Postweaning multisystemic wasting syndrome (PMWS), also known as porcine circovirus associated disease (PCVAD) in North America, is an emerging disease of swine that was initially identified in herds in Western Canada during the early 1990s.3 Major clinical signs associated with the disease, mainly wasting and respiratory distress, have been described in 6- to 16- week-old pigs.19 The most significant gross lesions of PMWS consist of generalized lymphadenopathy and interstitial pneumonia, although in some cases hepatitis and interstitial nephritis are observed. Lymphocyte depletion and accumulation of histiocytes and/or multinucleate giant cells in lymphoid tissues are the key characteristic histologic features that define the disease; in some cases, intracytoplasmic and intranuclear viral-like inclusions can be found in macrophages.
Porcine circovirus type 2 (PCV2) is considered the essential infectious agent of PMWS,3 and it can be found in virtually all tissues of severely PMWS-affected pigs; the highest amount of viral antigen and nucleic acid, however, is usually in the above-mentioned lymphoid lesions.19PCV2 is a small virus that belongs to the Circoviridae family, genus Circovirus, which has been described ultrastructurally (in tissues, cell cultures, and tissue homogenates) as a 12-23-nm diameter, nonenveloped virus.2,18 Crowther et al.2 have described the viral 3-dimensional features, showing a distinctly polygonal outline and 12 pentamer units containing 60 subunits in total. Pentamer units protrude only slightly, giving an overall diameter of about 20.5 nm.
Ultrastructural findings of lymphoid tissues of PMWS-affected pigs have been poorly investigated and always as complementary data of other studies. Specifically, VLPs have been described within lymphocytes, macrophages, epithelial cells, and hepatocytes;3,7,9 in most of the cases, viral particles were arranged in paracrystalline arrays. Further, intracytoplasmic icosahedral VLPs have been described in apoptotic macrophages. No morphogenesis studies on cell cultures have been carried out with PCV2, and only a single ultrastructural description has been published of PCV1, considered nonpathogenic for swine,1 in which large and small intracytoplasmic inclusions (ICI) were found in persistently infected PK15 cells.22 Therefore, taking into account the limited information regarding ultrastructural observations of PMWS-affected animals, the major objective of the present work was to exhaustively describe cellular and subcellular changes in lymph nodes of naturally diseased pigs.
Materials and Methods
A total of 6 pigs were used in the present study. Four of the pigs were diagnosed as PMWS, based on the individual case definition of the disease fulfilling 3 criteria:20 1) clinical signs mainly characterized by growth retardation, 2) moderate-to-severe lymphocyte depletion and granulomatous inflammation in lymphoid tissues, and 3) moderate-to-high amount of PCV2 in those tissues detected by in situ hybridization.16 The 2 remaining pigs were clinically healthy and had no lymphoid lesions, and they were negative for PCV2 using the mentioned in situ hybridization technique. All pigs were seronegative to porcine reproductive and respiratory syndrome virus (PRRSV) (measured by enzyme-linked immunosorbent assay [ELISA] test) and yielded negative results by means of a PRRSV reversetranscriptase- polymerase chain reaction (RT-PCR) in serum.
Small blocks (1 mm3) of mediastinal and superficial inguinal lymph nodes of all 6 pigs were taken during necropsy and fixed for ultrastructural studies with 2.5% paraformaldehyde (vol/vol) and 2% glutaraldehyde (vol/ vol) in 0.1 M phosphate buffer (PB), pH 7.4, during 24 hours. Subsequently, tissue blocks were washed with PB 0.1 M, pH 7.4, twice and stored in paraformaldehyde 2% (vol/vol) in 0.1 M PB, pH 7.4, at 4uC until further processing. Once histopathologic and in situ hybridization results were available (1 week after necropsy), the above-mentioned lymph nodes of the 6 selected pigs were postfixed with 1% osmium tetroxide at 4uC overnight in 0.5 M sodium acetate, followed by washes with distillated water. After sequential dehydration in acetone, samples were embedded in Eponate 12 resin (Ted Pella, Inc., Redding, CA) and polymerized at 60uC for 48 hours. Semithin, 90-nm-thick sections were obtained with Leica Ultracut UCT Microtome (Leica Microsystems GmbH, Wetzlar, Germany), stained with 1% aqueous toluidine blue solution, and observed with a light microscope to select suitable blocks. Ultrathin sections were cut at 60 to 70 nm thick, placed on electron microscopy (EM) copper grids and, then, contrasted with 2% uranyl acetate and Reynold´s lead citrate solution. Sections were observed with a Jeol 1400 (Jeol LTD, Tokyo, Japan) transmission electron microscope and photographed with Gatan Ultrascan ES1000 CCD camera 2048 3 2048 pixels. Ultrastructural changes were semiquantitatively scored from lack of (score 0) to severe or high amount (score 3). Mediastinal lymph node of pig No. 1 was finally not able to be examined by EM.
Results
Ultrastructural changes of inguinal and mediastinal lymph nodes were noted in PMWS-affected pigs (Table 1). Ultrastructural changes were mainly observed in histiocytes and, to a lesser extent, in dendritic cells, lymphocytes, and endothelial cells. No significant subcellular changes were observed in any control, non- PCV2- nfected pig.
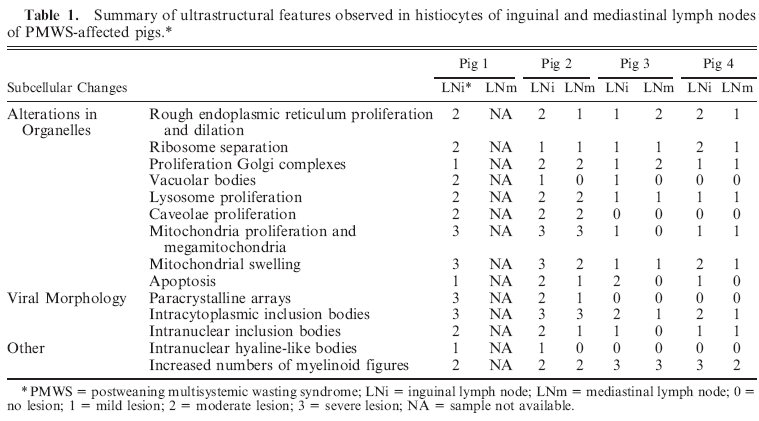
Ultrastructurally, histiocytes showed severe mitochondrial proliferation and enlargement (megamitochondria), with a swollen appearance due to disruption of cisternae and external membrane (Fig. 1a). Additional findings in histiocytic cells included proliferation and dilation of Golgi complex (Fig. 1a), diffuse proliferation and dilation of rough endoplasmic reticulum (RER) (Fig. 1b), and lysosomal proliferation, which was variably associated with evidence of phagocytosis (Fig. 1c). Whorled myelinoid figures were observed in the cytoplasm of histiocytes, most of them forming large and complex electron-dense ICIs and containing aggregates of VLPs. Whorled myelinoid figures were occasionally seen within damaged mitochondria (Fig. 1d). Increased numbers of plasmalemmal vesicles or proliferation of caveolae was observed in histiocytes of inguinal lymph nodes of pigs No. 1 and 2 (Fig. 1e).
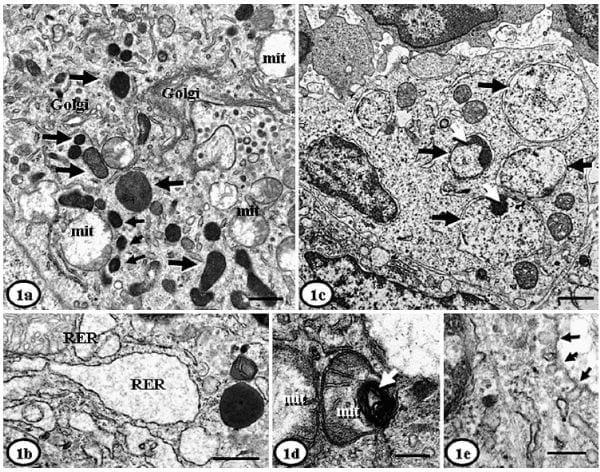
Fig. 1. Lymph node, histiocytes; PMWS-affected pigs. Fig. 1a. Pig No. 1. Intracytoplasmic inclusion bodies (arrows) distributed throughout the cytoplasm, next to enlarged and swollen mitochondria (mit); dilated Golgi complex is also present. Bar 5 1 mm. Fig. 1b. Pig No. 3. Proliferation and dilation of rough endoplasmic reticulum (RER). Bar 5 500 nm. Fig. 1c. Pig No. 1. Nuclei with dilated membranes, and the presence of 4 double-membranebounded giant phagolysosomes within the cytoplasm (black arrows); some of the phagolysosomes have a granular pattern, which corresponds to aggregates of virus-like particle (white arrows). Bar 5 1 mm. Fig. 1d. Pig. No. 1. Two degenerated mitochondria (mit) with disruption of cristae and a myelinoid figure (arrow). Bar 5 200 nm. Fig. 1e. Pig No. 2. Increased number of plasmalemmal vesicles or proliferation of caveolae (arrows). Bar 5 100 nm. Uranyl acetate and Reynold’s lead citrate solution.
ICIs had round, oval, or irregular shapes with variable electron density, and granular or homogeneous appearance (Fig. 2a). They were non-delimited or limited by internal membranes (Fig. 2b) (sometimes with well-defined myelinoid bodies) and were usually dispersed throughout the cytoplasm. ICIs were found in variable number (2-32/cell) and size (0.1-4.5 mm diameter). Small ones (,0.5 mm diameter) were surrounded by a double membrane and almost always contained icosahedral VLPs of approximately 8-17 nm diameter, organized either in paracrystalline arrays (often semicircular); some ICIs had a granular appearance. Large ICIs (.0.5 mm diameter) had a heterogeneous ultrastructural appearance with VLPs organized as a crystalline grille (paracrystalline arrays with a herringbone pattern) (Fig. 2c); some of large ICIs had also a granular appearance, sometimes within whorled myelinoid membranes. ICIs were usually closely related to (sharing the double membrane or in contact with) organelles such as mitochondria, RER, or Golgi complex (Fig. 2d). It was very frequent to find ICIs next to swollen mitochondria that had lysis of matrix, and sometimes disruption of the cisternae and external membrane. Sporadically, small (10-17 nm diameter) icosahedral VLPs were found within damaged mitochondria (Fig. 2e). Some histiocytes also had intranuclear inclusions (INIs), which were less common than ICIs, and were composed of nonmembrane bounded aggregates containing 10- to 13-nm-diameter VLPs (Fig. 2f).
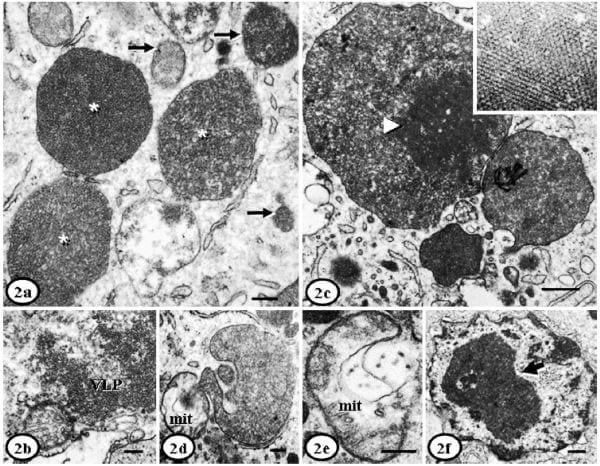
Fig. 2. Lymph node, histiocytes; PMWS-affected pigs. Fig. 2a. Pig. No. 2. Large (asterisks) and small (arrows) intracytoplasmic inclusion bodies containing granular content. The granular content corresponds to 15- to 17-nm diameter icosahedral virus-like particles (VLPs). Bar 5 200 nm. Fig. 2b. Pig No. 3. Intracytoplasmic inclusion bodies with 15-nm diameter icosahedral VLPs, with absence of any external membrane. Bar 5 200 nm. Fig. 2c. Pig No. 1. Complex intracytoplasmic inclusion body with multiple features, including VLPs in paracrystalline arrays, herringbone patterns (arrowhead, insert close-up) and amorphous electron-dense aggregates. Bar 5 200 nm. Fig. 2d. Pig No. 1. Large intracytoplasmic inclusion body with double external membrane and small virions inside (10 nm), next to a swollen mitochondria (mit) also containing VLPs. Bar 5 200 nm. Fig. 2e. Pig No. 1. A swollen mitochondria (mit) that contains 17-nm diameter icosahedral VLPs. Bar 5 100 nm. Fig. 2f. Pig No. 2. A large intranuclear inclusion (arrow) surrounded by clear space and composed of aggregates of indistinct VLPs (10–13 nm diameter). Bar 5 500 nm. Uranyl acetate and Reynold’s lead citrate solution.
Lymphocyte depletion was moderate to severe in all 4 cases. Lymphocytes, dendritic cells, and endothelial cells from PMWS-affected pigs showed mild alterations than histiocytes, and mitochondrial swelling was the most prominent finding. Lymphocytes also showed detachment of ribosomes from the RER membrane and peripheralization of nuclear chromatin. Cells (mainly lymphocytes) with necrotic and/or apoptotic appearance were occasionally found (in PMWS-affected pigs but not in healthy animals) (Fig 3a), and had nuclear shrinkage or peripheral margination of chromatin, enlargement and swelling of mitochondria, myelinoid figures, vacuolar bodies and dilation of RER. Phagocytosis of lymphocytes with giant lysosomes was sporadically observed in histiocytes of Pig No. 1 (Fig. 3b); such histiocytes occasionally contained VLPs. Loss of dendritic processes was a striking finding of dendritic cells. ICIs and VLPs were not observed in lymphocytes or dendritic cells.
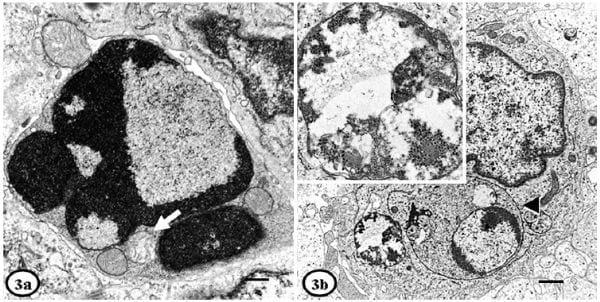
Fig. 3. Lymph node; PMWS-affected pigs. Fig. 3a. Pig No. 1. Apoptotic lymphocyte showing nuclear convolution, margination of chromatin and electron-lucent zones. Mitochondrial swelling (arrow) is observed in the cytoplasm. Bar 5 500 nm. Fig. 3b. Pig No. 1. Phagocytosis of a lymphocyte by a histiocyte (arrowhead), with an adjacent intracytoplasmic inclusion containing VLPs (insert close-up). Bar 5 1 mm. Uranyl acetate and Reynold’s lead citrate solution.
Discussion
This work represents the first extensive study on ultrastructural findings of lymphoid tissues from pigs with PMWS.20 Previous studies used electron microscopy to support histopathologic findings in affected pigs,3,7,9 showing the existence of inclusion bodies that were sometimes composed of paracrystalline arrays of PCV2-like particles. In the present study, the main significant organellar changes associated with viral infection were observed in histiocytes, affecting mitochondria, RER, Golgi complex, and lysosomes. Similar ultrastructural changes were observed in other cell populations, such as lymphocytes and dendritic cells, although to a lesser frequency. As a whole, the observed changes are indicative of secretory activation and inflammation.5 Most of these ultrastructural changes were associated with the presence of VLPs in the cytoplasm of affected cells (and occasionally in the nucleus), which were similar to those observed with other viruses such as African swine fever,5 classical swine fever,6 and rubella virus.
According to the literature, viral factories (VFs) refer to aggregates of viral particles of different shapes and sizes, which represent viral maturation structures that induce changes in cellular organization.12 In the present study, we observed inclusion bodies containing VLPs associated with important alterations in organelles of histiocytes. Therefore, ICIs and INIs can be classified as VFs. In fact, membrane structures of damaged organelles appear to be used to assemble VFs, where immature virions of different sizes (nonenveloped or enveloped by a double membrane) were observed. Therefore, the presence of VFs suggests that viral replication does occur in histiocytes. This result is contradictory with another reports indicating that macrophages do not represent a significant target cell for PCV2 replication in vitro.4 However, other studies have described that at least a certain proportion of macrophages can support virus replication in vitro, perhaps with variability in susceptibility between pigs and also between PCV2 isolates.10 Therefore, the results obtained in the present study support the latter results that PCV2 replicates to a certain extent in macrophages in vivo.
It has been demonstrated that DNA and RNA viruses use RER and Golgi complex for replication. As examples, African swine fever virus (ASFV) capsid assembly and envelopment take place in the RER membrane and cisternae, respectively;5 herpesviruses acquire the secondary envelope and proteins for capsid envelope in the trans-Golgi area;11 and viral assembly of Bunyamwera is produced in the Golgi region in connection to mitochondria.17 In that sense, PCV2 replication is apparently related to proliferation anddilation of RER and Golgi complex, since VLPs were consistently present next to those organelles. PCV2 VFs were also surrounded by proliferated and degenerated mitochondria; this feature has also been described in some DNA viruses such as ASFV, where VFs are observed next to these organelles.5 Of interest, immature PCV2-like particles were also observed within mitochondria, suggesting that the virus might replicate within this organelle. Viral replication within mitochondria is not apparently described in the literature for other viruses; although some RNA viruses can use the mitochondrial membrane for replication, they are not present within the organelle.12 In any case, the potential replication of PCV2 within mitochondria deserves further research.
Previous studies have described PCV2 virions by electron microscopy, revealing small nonenveloped VLPs of 12 to 23 nm diameter in cytoplasm, and 10 to 12 nm diameter in nucleus.3,7,9 The present study further confirms published data, in which icosahedral/granular 12-18 nm diameter VLPs were mainly found in doublemembrane- bounded VFs containing electron-dense material, and icosahedral 15-18 nm diameter VLPs were found in non-membrane-bounded, irregular VFs. Large electron-dense VFs with paracrystalline arrays and whorled myelinoid membranes occupied wide cytoplasmic areas, as it has been described for PCV1 in PK- 5 cells.22 These viral macrostructures probably contain accumulations of large quantities of protein, as it is usually described for viruses in general.12 In contrast, non- embrane-bounded intranuclear VFs were occasionally seen in histiocytes, as it has been also described for other circoviruses including PCV1.21,22 The potential relationship between presence of intranuclear VFs and nuclear changes could not be verified in the present study.
Although apoptosis is considered insignificant as a potential mechanism for lymphocyte depletion in natural occurring PMWS,14 apoptosis of lymphocytes was observed in PMWS-affected pigs but not in healthy ones in the present study. This apparent incongruence may be explained by the different methodologies used to measure apoptosis.14 It has also been suggested that lymphocytes might support PCV2 replication, although to a low degree.13 However, VLPs were not observed in lymphocytes in the present study.
In summary, this is the first detailed study of the ultrastructural findings in lymphoid tissues from PMWS-affected pigs. This work should be further complemented with the development of specific techniques that allow viral detection and localization in cellular organelles of tissues from PMWS-affected animals, and also with studies on morphogenesis of the virus in vitro. The accomplishment of these objectives would allow a better understanding of the pathogenesis of PMWS and PCV2 infection.
Acknowledgements
We are grateful to personnel of Servei de Microscopia from Universitat Auto`noma de Barcelona, especially to Alejandro Sanchez-Chardi, Francisca Cardoso, and Onofre Castell; and Dr. Cristina Risco from Centro Nacional de Biotecnología, Consejo Superior de Investigaciones Científicas, for their excellent technical support. This work was partly funded by the Projects No. QLRT-PL-1999-00307 and 513928 from the Fifth and Sixth Framework Programmes, respectively, of the European Commission and the Consolider Ingenio 2010-PORCIVIR (Spanish government). PhD studies of Carolina Rodriguez Carin˜o are funded by a bursary of Universidad Central de Venezuela.
References
1 Allan GM, Ellis JA: Porcine circoviruses: a review. J Vet Diagn Invest 12:3-14, 2000
2 Crowther RA, Berriman JA, Curran WL, Allan GM, Todd D: Comparison of the structures of three circoviruses: chicken anemia virus, porcine circovirus type 2, and beak and feather disease virus. J Virol 77:13036-13041, 2003
3 Ellis J, Hassard L, Clark E, Harding J, Allan G, Willson P, Strokappe J, Martin K, McNeilly F, Meehan B, Todd D, Haines D: Isolation of circovirus from lesions of pigs with postweaning multisystemic wasting syndrome. Can Vet J 39:44-51, 1998
4 Gilpin DF, McCullough K, Meehan BM, McNeilly F, McNair I, Stevenson LS, Foster JC, Ellis JA, Krakowka S, Adair BM, Allan GM: In vitro studies on the infection and replication of porcine circovirus type 2 in cells of the porcine immune system. Vet Immunol Immunopathol 94:149-161, 2003
5 Gomez-Villamandos JC, Bautista MJ, Carrasco L, Caballero MJ, Hervas J, Villeda CJ, Wilkinson PJ, Sierra MA: African swine fever virus infection of bone marrow: lesions and pathogenesis. Vet Pathol 34:97-107, 1997
6 Gomez-Villamandos JC, Salguero FJ, Ruiz-Villamor E, Sanchez-Cordon PJ, Bautista MJ, Sierra MA: Classical swine fever: pathology of bone marrow. Vet Pathol 40:157-163, 2003
7 Hirai T, Nunoya T, Ihara T, Kusanagi K, Kato T, Shibuya K: Acute hepatitis in a piglet experimentally inoculated with tissue homogenates from pigs with postweaning multisystemic wasting syndrome. J Vet Med Sci 65:1041-1045, 2003
8 KiupelM, StevensonGW,Choi J, Latimer KS,Kanitz CL, MittalSK: Viral replication and lesions inBALB/c mice experimentally inoculated with porcine circovirus isolated from a pig with postweaning multisystemic wasting disease. Vet Pathol 38:74-82, 2001
9 Kiupel M, Stevenson GW, Mittal SK, Clark EG, Haines DM: Circovirus-like viral associated disease in weaned pigs in Indiana. Vet Pathol 35:303-307, 1998
10 Mankertz A, Domingo M, Folch JM, LeCann P, Jestin A, Segales J, Chmielewicz B, Plana-Duran J, Soike D: Characterisation of PCV-2 isolates from Spain, Germany and France. Virus Res 66:65-77, 2000
11 Mettenleiter TC: Herpesvirus assembly and egress. J Virol 76:1537-1547, 2002
This article was originally published at Veterinary Pathology (VET) Journal, 2009 46: 729 DOI: 10.1354/vp.08-VP-0141-R-FL. Engormix.com thanks the authors and the journal for this contribution.