Vitamin A in poultry and swine nutritional immunology
The role of vitamin A in non-ruminant immunology
Vitamin A (retinol) is an essential micronutrient with a crucial role in the immune system of non-ruminant animals, such as swine and poultry. It includes three chemical compounds with distinct properties and functions in the body: retinol, retinal, and retinoic acid. In monogastric feed, vitamin A is primarily present in the form of retinyl esters. The metabolism of dietary vitamin A esters involves their conversion to retinol, which is then transported to different tissues and cells for further metabolism into active forms such as retinoic acid. These active forms of vitamin A have been found to play a crucial role in regulating both innate and adaptive immune responses. Specifically, they are involved in the differentiation, proliferation, and function of immune cells such as T and B lymphocytes, as well as dendritic cells. Vitamin A deficiency can lead to impaired cellular immunity, reduced antibody production, and consequently an increased susceptibility to infections. In swine and poultry, hypovitaminosis A can also affect gut-associated lymphoid tissues, leading to gut-related health problems and compromised growth performance. On the other hand, vitamin A supplementation has been shown to have immunomodulatory effects on non-ruminant immune responses. By administering or supplementing retinol, immune cell proliferation, antibody production, and cytokine secretion can be enhanced, which can ultimately result in improved immune function and disease resistance. Therefore, vitamin A has potential applications as an immuno-micronutrient for improving health and preventing diseases in swine and poultry. However, the optimal dosage and timing of vitamin A supplementation need to be carefully determined based on the specific requirements of different non-ruminant species and their production stages. Overall, a better understanding of the role of vitamin A in non-ruminant nutritional immunology could have significant implications for animal health and productivity and could inform the development of effective dietary strategies to optimize immune function and prevent diseases in swine and domestic fowl. This review paper aims to offer valuable insights into the role of vitamin A in the nutritional immunology of non-ruminants while also emphasizing the current gaps in knowledge and potential areas for further research.
KEYWORDS vitamin A, retinol, non-ruminants, swine, poultry, supplementation, immunity.
1 Introduction
2 Vitamin A metabolism and function
3 The impact of vitamin A deficiency on immune function
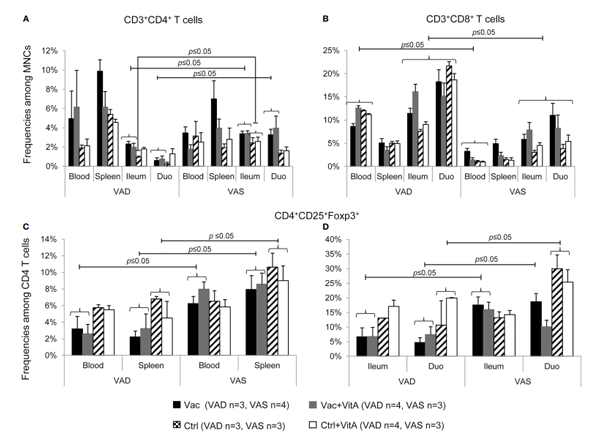
4 The immunomodulatory effects of vitamin A supplementation
4.1 Research in swine
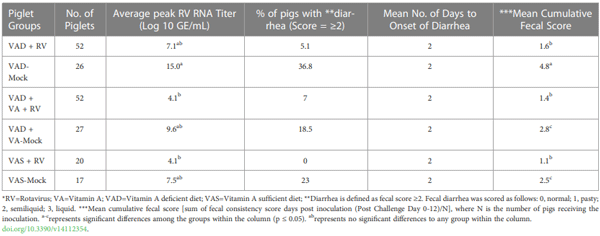
4.2 Research in poultry
4.3 Research in ovo
5 Effect of hypervitaminosis A on immune function
6 The potential applications of vitamin A as an immuno-micronutrient for improving health and preventing diseases
7 Conclusion
Author contributions
Acknowledgments
Conflict of interest
Publisher’s note
Abd El-Wahab, A.,Visscher, C., Ratert, C., Kölln,M., Diephaus, D., Beineke, A., et al. (2017). Outcome of an experimental study in growing turkeys suspected of having a diet related, uncommon and uncoordinated gait. Vet. Sci. 4, 49. doi: 10.3390/vetsci4040049
Ahmad, S. M., Haskell, M. J., Raqib, R., and Stephensen, C. B. (2009). Markers of innate immune function are associated with Vitamin A stores in men. J. Nutr. 139, 377– 385. doi: 10.3945/jn.108.100198
Alagawany, M., Elnesr, S. S., Farag, M. R., Tiwari, R., Yatoo, M. I., Karthik, K., et al. (2020). Nutritional significance of amino acids, vitamins and minerals as nutraceuticals in poultry production and health - a comprehensive review. Vet. Q. 41, 1–29. doi: 10.1080/01652176.2020.1857887
Alizadeh, M., Astill, J., Alqazlan, N., Shojadoost, B., Taha-Abdelaziz, K., Bavananthasivam, J., et al. (2022). In ovo co-administration of Vitamins (A and D) and probiotic lactobacilli modulates immune responses in broiler chickens. Poult. Sci. 101 (4), 101717. doi: 10.1016/j.psj.2022.101717
Al Tanoury, Z., Piskunov, A., and Rochette-Egly, C. (2013). Vitamin A and retinoid signaling: genomic and nongenomic effects. J. Lipid Res. 54, 1761–1775. doi: 10.1194/ jlr.R030833
Amimo, J. O., Michael, H., Chepngeno, J., Raev, S. A., Saif, L. J., and Vlasova, A. N. (2022). Immune impairment associated with Vitamin A deficiency: insights from clinical studies and animal model research. Nutrients 14, 5038. doi: 10.3390/ nu14235038
Arts, R. J. W., Blok, B. A., van Crevel, R., Joosten, L. A. B., Aaby, P., Stabell Benn, C., et al. (2015). Vitamin A induces inhibitory histone methylation modifications and down-regulates trained immunity in human monocytes. J. Leukoc. Bio. 98, 129–36. doi: 10.1189/jlb.6AB0914-416R
Baldwin, T. J., Rood, K. A., Kelly, E. J., and Hall, J. O. (2012). Dermatopathy in juvenile Angus cattle due to Vitamin A deficiency. J. Vet. Diagn. Invest. 24 (4), 763–766. doi: 10.1177/1040638712445767
Bchini, R., Vasiliou, V., Branlant, G., Talfournier, F., and Rahuel-Clermont, S. (2013). Retinoic acid biosynthesis catalyzed by retinal dehydrogenases relies on a ratelimiting conformational transition associated with substrate recognition. Chem. Biol. Interact. 202, 78–84. doi: 10.1016/j.cbi.2012.11.019
Blair, R., Aherne, F. X., and Doige, C. E. (1992). Tolerance of growing pigs for dietary Vitamin A, with special reference to bone integrity. Int. J. Vitamin Res. 62 (2), 130–133.
Blomhoff, R., and Blomhoff, H. K. (2006). Overview of retinoid metabolism and function. J. Neurobiol. 66, 606–630. doi: 10.1002/neu.20242
Bozhkov, A., Ionov, I., Kurhuzova, N., Novikova, A., Katerynych, O., and Akzhyhitov, R. (2021). Vitamin A intake forms resistance to hypervitaminosis a and affects the functional activity of the liver. Clin. Nutr. 41, 82–97. doi: 10.1016/ j.nutos.2021.12.003
Cantorna, M. T., Nashold, F. E., and Hayes, C. E. (1994). In Vitamin A deficiency multiple mechanisms establish a regulatory T helper cell imbalance with excess Th1 and insufficient Th2 function. J. Immunol. 152 (4), 1515–1522. doi: 10.4049/ jimmunol.152.4.1515
Carazo, A., Macákova, K., Matousova ̌ ́, K., Kujovská Krčmová, L., Protti, M., and Mladěnka, P. (2021). Vitamin A update: forms, sources, kinetics, detection, function, deficiency, therapeutic use and toxicity. Nutrients 13, 1–36. doi: 10.3390/nu13051703
Chang, K., Kim, Y., and Saif, L. J. (2012). “Rotavirus and reovirus,” in Diseases of swine, 10th ed. Eds. J. J. Zimmerman, L. A. Karriker, A. Ramirez, K. J. Schwartz and G. W. Stevenson (West Sussex, UK: Wiley-Blackwell), 621–634.
Chattha, K. S., Kandasamy, S., Vlasova, A. N., and Saif, L. J. (2013). Vitamin A deficiency impairs adaptive b and t cell responses to a prototype monovalent attenuated human rotavirus vaccine and virulent human rotavirus challenge in a gnotobiotic piglet model. PloS One 8, e82966. doi: 10.1371/journal.pone.0082966
Chen, F., Jiang, Z., Jiang, S., Li, L., Lin, X., Gou, Z., et al. (2015). Dietary Vitamin A supplementation improved reproductive performance by regulating ovarian expression of hormone receptors, caspase-3 and fas in broiler breeders. Poult. Sci. 95, 30–40. doi: 10.3382/ps/pev305
Chepngeno, J., Amimo, J. O., Michael, H., Jung, K., Raev, S., Lee, M. V., et al. (2022). Rotavirus a inoculation and oral Vitamin A supplementation of Vitamin A deficient pregnant sows enhances maternal adaptive immunity and passive protection of piglets against virulent rotavirus a. Viruses 14, 2354. doi: 10.3390/v14112354
Clagett-Dame, M., and Knutson, D. (2011). Vitamin A in reproduction and development. Nutrients 3 (4), 385–428. doi: 10.3390/nu3040385
Combs, G. F., and McClung, J. P. (2017). “Vitamin A,” in The vitamins. Eds. G. F. Combs and J. P. McClung (London, UK: Academic Press), 110–159.
Cordeiro, T. N., Sibille, N., Germain, P., Barthe, P., Boulahtouf, A., Allemand, F., et al. (2019). Interplay of protein disorder in retinoic acid receptor heterodimer and its corepressor regulates gene expression. Structure 27, 1270–1285. doi: 10.1016/ j.str.2019.05.001
Cortes, P. L., Tiwary, A. K., Puschner, B., Crespo, R. M., Chin, R. P., Bland, M., et al. (2006). Vitamin A deficiency in turkey poults. J. Vet. Diagn. Invest. 18 (5), 489–494. doi: 10.1177/104063870601800514
Coskun, B., Inal, F., Celik, I., Erganis, O., Tiftik, A. M., Kurtoglu, F., et al. (1998). Effects of dietary levels of Vitamin A on the egg yield and immune responses of laying hens. Poult. Sci. 77, 542–546. doi: 10.1093/ps/77.4.542
Cui, D., Moldoveanu, Z., and Stephensenand, C. B. (2000). High-level dietary Vitamin A enhances t-helper type 2 cytokine production and secretory immunoglobulin a response to influenza a virus infection in balb/c mice. J. Nutr. 130 85), 1132–1139. doi: 10.1093/jn/130.5.1132
Dalloul, R. A., Lillehoj, H. S., Shellem, T. A., and Doerr, J. A. (2002). Effect of Vitamin A deficiency on host intestinal immune response to eimeria acervulina in broiler chickens. Poult. Sci. 81 (10), 1509–1515. doi: 10.1093/ps/81.10.1509
Darroch, C. S. (2000). “Vitamin A,” in Swine nutrition. Eds. A. J. Lewis and L. L. Southern (New York: CRC Press), 263–280.
Davis, C. Y., and Sell, J. L. (1983). Effect of all-trans retinol and retinoic acid nutriture on the immune system of chicks. J. Nutr. 113, 1914–1919. doi: 10.1093/jn/113.10.1914
Davis, C., and Sell, J. (1989). Immunoglobulin concentrations in serum and tissues of Vitamin A-deficient broiler chicks after Newcastle disease virus vaccination. Poult. Sci. 68, 136–144. doi: 10.3382/ps.0680136
Dawson, H., Solano-Aguilar, G., Beal, M., Beshah, E., Vangimalla, V., Jones, E., et al. (2009). Localized Th1-, Th2-, T regulatory cell-, and inflammation-associated hepatic and pulmonary immune responses in ascaris suum-infected swine are increased by retinoic acid. Infect. Immun. 77, 2576–2587. doi: 10.1128/IAI.00827-07
Debelo, H., Novotny, J. A., and Ferruzzi, M. G. (2017). Vitamin A. Adv. Nutr. 8, 992– 994. doi: 10.3945/an.116.014720
den Hartog, L. A., and Ravindran, V. (2019). “Foreword,” in Poultry and pig nutrition. challenges of the 21st century. Eds. W. H. Hendriks, M. W. A. Verstegen and L. Babinszky (The Netherlands: Wageningen Academic Publishers), 15–37.
Doeschl-Wilson, A. B., Brindle, W., Emmans, G., and Kyriazakis, I. (2009). Unravelling the relationship between animal growth and immune response during micro-parasitic infections. PloS One 4, e7508. doi: 10.1371/journal.pone.0007508
Duriancik, D. M., Lackey, D. E., and Hoag, K. A. (2010). Vitamin A as a regulator of antigen presenting cells. J. Nutr. 140, 1395–1399. doi: 10.3945/jn.110.124461
EFSA (European Food Safety Authority) (2008). Scientific opinion of the panel on additives and products or substances used in animal feed (FEEDAP) on a request from the European commission on the consequences for the consumer of the use of Vitamin A in animal nutrition. EFSA J. 873, 1–81. doi: 10.2903/j.efsa.2009.873
EFSA (European Food Safety Authority) (2013). Scientific opinion on the safety and efficacy of Vitamin A (Retinyl acetate, retinyl palmitate and retinyl propionate) as a feed additive for all animal species and categories. EFSA J. 11, 3037. doi: 10.2903/ j.efsa.2013.3037
Endo, T., Mikedis, M. M., Nicholls, P. K., Page, D. C., and de Rooij, D. G. (2019). Retinoic acid and germ cell development in the ovary and testis. Biomolecules 9 (12), 775. doi: 10.3390/biom9120775
Faluyi, O. B., and Agbede, J. O. (2017). Dietary Vitamin A supplementary effects on performance and immuno-competence of broiler chickens. Arch. Zootech. 20 (1), 65– 75.
Friedman, A., and Sklan, D. (1989). Impaired T lymphocyte immune response in Vitamin A depleted rats and chicks. Br. J. Nutr. 62, 439–449. doi: 10.1079/ BJN19890044
Green, H. N., and Mellanby, E. (1928). Vitamin A as an anti-infective agent. Brit. Med. J. 2, 691. doi: 10.1136/bmj.2.3537.691
Gudas, L. J., and Wagner, J. A. (2011). Retinoids regulate stem cell differentiation. J. Cell. Physiol. 226 (2), 322–330. doi: 10.1002/jcp.22417
Guo, X., Han, J., Yan, S., Li, Y., Shi, B., and Zhao, Y. (2019). Effects of Vitamin A on growth performance, immunity and antioxidant unction of broilers. Chin. J. Anim. Nutr. 31, 3582–3589. doi: 10.3969/j.issn.1006-67x.2019.08.019
Gürbüz, M., and Aktaç, S. (2022). Understanding the role of Vitamin A and its precursors in the immune system. Nutr. Clin. Metab. 36 (2), 89–98. doi: 10.1016/ j.nupar.2021.10.002
Haaker, M. W., Vaandrager, A. B., and Helms, J. B. (2020). Retinoids in health and disease: a role for hepatic stellate cells in affecting retinoid levels. Biochim. Biophys. Acta 1865 (6), 158674. doi: 10.1016/j.bbalip.2020.158674
Harmon, B. G., Miller, E. R., Hoefer, J. A., Ullrey, D. E., and Leucke, R. W. (1963). Relationship of specific nutrient deficiencies to antibody production in swine. J. Nutr. 79, 263–268. doi: 10.1093/jn/79.3.263
Hoglen, N. C., Abril, E. A., Sauer, J. M., Earnest, D. L., McCuskey, R. S., Lantz, R. C., et al. (1997). Modulation of kupffer cell and peripheral blood monocyte activity by in vivo treatment of rats with all-trans-retinol. Liver 17, 157–165. doi: 10.1111/j.1600- 0676.1997.tb00799.x
Hu, Y., Zhang, L., Zhang, Y., Xiong, H., Wang, F., Wang, Y., et al. (2020). Effects of starch and gelatin encapsulated Vitamin A on growth performance, immune status and antioxidant capacity in weaned piglets. Anim. Nutr. 6, 130–133. doi: 10.1016/ j.aninu.2020.01.005
Huang, Z., Liu, Y., Qi, G., Brand, D., and Zheng, S. G. (2018). Role of Vitamin A in the immune system. J. Clin. Med. 7 (9), 258. doi: 10.3390/jcm7090258
Idi, A., Permin, A., Jensen, S., and Murrell, K. (2007). Effect of a minor Vitamin A deficiency on the course of infection with ascaridia galli (Schrank 1788) and the resistance of chickens. Helminthologia 44, 3–9. doi: 10.2478/s11687-006-0047-4
Jafari, N. V., and Rohn, J. L. (2022). The urothelium: a multi-faceted barrier against a harsh environment. Mucosal. Immunol. 15, 1127–1142. doi: 10.1038/s41385-022-00565-0
Kandasamy, S., Chattha, K. S., Vlasova, A. N., and Saif, L. J. (2014). Prenatal Vitamin A deficiency impairs adaptive immune responses to pentavalent rotavirus vaccine (RotaTeq®) in a neonatal gnotobiotic pig model. Vaccine 32 (7), 816–824. doi: 10.1016/ j.vaccine.2013.12.039
Kelly, M., and von Lintig, J. (2015). STRA6: role in cellular retinol uptake and efflux. Hepatobiliary Surg. Nutr. 4 (4), 229–242. doi: 10.3978/j.issn.2304-3881.2015.01.12
Langel, S. N., Paim, F. C., Alhamo, M. A., Lager, K. M., Vlasova, A. N., and Saif, L. J. (2019). Oral Vitamin A supplementation of porcine epidemic diarrhea virus infected gilts enhances IgA and lactogenic immune protection of nursing piglets. Vet. Res. 50, 101. doi: 10.1186/s13567-019-0719-y
Lauridsen, C. (2019). From oxidative stress to inflammation: redox balance and immune system. Poult. Sci. 98 (10), 4240–4246. doi: 10.3382/ps/pey407
Lauridsen, C., Matte, J. J., Lessard, M., Celi, P., and Litta, G. (2021). Role of vitamins for gastro-intestinal functionality and health of pigs. Anim. Feed Sci. Technol. 273, 114823. doi: 10.1016/j.anifeedsci.2021.114823
le Maire, A., Teyssier, C., Balaguer, P., Bourguet, W., and Germain, P. (2019). Regulation of RXR-RAR heterodimers by RXR- and RAR-specific ligands and their combinations. Cells 8 (11), 1392. doi: 10.3390/cells8111392
Lessard, M., Hutchings, D., and Cave, N. A. (1997). Cell-mediated and humoral immune responses in broiler chickens maintained on diets containing different levels of Vitamin A. Poult. Sci. 76, 1368–1378. doi: 10.1093/ps/76.10.1368
Li, B., Cai, S. Y., and Boyer, J. L. (2021). The role of the retinoid receptor, RAR/RXR heterodimer, in liver physiology. Biochim. Biophys. Acta Mol. Basis Dis. 1867 (5), 166085. doi: 10.1016/j.bbadis.2021.166085
Li, L., Liu, Z., Fang, B., Xu, J., Dong, X., Yang, L., et al. (2022). Effects of Vitamin A and K3 on immune function and intestinal antioxidant capacity of aged laying hens. Rev. Bras. Ciênc. Avic. 24 (4), eRBCA–2021-1572. doi: 10.1590/1806-9061-2021-1572
Lin, H., Wang, L. F., Song, J. L., Xie, Y. M., and Yang, Q. M. (2002). Effect of dietary supplemental levels of Vitamin A on the egg production and immune responses of heat-stressed laying hens. Poult. Sci. 81, 458–465. doi: 10.1093/ps/81.4.458
Lindemann, M. D., Brendemuhl, J. H., Chiba, L. I., Darroch, C. S., Dove, C. R., Estienne, M. J., et al. (2008). A regional evaluation of injections of high levels of Vitamin A on reproductive performance of sows. J. Anim. Sci. 86, 333–338. doi: 10.2527/jas.2007-0153
Machalaba, C. C., Loh, E. H., Daszak, P., and Karesh, W. B. (2015). Emerging diseases from animals. State World 2015, 105–116. doi: 10.5822/978-1-61091-611-0_8
Mahassni, S. H., and Al-Shaikh, N. A. (2013). Effects of Vitamin A overdose on the immune system in rats. Int. J. Pharm. Med. Bio. Sc. 2 (4), 80–91.
Mao, G. E., Collins, M. D., and Derguini, F. (2000). Teratogenicity, tissue distribution, and metabolism of the retro-retinoids, 14-hydroxy-4,14-retro-retinol and anhydroretinol, in the C57BL/6J mouse. Toxicol. Appl. Pharmacol. 163 (1), 38– 49. doi: 10.1006/taap.1999.8828
McDowell, L. R. (2000). “Vitamin A,” in Vitamins in animal and human nutrition. Ed. L. R. McDowell (USA: Iowa State University Press), 5–90. doi: 10.1002/ 9780470376911
McDowell, L. R. (2006). Vitamin nutrition of livestock animals: overview from vitamin discovery to today. Can. J. Anim. Sci. 86, 171–179. doi: 10.4141/A05-057
McGill, J. L., Kelly, S. M., Guerra-Maupome, M., Winkley, E., Henningson, J., Narasimhan, B., et al. (2019). Vitamin A deficiency impairs the immune response to intranasal vaccination and RSV infection in neonatal calves. Sci. Rep. 9 (1), 15157. doi: 10.1038/s41598-019-51684-x
McKenna, N. J. (2012). EMBO retinoids 2011: mechanisms, biology and pathology of signaling by retinoic acid and retinoic acid receptors. Nucl. Recept. Signal. 10, e003. doi: 10.1621/nrs.10003
National Research Council (NRC) (1994). Nutrient requirements of poultry. 9th Edition (Washington DC: National Academy Press).
Niu, X., Ding, Y., Chen, S., Gooneratne, R., and Ju, X. (2022). Effect of immune stress on growth performance and immune functions of livestock: mechanisms and prevention. Anim. (Basel) 12 (7), 909. doi: 10.3390/ani12070909
O’Byrne, S. M., and Blaner, W. S. (2013). Retinol and retinyl esters: biochemistry and physiology. J. Lipid Res. 54 (7), 1731–1743. doi: 10.1194/jlr.R037648
Oliveira, L. M., Teixeira, F. M. E., and Sato, M. N. (2018). Impact of retinoic acid on immune cells and inflammatory diseases. Mediators Inflamm. 2018, 3067126. doi: 10.1155/2018/3067126
Ortega, J. T., and Jastrzebska, B. (2019). The retinoid and non-retinoid ligands of the rod visual g protein-coupled receptor. Int. J. Mol. Sci. 20 (24), 6218. doi: 10.3390/ ijms20246218
Palczewski, K. (2014). Chemistry and biology of the initial steps in vision: the friedenwald lecture. invest. Ophthalmol. Vis. Sci. 55 (10), 6651–6672. doi: 10.1167/ iovs.14-15502
Palludan, B. (1961). The teratogenic effect of Vitamin A deficiency in pigs. Acta Vet. Scand. 2, 32–59. doi: 10.1186/BF03547330
Papanikolaou, Y., and Fulgoni, V. L.III. (2020). Eggs are cost-efficient in delivering several shortfall nutrients in the American diet: a cost-analysis in children and adults. Nutrients 12 (8), 2406. doi: 10.3390/nu12082406
Park, P. S. (2014). Constitutively active rhodopsin and retinal disease. Adv. Pharmacol. 70, 1–36. doi: 10.1016/B978-0-12-417197-8.00001-8
Pino-Lagos, K., Guo, Y., and Noelle, R. J. (2010). Retinoic acid: a key player in immunity. Biofactors 36 (6), 430–436. doi: 10.1002/biof.117
Raghu, P., and Sivakumar, B. (2004). Interactions amongst plasma retinol-binding protein, transthyretin and their ligands: implications in Vitamin A homeostasis and transthyretin amyloidosis. Biochim. Biophys. Acta 1703 (1), 1–9. doi: 10.1016/ j.bbapap.2004.09.023
Raja Kumari Kallam, N., and Sejian, V. (2021). Gut health and immunity in improving poultry production. advances in poultry nutrition research. IntechOpen. doi: 10.5772/intechopen.95989
Reboul, E. (2013). Absorption of Vitamin A and carotenoids by the enterocyte: focus on transport proteins. Nutrients 5 (9), 3563–3581. doi: 10.3390/nu5093563
Riabroy, N., and Tanumihardjo, S. A. (2014). Oral doses of a-retinyl ester track chylomicron uptake and distribution of Vitamin A in a male piglet model for newborn infants. J. Nutr. 144, 1188–1195. doi: 10.3945/jn.114.191668
Rizvi, F., Asghar, K., and Hafeez, M. A. (2003). Effect of Vitamin A deficient diet on immune response in Newcastle disease infected broilers. Pak. J. Life Soc Sci. 1 (1), 13–16.
Romagnani, S. (1999). Th1/Th2 cells. Inflamm. Bowel. Dis. 5 (4), 285–294. doi: 10.1097/00054725-199911000-00009
Sanda, M. E., and Oyewole, B. O. (2015). Effects of Vitamins A and C on performance, immune response and haematology of broilers vaccinated against Newcastle disease. Int. J. Res. Stud. Biosci. 3 (5), 37–42.
Scarola, S. J., Perdomo Trejo, J. R., Granger, M. E., Gerecke, K. M., and Bardi, M. (2019). Immunomodulatory effects of stress and environmental enrichment in long-evans rats (Rattus norvegicus). Comp. Med. 69 (1), 35–47. doi: 10.30802/AALAS-CM-18-000025
Schat, K. A. (2022). The importance of the bursa of fabricius, B cells and T cells for the pathogenesis of marek’s disease: a review. Viruses 14. doi: 10.3390/v14092015
Schuchardt, J. P. (2007). Biological activity of a novel retinoic acid metabolite, s-4- oxo-9-cis-13,14-dihydro-retinoic acid Vol. 152 (Hannover: Gottfried Wilhelm Leibniz Universität). PhD Thesis.
Sepehri Moghaddam, H., and Emadi, M. (2014). The effect of threonine and Vitamin A on immune system in broiler chickens. Int. J. Adv. Biol. Biomed. Res. 2, 756–763.
Shojadoost, B., Alizadeh, M., Taha-Abdelaziz, K., Shoja Doost, J., Astill, J., and Sharif, S. (2021). In ovo inoculation of Vitamin A modulates chicken embryo immune functions. J. Interferon Cytokine Res. 41, 20–28. doi: 10.1089/jir.2020.0212
Sideeg, R. M. (1996). Effect of dietary Vitamin A and nigella sativa on the performance of broiler chicks Vol. 21 (Sudan: University of Khartoum). Master Thesis.
Sijtsma, S. R., Rombout, J. H. W. M., West, C. E., and van der Zijpp, A. J. (1990). Vitamin A deficiency impairs cytotoxic T lymphocyte activity in Newcastle disease virus-infected chickens. Vet. Immunol. Immunopathol. 26 (2), 191–201. doi: 10.1016/ 0165-2427(90)90067-3
Sklan, D., Melamed, D., and Friedman, A. (1994). The effect of varying levels of dietary Vitamin A on immune response in the chick. Poult. Sci. 73, 843–847. doi: 10.3382/ps.0730843
Sklan, D., Melamed, D., and Friedman, A. (1995). The effect of varying levels of Vitamin A on immune response of the turkey. Br. Poult. Sci. 36, 385–392. doi: 10.1080/ 00071669508417785
Smith, N. W., Fletcher, A. J., Hill, J. P., and McNabb, W. C. (2022). Modeling the contribution of meat to global nutrient availability. Front. Nutr. 9. doi: 10.3389/ fnut.2022.766796
Sole, E., González-Prendes, R., Oliinychenko, Y., Tor, M., Ros-Freixedes, R., Estany, J., et al. (2022). Transcriptome shifts triggered by Vitamin A and SCD genotype interaction in duroc pigs. BMC Genomics 23, 16. doi: 10.1186/s12864-021-08244-3
Steinhoff, J. S., Lass, A., and Schupp, M. (2022). Retinoid homeostasis and beyond: how retinol binding protein 4 contributes to health and disease. Nutrients 14 (6), 1236. doi: 10.3390/nu14061236
Stephensen, C. B. (2001). Vitamin A, infection, and immune function. Annu. Rev. Nutr. 21, 167–192. doi: 10.1146/annurev.nutr.21.1.167
Surai, A. P., Surai, P. F., Steinberg, W., Wakeman, W. G., Speake, B. K., and Sparks, N. H. C. (2003). Effect of canthaxanthin content of the maternaldiet on the antioxidant system of the developing chick. Br. Poult. Sci. 44, 612–619. doi: 10.1080/ 00071660310001616200
Thumbi, S. M., Njenga, M. K., Marsh, T. L., Noh, S., Otiang, E., Munyua, P., et al. (2015). Linking human health and livestock health: a "one-health" platform for integrated analysis of human health, livestock health, and economic welfare in livestock dependent communities. PloS One 10 (3), e0120761. doi: 10.1371/ journal.pone
Timoneda, J., Rodrıguez-Ferna ́ ́ndez, L., Zaragozá, R., Marın, M. P., Cabezuelo, M. T., ́ Torres, L., et al. (2018). Vitamin A deficiency and the lung. Nutrients 10 (9), 1132. doi: 10.3390/nu10091132
Villamor, E., and Fawzi, W. (2005). Effects of Vitamin A supplementation on immune responses and correlation with clinical outcomes. Clin. Microbiol. Rev. 18, 446–464. doi: 10.1128/CMR.18.3.446-464.2005
Vlasova, A. N., Chattha, K. S., Kandasamy, S., Siegismund, C. S., and Saif, L. J. (2013). Prenatally acquired Vitamin A deficiency alters innate immune responses to human rotavirus in a gnotobiotic pig model. J. Immunol. 190, 4742–4753. doi: 10.4049/ jimmunol.1203575
Wan, X. L., Zheng, X. C., Liang, J. R., Xiao, X., Yang, H. M., and Wang, Z. Y. (2022). Dietary Vitamin A supplementation improves intestinal morphology and immune performance of goslings. J. Anim. Feed Sci. 31, 217–223. doi: 10.22358/jafs/150174/2022
Wang, Y., Li, L., Gou, Z., Chen, F., Fan, Q., Lin, X., et al. (2020). Effects of maternal and dietary Vitamin A on growth performance, meat quality, antioxidant status, and immune function of offspring broilers. Poult. Sci. 99, 3930–3940. doi: 10.1016/ j.psj.2020.03.044
West, Jr. K.P., Pokhrel, R. P., Katz, J., LeClerq, S. C., Khatry, S. K., Shrestha, S. R., et al. (1991). Efficacy of Vitamin A in reducing preschool child mortality in Nepal. Lancet 338, 67–71. doi: 10.1016/0140-6736(91)90070-6
Yang, H., Liang, J., Dai, H., Wan, X., and Wang, Z. (2020). Effects of Vitamin A supplementation in the diet of breeding geese on offspring intestinal tissue morphology and immune performance. Asian-Australas. J. Anim. Sci. 33 (9), 1463–1469. doi: 10.5713/ajas.19.0890
Yang, Y., Luo, J., Yu, D., Zhang, T., Lin, Q., Li, Q., et al. (2018). Vitamin A promotes leydig cell differentiation via alcohol dehydrogenase. Front. Endocrinol. (Lausanne) 9. doi: 10.3389/fendo.2018.00644
Yee, M. M. F., Chin, K.-Y., Ima-Nirwana, S., and Wong, S. K. (2021). Vitamin A and bone health: a review on current evidence. Molecules 26 (6), 1757. doi: 10.3390/molecules26061757
Yuan, J., Roshdy, A. R., Guo, Y., Wang, Y., and Guo, S. (2014). Effect of dietary Vitamin A on reproductive performance and immune response of broiler breeders. PloS One 9 (8), e105677. doi: 10.1371/journal.pone.0105677
Zhang, L., Hou, Y., Ma, Z., Xie, J., Fan, J., Jiao, Y., et al. (2023). Effect of oral Vitamin A supplementation on host immune response to infectious bronchitis virus infection in SPF chicken. Poult. Sci. https. doi: 10.1016/j.psj.2023.102701
Zheng, Y., Xie, Y., Geldhof, P., Vlaminck, J., Ma, G., Gasser, R. B., et al. (2020). High anti-ascaris seroprevalence in fattening pigs in sichuan, China, calls for improved management strategies. Parasitol. Vectors. 13, 60. doi: 10.1186/s13071-020- 3935-4
Zhou, H. B., Huang, X. Y., Bi, Z., Hu, Y. H., Wang, F. Q., Wang, X. X., et al. (2021). Vitamin A with L-ascorbic acid sodium salt improves the growth performance, immune function and antioxidant capacity of weaned pigs. Animal 15, 1751–7311. doi: 10.1016/j.animal.2020.100133


Many feeds have VitA conntent. How to create a VitA deficiency?, What is the last impaired metabolic reaction involving VitA participation? Should we formulate to that extent? or just for economic productive purposes?






.jpg&w=3840&q=75)