beede, 1995).
Experiment to determine response to varying DCAD
We completed an experiment (Giesy et al., 1997) that provided new data on the above two options.
The objective of the study was to determine the blood Ca responses to varying DCAD concentrations. Four non-pregnant, non-lactating Holstein cows were used in a complete 4 x 4 latin square design. Each cow was fed one of four DCAD concentrations, +30, +10, -10, or -30 meq/ 100g DM. Diets were fed as total mixed rations with alfalfa hay, grass hay and alfalfa silage as the forages. Cows were fed these diets for 14 days then given EDTA intraveneously to mimic subclinical hypocalcemia. Blood samples were taken immediately prior to the onset of infusion of EDTA and once every 30 minutes thereafter for 8 hrs.
Blood was analyzed for both total and ionized Ca.
Urine pH also was measured. From the results of this study, we observed that serum total Ca was highly variable and, although somewhat responsive to DCAD, did not seem to correlate well with DCAD. Blood ionized Ca, the freely available fraction of Ca, was much less variable and was very responsive to DCAD. Figure 4 shows the blood ionized Ca response to varying DCAD. We also saw an increase in blood Ca at each of the decreasing levels of DCAD. This indicates that increases in blood Ca can be achieved even when DCAD is not lowered to -10.
USING URINE PH TO MONITOR DCAD PROGRAMS
Upon feeding anionic salts, urine pH changes quickly (within 2-4 days). Monitoring urine pH, therefore, can be a useful tool to determine whether the ration is having the desired physiological effect.
The urine pH response to DCAD from the study of Giesy et al. (1997) is shown in Figure 5. This tight relationship between DCAD and urine pH was similar to that found in a recent experiment with periparturient cows (Moore et al., 1997). In that study they fed diets for 21 days to prepartum cows with DCAD of +14, 0, and -5 meq (Na + K) - (Cl + S)/100g of dietary DM. Supplemental anions were provided from calcium chloride, magnesium sulfate, and magnesium chloride. Total dietary Ca varied (0.44, 0.97, and 1.5% Ca) with the three decreasing DCAD levels, respectively. The source of supplemental Ca was from increasing calcium chloride and calcium carbonate in the 0 and -5 meq diets. Urine pH of close-up cows immediately before calving was 7.98, 7.0, and 6.21 for +14, 0, and -5 meq, respectively.
 |
| Figure 4. Blood ionized Ca response to four levels of DCAD following infusion with EDTA to mimic hypocalcemia (from Giesy et al., 1997). |
 |
| Figure 5. Urine pH response to four levels of DCAD (from Giesy et al., 1997). |
A common question that arises when urine pH monitoring programs are instituted is “When should urine pH be collected?” Goff and Horst (1998) evaluated the effect of time after feeding on urine pH. In their first study they fed 21 nonpregnant dry Jersey cows twice per day (at 8 a.m. and 8 p.m.) either a +32 or -14 DCAD diet ((meq Na + K) – (Cl + S)/100g diet DM). The negative DCAD diet had hydrochloric acid (HCl) added to it. Urine pH was measured just before feeding at 8 a.m. and again 3, 6, 9, and 12 hrs later.
In this twice daily feeding study, urine pH averaged 8.2 for controls and 7.3 for HCl treatment throughout the day, but there was no significant diurnal variation in urine pH (Figure 6a). In their second trial, 25 dry cows were fed just once per day. As before, the HCl diet significantly reduced urine pH, but in contrast to the first study, there was a significant diurnal variation in urine pH (Figure 6b). Urine pH of cows given HCl was 7.04 at feeding time and 6.17 at 3 hrs after feeding. The study demonstrated there could be diurnal shifts in urine pH when close-up cows are only fed once per day.
When establishing an on-farm urine pH measuring protocol, the best strategy would be to collect urine from about 10 cows at the same time of day each week, preferably 2-6 hours after feeding. This can be done on-farm, using standard pH paper or a field pH meter. If the average urine pH is much greater than 6.5 (the target level), the ration is not affecting acid-base status enough to significantly alter blood Ca concentration at calving. Measure urine pH the same time every week to ensure that the desired effect is being maintained.
Conclusions
Much more information on the use of urine pH as a monitoring tool is now available, including the quantitative relationship to DCAD and the effect of feeding on diurnal variations in urine pH. |
References
Author: ELLIOT BLOCKBeede, D.K., C.A. Risco, G.A. Donovan, C. Wang, L.F. Archbald, and W.K. Sanchez. 1992. Nutritional management of the late pregnant dry cow with particular reference to dietary cationanion difference and calcium supplementation. Proc. 24th Ann. Convention Am. Assoc. Bovine Practitioners, p. 51.
Beede, D.K. 1995. Macromineral element nutrition for the transition cow: Practical implications and strategies. Proc. Tri-State Nutrition Conf., Ft. Wayne, IN, p. 175.
Bertics, Sandra J. , Ric R. Grummer, Carlos Cadorniga-Valino, and Emily E. Stoddard. 1992. Effect of prepartum dry matter intake on liver triglyceride concentration and early lactation. J. Dairy Sci. 75:1914-1922.
Block, E. 1984. Manipulating dietary anions and cations for prepartum cows to reduce incidence of milk fever. J. Dairy Sci 67:2939.
Coppock, C.E. 1986. Mineral utilization by the lactating cow—chlorine. J. Dairy Sci. 69:595.
Curtis, C.R., H.N. Erb, C.J. Sniffen, and R.D. Smith. 1984. Epidemiology of parturient paresis: predisposing factors with emphasis on dry cow feeding and management. J. Dairy Sci. 67:817.
Delaquis A.M. and E. Block. 1995. Dietary cationanion difference, acid-base status, mineral metabolism, renal function, and milk production of lactating cows. J. Dairy Sci. 78:2259
Escobosa, A., C.E. Coppock, L.D. Rowe Jr., and C.E. Gates. 1984. Effects of dietary sodium bicarbonate and calcium chloride on physiological responses of lactating dairy cows in hot weather. J. Dairy Sci. 67:574.
Giesy, J.G., W.K. Sanchez, M.A. McGuire, J.J. Higgins, L.A. Griffel, and M.A. Guy. 1997. Quantifying the relationship of dietary cation-anion difference to blood calcium in cows during hypocalcemia. J. Dairy Sci. 80(Suppl. 1):142.
Goff, J.P. and R. L. Horst. 1997. Comparison of sulfuric acid with hydrochloric acid as a source of acidifying anions. J. Anim. Sci. 75(Suppl. 1):98.
Goff, J.P.,and R.L. Horst. 1998. Effect of time after feeding on urine pH determinations to assess response to dietary cation-anion adjustment. J. Dairy Sci. 81(Suppl. 1):44.
Goff, J.P., R. Ruiz, and R.L. Horst. 1997a. Relative acidogenic activity of commonly used anionic salts - re-thinking the dietary cation-anion difference equations. J. Dairy Sci. 80(Suppl. 1):169.
Goff, J.P. R. L. Horst, T.A. Reinhardt, and D.R. Buxton. 1997b. Preventing milk fever in dairy cattle. Proc. Tri-State Dairy Nutrition Conf., Ft. Wayne, IN, p. 41.
Guard, C. 1996. Fresh cow problems are costly: culling hurts the most. Page 100 in Proc. 1994 Ann. Conf. Vet. Cornell Univ., Ithaca, NY. Horst, R.L., J.P. Goff, T.A. Reinhardt, and D.R. Buxton. 1997. Strategies for preventing milk fever in dairy cattle. J. Dairy Sci. 80:1269.
Jordan E.R., and Fourdraine R. 1993. Characterization of the management practices of the top DHI milk producing herds in the country. J. Dairy Sci. 76:3247
Joyce, P.W., W.K. Sanchez, and J.P. Goff. 1997. Effect of anionic salts in prepartum diets based on alfalfa. J. Dairy Sci. 80:2866.
Leach, R.M. 1979. Dietary electrolytes: Story with many facets. Feedstuffs. April 30, p. 27.
Mongin, P. 1980. Electrolytes in nutrition: review of basic principles and practical application in poultry and swine. In: Third Ann. Int. Mineral Conf. Orlando, FL. p.1.
Moore, S.J., M.J. VandeHaar, B.K. Sharma, T.E. Pilbeam, D.K. Beede, H.F. Bucholtz, J.S. Liesman, R.L. Horst, and J.P. Goff. 1997. Varying dietary cation anion difference (DCAD) for dairy cattle before calving. J. Dairy Sci. 80(Suppl. 1):170.
National Research Council. 1989. Nutrient Requirements of Dairy Cattle. 6th. rev. ed. Natl. Acad. Sci., Washington, D.C.
Oetzel, G.R., J.D. Olson, C.R. Curtis, M.J. Curtis, and M.J. Fettman. 1988. Ammonium chloride and ammonium sulfate for prevention of parturient paresis in dairy cows. J. Dairy Sci. 71:3302.
Rodriguez, L.A., T.E. Pilbeam, and D.K. Beede. 1997. Effects of dietary anion source and calcium concentration on dry matter intake and acid-base status of nonlactating nonpregnant Holstein cows. J. Dairy Sci. 80(Suppl. 1): 241.
Sanchez, W.K.. 1999. Another new look at DCAD for the postpartum dairy cow. Proc. 1999 Mid- South Ruminant Nutrition Conf. p.
Sanchez, W.K. and D.K. Beede. 1991. Interrelationships of dietary Na,K and Cl and cation-anion difference in lactation rations. In: Proc. Florida Rum. Nutr. Conf. University of Florida, Gainesville. p.31.
Sanchez, W.K., D.K. Beede, and J.A. Cornell. 1994a. Interactions of sodium, potassium, and chloride on lactation, acid-base status, and mineral concentrations. J. Dairy Sci. 77:1661.
Sanchez, W.K., D.K. Beede, and M.A. DeLorenzo. 1994b. Macromineral element interrelationships and lactational performance: Empirical models from a large data set. J. Dairy Sci. 77:3096.
Shohl, A.T. 1939. Mineral metabolism. Reinhold Publishing Corp, New York.
Shohl, A.T. and A. Sato. 1922. Acid-base metabolism. I. Determination of base balance. J. Biol. Chem. 58:235.
Tucker, W.B., G.A. Harrison, and R.W. Hemken. 1988. Influence of dietary cation-anion balance on milk, blood, urine, and rumen fluid in lactating dairy cattle. J. Dairy Sci. 71:346.
Tucker, W.B. and J.F. Hogue. 1990. Influence of sodium chloride or potassium chloride on systemic acid-base status, milk yield, and mineral
Arm & Hammer Animal Nutrition Group, Church and Dwight Co., Inc., Princeton, NJ, USA

Very useful information.

Good article about dietary cation-anion balance in lactating cows. See "Another New Look at DCAD for the Prepartum Dairy Cow" by William K. Sanchez, Ph. DK.






KeyShure Effects of an organic source of copper, manganese and zinc on dairy cattle productive performance, health status and fertility

FIGAP MEXICO 2026 - INTERNATIONAL EXHIBITION





United States