Specific Causes of Reproductive Failure
Fertility in cattle has many pre-requisites and components, which require males and females to be functionally capable of reaching the last step, the birth of a normal, vital calf, thus defining the general breeding goal: cows should return to normal ciclicity early postpartum, show strong and regular estrous signs, conceive after AI, carry their pregnancy to term, calve easily and give birth to viable and healthy calves. Low calving rates relate to the inability of the cow to resume ovarian activity and thus failure to cycle and to express estrous signs (anovulatory and behavioral anestrus, irregular estrous cyclicity, etc); as well as to reproductive wastage due to fertilization failure, early and late embryonic mortality, fetal mortality (abortions) and stillbirths. Fertilization rates (% of ova being fertilized), are generally high after AI under controlled conditions. However, decreasing rates are now seen in high-producing Holstein cows. There are probably many causes why 10 - 25% of ova are not fertilized after AI, and they can be of either male or female origin. But, to what extent does husbandry incide?
Sire fertility relates both to sperm numbers and their quality at the site of fertilization in the oviduct. The male not only contributes to fertilization in the female but also, through the quality of spermatozoa, to the ability of the embryo to survive throughout pregnancy. However, many details concerning the structure and function of the spermatozoon are yet poorly understood. Spermatozoa are transcriptionally silent as a consequence of the highly condensed chromatin architecture and there is almost no cytoplasm capable of supporting translation. However, at least in primates and rodents, spermatozoa carry a full complement of mRNAs that can, under certain conditions, be translated de novo. The spermatozoa delivered by fertile males contain up to 3,000 different mRNA species and regulatory micro-RNAs and non-coding RNAs. The functionality of these RNA molecules in fertilized embryos is largely unknown. Only some of these mRNAs have shown to be important through their established roles during development. Many of these mRNAs are male-specific, potentially affecting phenotypic traits in the offspring. This epigenetic phenomenon can involve the transmission of extra-chromosomal episomal elements. Recent studies have revealed that the presence of aberrant RNA in defective spermatozoa might influence and even disrupt early embryogenesis but there is no information whether such aberrant RNA would cause epigenetic-like defects, visible later during fetal development. Differences in individual transcriptome profiles in AI-bull sires have been recently shown, indicating a relation between presence of specific transcripts relevant for cell functions and the level of fertility after AI. Determination of the sperm transcriptome is thus likely to increase our understanding of reproductive success (and failure), provided studies are done on proven AI-bulls, where all other possible confounding factors are minimized or, at least, controlled. Since the spermatozoon is a particular cell type, conventional microarrays do not presently contain all their transcripts, particularly those low abundant. Major attention must, therefore, be put on adaptations of current extractions protocols for spermatozoa.
Female fertility is influenced by numerous factors (see Table 4 and Fig. 21). Establishment of lactation postpartum leads to the inhibition of LH pulsatility and a subsequent normal anovulatory anestrus (more dramatic when suckling is present). In suckling and low-milk yield cows, this period is followed by a resumption of gonadotrophic and ovarian activity and, most often, normal fertility. High-producing dairy cows, however, have often extended periods of anovulatory anestrus, as a consequence of the inadequate hormonal balance that the catabolic NEB causes (low blood levels of LH, insulin and IGF-I), resulting in impaired follicle development, estrous signs, LH surge and ovulation. Moreover, high-producing cows in NEB experience a delayed onset of postpartum ovarian activity and reduced P4 levels, the latter caused by a lack of ovarian luteinized tissue or a higher rate of metabolism of the hormone by increased feed intake.
Figure 21. Primary, non-infectious, causes for reproductive failure in high-producing dairy cattle
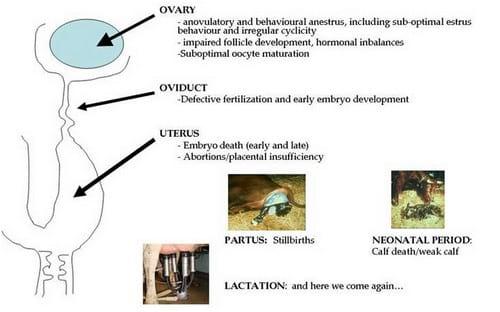
High producing cows with these hormonal imbalances have shorter estrous cycles and depict fewer estrus signs than expected (Table 1,Fig. 12), owing to sub-optimal estradiol levels. Hormonal imbalance (LH levels, metabolic rates), as well as genetic factors relate to the occurrence of twinning and cystic ovarian disease in high milk yield cows. Oocyte quality, built upon a total maturation time in the ovary of around 3 months, is very sensitive to negative influences such as nutritional deficiencies or over-conditioning (Table 4). Accumulation of NEFA derived from the adipose tissue during NEB in the follicle fluid constrains the proliferation and health of the granulosa cells and thus jeopardizes oocyte development. Extension of the pre-ovulatory phase, i.e. delayed ovulation, due to suprabasal P4-levels causes ageing processes in the oocyte and compromises fertility, leading to repeat breeding by fertilization failure. After fertilization, an embryo is formed which develops in the oviduct during the first 3 - 4 days before it enters the uterus. The embryonic period lasts up to day 42 after fertilization and involves a series of critical periods, of which one comprises the first three weeks of development ("early embryonic death"), accounting for ~20% of the total losses. From this moment up to day 42, losses are termed "late embryonic death". While fertilization failure and early embryonic death relate to a low genetic index of the female, up to 25% of late embryonic deaths have been seen in cows with genetic potential for a high milk yield but mainly related to milk yield rather than to their genetic index. Some of the early embryo losses might result from a malfunctional cytoplasm which impairs further development of the fertilized oocyte, a situation seen in over-conditioned repeat breeders. Cytokines can adversely affect uterine function and indirectly cause early embryonic death in relation to mastitis during early lactation.
Early embryonic death can also result from a sub-optimal combination of genes arisen during fertilization. Genetic selection for milk production has also been related to early embryonic death owing to impaired oocyte quality, lowered P4 concentrations, or both. As well, reduced levels of P4 and of insulin-like growth factors can induce a sub-optimal uterine microenvironment, incapable of sustaining early embryonic life. An early loss will lead to a new estrus (within 17 - 24 days from AI) giving the dam a new chance to conceive by minimal costs in time. However, there might be other reasons behind embryonic losses, even at a later stage, resulting in a costly delayed return to estrus. The early establishment of a pregnancy is controlled by complicated endocrine events, including the regulation of the expression of interferon-tau (IFTN), the maternal pregnancy recognition factor in the bovine. Owing to its role in regulating IFTN expression, the Fibroblast growth factor-2 (FGF2) gene relates to early embryonic death. A hormonal asynchrony causes early embryonic death, and might be linked to the general trend of decreased fertility where lower levels and atypical temporal release (atypical profiles) of serum P4 have been detected. It is possible that high-producing cows have higher metabolic rates and a larger blood flow through the liver, which leads to lower concentrations of P4 and estradiol reaching the target cells. Even husbandry-driven stress is nowadays regarded as a major contributor to endocrine disorders, causing repeat breeding and early embryonic death. Excess rumen-degradable protein has been associated with increased embryonic mortality and, possibly, fertilization failure, because an excess of protein and a deficit of energy in the feed ration increases the production of ammonia that, when converted into urea in the liver, causes embryo mortality through an exacerbation of NEB and reduced plasma P4-levels, an alteration of uterine pH and increased secretion of PGF2α. During the remaining gestation (day 43 to term, called fetal period), losses such as abortions are less prevalent, but an increased incidence (>12%) of early fetal loss (between 45 and 60 days of gestation has been recorded and related to current intensive management systems, hot climate, and to animal factors of non-infectious nature, including twinning, and sire (often related to infections or carried genes). Plasma concentrations of pregnancy-associated glycoproteins (PAG) on day 35 of gestation can also indicate subsequent fetal loss with a strong relation to sire.
The epigenetic programming of the gamete genome is crucial for normal development and its modification influences heritable patterns of chromatin conformations and gene expression, leading to stable phenotypic differences among specialized cell types, as for instance those of the placenta. Some syndromes in cattle such as the "large offspring syndrome" observed in in vitro produced animals or in cloned animals, have been considered the consequence of existing epigenetic errors in the donor cell genome or of an incomplete epigenetic reprogramming during development. Particular placentopathies in cloned founder cows reflect epigenetic effects, but there is yet much to unravel in order to define the molecular mechanisms involved and when they arise. For instance, whether aberrant paternal mRNA would cause epigenetic-like defects, visible either during early embryo development or later during fetal development, is yet to be determined, although seemingly related to the fertility of the males.
Calves born dead or dying within 24 hours are termed stillborn. Stillbirth is costly, not only because of the loss of the calf, but also because it leads to a higher risk for diseases in the dam and culling. In Sweden, stillbirth rates are significantly higher in SH-heifers (first calvers, ~11 %) compared with SH cows and SR heifers or cows (6%), a difference that has increased during the last 20 years. Noteworthy, for second calvers, hardly any differences exist in either calving difficulty or stillbirth between breeds. Only half of the stillbirths are associated with calving difficulties indicating low fetal or calf vitality as one reason. Although it is a multi-factorial problem, the genetic factor is significant, as shown by an association between a higher proportion of Holstein genes and increased stillbirth rates, present already by the 1990´s. Such a relationship was not present in Scandinavian Red breeds. Being the 1st-parity animals overrepresented, this category should be the main source of information for further genetic evaluation.
Calf survival is also multi-factorial, including placental insufficiency, prolonged parturition, poor- or abnormal maternal behaviour, bad udder conformation, etc. Although the physiological cause behind the lower vitality of the fetus or calf is unknown, placental insufficiency is related to lower levels of placenta-specific hormones measured during the last six weeks of gestation in heifers with stillborn calves. Whether such placental insufficiency relates to epigenesis, to hormonal imbalance under production or husbandry constraints, to maternal behaviour or the concerted interplay of these factors, remains to be determined. All these maternal factors can extend the interval from birth until 1st suckling, a simple indicator for low calf vitality. Group-housing during calving might also cause delayed 1st suckling due to disturbing older or higher-ranked cows with a strong maternal interest in newborn calves.
Strategies to Ensure Good Fertility in High-producing Dairy Cattle
As already mentioned, infertility in dairy cattle is multifaceted. Addressing it requires, therefore, a multi-disciplinary approach. However, not all solutions intended for amelioration of the problem are long-lasting, feasible or acceptable. Some measures can be applied rather immediately while others require further research and long-term strategies. From an EU perspective, discussions about animal ethics and welfare demand higher longevity of dairy cows and bans on the use of drugs for improving reproduction, especially hormones. Emphasis is needed on the relation between management and genetic gains, considering that many of the current problems with dairy cow fertility are a logical consequence of the low profit margins of the dairy sector, which impinges for high milk yields. With a global trend of strict cost controls, increased herd sizes, and changing farming systems, there is an unfortunate association with shortages of skilled labour and less time to look for physiological signals in the herd, the basis for good management. Moreover, we should recognize our limited ability to prevent and treat diseases, to appropriately manage, feed and select dairy cows with desirable reproductive traits. Excellent papers on strategies for increasing fertility in dairy cows have been written. Moreover, a summary of strategies proposed for restoring fertility in high-producing dairy cows is hereby presented in Table 5.
Table 5. Strategies to ensure high fertility among high-producing dairy cattle, pros and cons
Short- and Medium-term Strategies to Ameliorate Fertility Problems
Manipulation of the Oestrous Cycle and Control of Ovulation
From a veterinary medicine perspective, manipulation of the estrous cycle and the control of ovulation appears as a good short-term strategy. Application of methods to control the development of follicle growth, the promotion of ovulation in anestrus cows, the regression of the corpus luteum in cyclic cows and the synchronization of estrus and ovulation at the end of treatment, before AI (on spontaneous or expected estrus) or mating have been thoroughly studied. However, most of these approaches imply the use of hormones (estradiol, P4, GnRH, PGF2α) administered by different treatment regimes and with different intensities. Because there is large variation between countries regarding the availability and regulations of these treatments, it is difficult to consider this alternative as a feasible general strategy. A similar situation applies to the use of exogenous recombinant bovine somatotropin (rbST) around AI or of exogenous P4 after ovulation, in order to increase expected low levels of IGF-I or P4, respectively. Both hormones are banned already, or the public concern prevent their application. The need for multiple applications and the presence of a veterinarian (sometimes enforced by law) as well as the treatment costs, counteract their use. Moreover, we still do not know of eventual long-term effects of hormonal manipulations on genetics, thus shadowing our possibilities to safely select the best cows. Either way, these are no sustainable alternatives.
Dietary Management
Use of diets designed to improve fertility by counteracting specific points in relation to NEB has always been an attractive way to circumvent the impairment of reproduction during early lactation that the partitioning of energy causes. However, the cow is biologically driven to mobilize body fat when she is fatter than her biological target condition, which makes this strategy difficult. Several ways have been attempted to reduce the effect of NEB, namely
- The reduction of BC at calving, to avoid the limiting negative feedback effect of body fat on the cow´s DMI (the procedure require high-quality diets are available for these -hopefully- "thinner" cows
- The feeding of low-protein diets that reduce body-fat mobilization by provoked imbalance in the protein-to-energy ratio
- The change of carbohydrate source in the diet to increase dietary energy concentration (increasing starch or fat and decreasing forage content, a risky procedure because of its implications on rumen function, milk composition, nutrient partition and metabolic hormones
- The inhibition of milk fat synthesis with exogenous conjugated linoleic acid (trans-10, cis-12 CLA), thus limiting energy output in milk
Moreover, although there are specific nutrients designed to trigger the endocrine system of the cow during early lactation; one must consider that while insulin (as well as GH, IGF, LH and leptin) can profoundly influence ovarian function (follicular development in particular), the roles of GH, IGF-I and leptin appear to be more related to milk yield, BC and LW than with the whole nutritional status. Changes in diet composition can, on the other hand, elicit large changes in insulin levels, since plasma insulin concentrations are positively related to dietary starch concentration. Therefore, attempts have been made to use exogenous propylene glycol or hyperinsulinemic diets with the purpose of "fooling" the cow into a virtual anabolic condition, increasing glucose and insulin concentrations in circulating blood. However, insulin (directly or via glycogenic substance) has differential effects, being able to stimulate resumption of cyclicity but also cause detrimental effects on oocyte competence. Moreover, it has recently been shown that insulin alters the enzymes responsible for the catabolism of P4 in the liver, thus decreasing the decay of the hormone. A recent review on the relation between nutrition of high-yield dairy cows and oocyte and embryo quality has been published.
On the other hand, exogenous conjugated linoleic acid (CLA) seems to exert positive effects on oocytes, and to increase P4 and PGF2α levels. Combinations of these designed diets are obviously more attractive, because they have proven to increase pregnancy rates dramatically (from 27 to 60%), implying that cows producing ~10,000 kg would have a fertility comparable to cows producing ~6,000 kg, by increasing insulin status immediately postpartum, and then reducing insulin status during the mating period.
Managing the Dry Period
Shorthening or eliminating the dry period has been postulated as a suitable way to quickly enhance fertility in dairy cows. This management can increase DMI during the transition period, decrease milk energy output, or both. By increasing the energy status of dairy cows, there is an indirect increase in reproductive efficiency. However, such practice does not apply in general as it may have negative effects on udder health and total milk yield, and should be considered on a herd-to-herd basis.
Extended Lactation and Differential Milking
Most veterinary attention in dairy cattle is required from one week before to 10 weeks after calving, confirming that calving is a welfare risk. Moreover, it is important to avoid the impact of NEB on the resumption of reproductive function. For these reasons, voluntarily delaying the 1st postpartum AI, and attempting to have the cow calving at a calving interval of maybe 18 months or so, leads to (a) the cow prolonging its lactation (so-called "persistent" lactation), (b) no need to look for estrus during the milk peak at early lactation, and (c) with AI done later during lactation, the cow having a better chance of getting pregnant. On the other hand, because different cows have individual lactation curves, it seems difficult to select those individuals which are to be grouped for short respectively long calving intervals. Increasing milking frequency by the use voluntary milking (using robotics) promotes extended lactation, but leads to a delay in the appearance of ovulatory estrus.
Moreover, using extended calving intervals means fewer calves are born per cow, persistent lactation can lead to udder health problems in cows with high somatic counts and milk production can only be reasonably maintained by compensatory feeding during the declining phase. To be economically acceptable for the farmer, the milk yield has to be maintained over time and thus requires proper management. An alternative strategy is to flatten the peak of the milk curve by employing once-daily milking in early lactation (but risking udder problems…). Such practice promotes earlier resumption of ovarian cyclicity by increasing nutritional status, avoiding the impact of NEB and, further, the cow maintaining BC throughout lactation. It may also be an alternative to genetically select for flatter lactation curves.
Use of High-fertility Bulls
Use of AI with semen from sires with proven high-fertility is probably the most obvious and simple recommendation. However, in order to prolong the term of this strategy from short to medium, the breeding selection must be appropriate, i.e., including fertility traits with a certain weight in order to warrant that the improvement of reproductive performance is selected while still maintaining enough yearly increase in milk yield, thus warranting the rentability of the dairy production in a longer perspective.
Crossbreeding
Use of semen from other breeds where the decline in fertility is not a severe problem is also a medium-range alternative to halt fertility deterioration, although it might not be the best long-term strategy. The procedure is already customary in New Zealand where Jersey is crossed with Holstein, and has proven attractive for other markets, provided the semen comes from appropriately progeny-tested sires, and with production levels close to the breed in question. A valid example is the use of SR semen for crossbreeding on Holstein, now attaining production levels quite similar to those pure Holsteins, but with superior fertility. This strategy can also be used to combat inbreeding in herds where the problem is large. In particular, the development of multiple lines with similar capacity for milk production is attractive, based on the assumption that crossbreeding could be used to capitalize on heterosis. However, we should always bear in mind that cross-breeding is not per se genetic improvement and that genetic selection is still needed within the breeds used.
Full-purpose Buildings, Management and Automated Systems
Well-designed cow barns, with good width and slip resistance of the flooring of alleys, passageways and holding areas suport best animal well-being and allow for efficient health management, including reproduction, by promoting expression of behavioural estrus, and providing the best opportunities for estrus detection by the staff. However, detection of estrus or of health problems (lameness, for instance) is often constrained by the shortage of skilled and experienced personnel to spend enough time with the animals. This problem is, unfortunately, aggravated by the increasing display of weaker signs of oestrus by high-producing cows, which are not always even detected by the use of mounting-detector. For these reasons, automated systems or the supervision have been designed, including those monitoring the activity by the cows (as ALPROtm or Afiacttm, among others available in the market) with heritabilities of ~0.17 for estrus control.
Other methods relate to specific hormones. For instance, P4 concentrations in blood or milk can be used to monitor the interval from calving to initiation of luteal activity, with a heritability of ~0.2 . Owing to the close relationship between P4 levels, peak milk production and ML of the dairy cow, P4 profiles appear suitable to evaluate fertility values of individual cows even at infrequent intervals (e.g., once monthly). Different fertility variables can then be derived from this P4-profile, as the interval from calving to initiation of luteal activity, the interval from calving-to-ovulatory-oestrus, etc, variables that are less affected by management decisions and more influenced by the cows' own physiology. Perhaps more importantly, P4 levels in milk can today be determined on-line, together with other indicators for presence of preclinical mastitis (lactate dehydrogenase LDH), hepatic function (beta-hydroxybutyrate, BHB) and of intake protein balance (urea), using automated equipments (such as Herd-navigator). This on-line detection allows the diagnosis of problems and help improving the management of the high-producing cow at an individual basis. However, these indicators can only aid solving the individual problems that appear. Unless they are used to perform proper selection of the animals, the equipment will simply be another diagnostic tool (albeit economical for large exploitations). However, if linked to activity recorders, they may improve our capability to better time estrus and thus lead to a better timing of AI. Other useable markers are easier, such as the BCS, but there are no automated systems yet available for this indicator.
Long-term Strategies to Reach a Sustainable Improvement of Dairy Cow Fertility
Correction for the Weight of Different Functional Traits in the Breeding Goal
Following the Scandinavian decision to include fertility traits in the breeding goal for selection of dairy cattle (31), many countries are now incorporating similar approaches in their selection strategies, but the relative weight of health and reproduction still varies largely as indices of breeding selection (e.g., from 0.03 to >30%). As already mentioned, reproductive traits have low heritabilities, but their genetic variation is large, thus making it possible to genetically select for good fertility without constraining the gains in milk yield. Thus, a higher weight for breeding values of several functional traits such as fertility, claw health, longevity and milk yield have to be applied. Over many years, this has been proven in Scandinavia, where adjustments of the weight of different functional traits can provide more appropriate breeding indices for selection and even increase farmer´s profits. These data suggest that the current situation with dairy reproduction genetics is not a cul-de-sac. We need simply to work towards a lower progress regarding milk yield but with major gains in cow health. Economic calculations can be done for each of the weightings so that farmers and breeding organizations can best argument for the value of the changes towards sustainability, thus avoiding unnecessary, short-sighted discussions. Moreover, we should perhaps re-consider the use of the trait measurements we have used for more than 50 years! The genetic accounting systems (as Best Linear Unbiased Prediction, BLUP; Animal Model, AM; Test Day Model, TDM etc) might not fully compensate for the recent dramatic changes we have experienced in environmental requirements and management practices. Perhaps we should consider the replacement of "non-return rate (NRR)" by "pregnancy rate (PR)" when basically all high-producing cows are pregnancy-tested using various means (trans-rectal palpation, ultrasound, pregnancy-specific protein determinations, etc). Such considerations seem imperative in the face or our inability to accurately detect estrous signs.
We should also strive to develop accurate and practical methods and determine best markers for the complex physiological traits that relate to fertility constraints, to measure welfare traits under different environmental conditions and incorporate them into the breeding goals and selection schemes. As an example, we need to determine the strength of display of estrus signs, when conceptus losses occur in relation to characteristics such as breed, milk yield, production-related pathologies or husbandry. Without these markers, our capacity to design proper strategies is undermined. Markers must also be practical in order to ensure commercial use. Use of on-line measured indicators (see above point vii) can, provided that the monitoring can be done at low cost and the system widely adapted for most on-farm use, lead to information be easily incorporated into the selection schemes for bull sires. In any case, the most important trait measurements should e based on classical physiological data such as P4 levels, estrous activity, BCS, but also on novel ones such as measurements of the strength of estrous signaling (rather than the classical standing or mounting marks), or the steroid clearance in the liver (with cows selected for low steroid hepatic clearance rate). In this respect, we also should consider to strengthen the selection weight for the timing of the 1st ovulation postpartum, the earlier the better, as it is performed in other species. Indicators for this trait are available, such as P4 levels (see above). To optimize fertility we also need to consider the quality of the bull semen with regards to RNA content as described above.
As mentioned already, another potentially useable marker is the BCS, a moderately heritable (0.09 - 0.45) trait favourably related to fertility and survival. Selection for milk yield increases the NEB and lowers the BC of high-producing dairy cows, particularly when these animals have also been selected for "angularity" (dairy phenotype, see Fig. 11), a trait that has a strong genetic relation with BC. Because there are positive genetic associations between BC and reproductive performance, selection programs based on mid-lactation BCS (when the genetic variance for BCS is largest and the genetic correlations between BCS and fertility strongest). The BC is being used as a predictor trait for genetic merit for fertility in The Netherlands as well as in Ireland and the UK. Increasing the BC of dairy cows by selection would perhaps increase their genetically-determined set point for BC during lactation and thus diminish the effects of NEB, if present. Such thoughts have lead to the "re-development" of other concepts regarding the need for appropriate selection of dairy cows, such as the concept of developmental programming, which basically implies that the plane of maternal nutrition can have an impact on the reproductive function of the descendant and its fertility. To this end, our current use of traditional breeding value estimation procedures, and ot multitrait selection based on large databases and biobanks, might need from the inclusion of other, more suitable markers, to provide further gain in genetic evaluations, where traits such as milk yield, calving interval and survival can be combined to provide selection for longevity while maintaining acceptable levels of milk production. However, these methods are still too costly and too slow to resolve the problems we face today.
Prospective Genetic Selection Strategies
Over the past decade, whole-genome scanning using microsatellite markers on specific experimental designs, has led to the identification of several quantitative trait loci (QTL) for characteristics related to health, fertility and production, including single trait QTLs for the maternal (chromosomes 18, X/Y) and paternal (chromosomes 10, 18) effect on NRR. Use of QTL in selection is most beneficial for low heritability traits, sex-linked traits and traits expressed late in life, such as daughter fertility, a commonly used breeding value in selection (in Sweden, since he early 1970s). Multiple trait QTL regions have also been found for the combination of non-return rate and udder characteristics, and between production traits and non-return rate. Linkage to genetic markers have also included ovulation rate and multiple ovulations, associated with QTL on chromosome 7, a chromosome where many genes linked to endocrine and fertility aspects of dairy cows have been identified. However, because the region of interest as derived from a QTL scan is often quite large and can contain several hundred genes, there is a need for the simultaneous profiling of gene expression of many genes by RNA expression arrays. One of the available strategies is the candidate gene approach, where physiological findings are used to identify genes whose variation can have influence the trait of interest. The genetic variation between animals in a particular gene (such as that coding for GnRH or the bLH receptor) is, at the end, linked to the phenotypic information.
However, we must bear in mind that fertility is one of the most difficult and complex traits owing to low heritabilities caused by the polygenic nature of reproductive traits and the strong environmental influences on reproduction and the long generation intervals in the bovine. In fact, thus far, no gene with a causative mutation has been identified, which underlies a detected QTL effect concerning reproductive traits in cows. For bovine fertility, many genes account only for a small amount of the phenotypic variation, the rest being caused by the environment. This is probably why, despite several identified QTLs have been incorporated into selection strategies by breeding companies globally, the rate of success is low, or very low (rev by). Moreover, the identification of the variation in the genes involved is very costly, mostly because it is time-consuming.
The DNA microarray technology, which includes various versions of bovine microarrays, provides a tool for the analysis of tens of thousands of genes simultaneously, so that transcriptional and genomic changes can be identified on a global scale. To date, public databases comprise expressed genes from a multitude of bovine tissues, including embryos. Gene expression profiling using this technology for functional genomics is expected to aid the identification of genes and/or gene networks, that can be best linked to fertility and its evident polygenic trait character, thus allowing for genomic selection. Studies on epigenetics, using high-throughput methods for analyses of DNA cytosine methylation patterns are likely to provide support for specific areas relevant for fertility, such as early embryonic development and placenta formation and function, where maternal and paternal genome components interact.
Most likely, advances in the recording of Single Nucleotide Polymorphisms (SNPs) are being made, particularly with regard to the speed of genotyping individual animals for many tens of thousands of SNPs, thus opening for the inclusion of genome-wide marker information in the prediction of breeding values. Genomic selection of young bulls is ongoing in The Netherlands, Norway and Denmark/Sweden. Genome-wide association analysis of tissue samples using whole-genome SNP-arrays (such as the BovineSNP50 BeadChip, Illumina Inc, USA) would provide information regarding the most important genetic factors that influence fertility and reproduction. Such genomic selection predicts breeding values for a large number of those haplotypes across the entire genome that are derived from combinations of marker alleles, thus increasing the degree of security for breeding values by 20-30%, compared to today´s methodology. Selection programs based on such information should improve reproductive success in the future, considering more information shall be gathered for attributes with low hereditability such as fertility. The most obvious application for this yet developing technology focus on dairy calves (which obviously do not have any phenotypic records yet), so that conventional progeny testing can, ultimately, be virtually waived, with enormous cost savings. However, although these emerging technologies are yet to be fully established and cross-checked with the evolving traditional breeding estimations, so that the risk of late detection of undesired side-effects of selection are minimized, they promise to have a large impact on our understanding about the genetic factors controlling reproductive success.
Concluding Remarks
The current situation with dairy reproduction genetics is not a cul-de-sac. However, we need to work towards a more balanced progress regarding milk yield and cow health, by either a direct selection for strategic fertility traits or by indirectly selecting for longevity or body condition score. The success shown by the Nordic countries in keeping a largely unchanged genetic trend in female fertility and calving traits while increasing genetic gains in production has shown this is possible. Either way, it requires using appropriate weightings of the available breeding values in traditional breeding analyses systems, or a combination of these traditional systems with novel developments in genome-mapping and functional genomics, pertaining genomic selection. However, because this long-term, hopefully permanent solution will take time to become effective, in the mid-term, we need to gain a greater understanding of the interactions between nutrition, management and fertility to better manage the current negative trends in dairy cattle health and welfare. More awareness of barn constructions to help appropriate display and detection of estrous signs, as well as better flooring, more and safer (less stressful) space for animal-animal and animal-man interaction are pending solutions. Application of specially designed diets, modification of body condition and milking profiles, and better use of markers (behaviour, P4-profiles, estrus selection tools, etc.) are mandatory. Increasing our yet fragmentary knowledge on reproductive losses before calving is essential both in terms of the period of occurrence (embryonic respectively fetal), and the relationship between sire influence (potential epigenetic effects), embryo potential, and placental health, the latter determining the presence of stillbirth or the birth of weak calves. Last but not least, use of appropriate sires for breeding remains the best option for genetic improvement of fertility while yet maintaining high levels of milk yield.
It is clearly imperative for the breeding and dairy industry to put more emphasis on fertility, health and longevity, as well as for the pertinent research organisations and SME´s to engage in trait-orientated research, in order to be able to measure traits properly and at low cost, and to ensure that they are incorporated into the selection schemes, alongside with a possibly increasing use of genomic information. However, unless we gather more information on accurate phenotypes, there is an inherent risk that the molecular approach, such as genomic selection, lacks proper counter-information on animal physiology, behaviour and pathological constraints, thus becoming less beneficial than expected.
Acknowledgements
The authors´ own studies have been supported by grants from FORMAS and the Swedish Farmers´ Foundation for Agricultural Research(SLF), Stockholm, Sweden.
References
Sørensen AC, Lawlor T, Ruiz F. A survey on fertility in the Holstein populations of the world. In: Proceedings of the Int Conf on Fertility in dairy cows, Liverpool Hope University, UK, 30-31 August 2007 (EAAP Satellite Meeting), 2007; 1: 17
Humblot P. Use of pregnancy specific proteins and progesterone assays to monitor pregnancy and determine the timing, frequencies and sources of embryonic mortality in ruminants. Theriogenology 2001; 56: 1417-1433.
Andersen-Ranberg IM, Klemetsdal G, Heringstad B, et al. Heritabilities, genetic correlations and genetic change for female fertility and protein yield in Norwegian dairy cattle. J Dairy Sci 2005; 88: 348-355
Royal MD, Darwash AO, Flint APF, et al. Declining fertility in dairy cattle: changes in traditional and endocrine parameters of fertility. Anim Sci 2000; 70: 487-501
Sartori R, Gümen JN, Guenther JN, et al. Comparison of artificial insemination versus embryo transfer in lactating dairy cows. Theriogenology 2006; 65: 1311-1321
Bousquet, D, Bouchard E, DuTremblay D. Decreasing fertility in dairy cows: Myth or reality? In: Proceedings of the 23rd World Buiatrics Congress, Quebec, Canada, July 11-16, 2004, 6 pp
Lucy MC. Non-lactational traits of importance in dairy cows and applications for emerging biotechnologies. New Zealand Vet J 2005; 53: 406-415
Kuhn M, Hutchinson J. Factors affecting heifer fertility in US Holsteins. J Dairy Sci 2005; S88: A11
Pryce JE, Royal MD, Garnsworthy PC, et al. Fertility in the high-producing dairy cow. Livest Prod Sci 2004; 86: 125-135
Beever DE. The impact of controlled nutrition during the dry period on dairy cow health, fertility and performance. Anim Reprod Sci 2006; 96: 212-226
Veerkamp RF, Beerda B. Genetics and genomics to improve fertility in high producing dairy cows. Theriogenology 2007; 68S: S266-S273
Löf E, Gustafsson H, Emanuelson U. Associations between herd characteristics and reproductive efficiency in dairy herds. J Dairy Sci 2007; 90: 4897-4907
Roxström A, Strandberg E, Berglund B, et al. Genetic and environmental correlations among female fertility traits and milk production in different parities of Swedish Red and White dairy cattle. Acta Agric Scand, Sect A, Anim Sci 2001; 51: 7-14
Roxström A, Strandberg E, Berglund B, et al. Genetic and environmental correlations among female fertility traits and the ability to show oestrus, and milk production. Acta Agric Scand, Sect A, Anim Sci 2001; 51: 192-199
Hultgren J, Manske T, Bergsten C. Associations of sole ulcer at claw trimming with reproductive performance, udder health, milk yield, and culling in Swedish dairy cattle. Prev Vet Med 2004; 62: 233-251
Oltenacu PA, Algers B. Selection for increased production and the welfare of dairy cows: Are new breeding goals needed? Ambio 2005; 34: 311-315
Lindhé B. (Where is our breeding work going?] Vart är vi på väg i avelsarbetet? (In Swedish), Avelskuriren 2007; 2: 3-4
Chagas LM, Bass JJ, Blache D, et al. Invited review: New perspectives on the roles of nutrition and metabolic priorities in the subfertility of high-producing dairy cows. J Dairy Sci 2007; 90:4022-4032
Lucy MC. Reproductive loss in high-producing dairy cattle: Where will it end? J Dairy Sci 2001; 84: 1277-1293
Funk DA. Major advances in globalization and consolidation of the artificial insemination industry. J Dairy Sci 2006; 89: 1362-1368
Berglund B. Genetic improvement of dairy cow reproductive performance. Reprod domest Anim 2008; 43 S2: 89-95
Christie MF, Allcock J, Royal MD. Reproductive challenges facings today´s dairy industry. Proc. Int Conf on Fertility in dairy cows, Liverpool Hope University, UK, 30-31 August 2007 (EAAP Satellite Meeting), 2007; 1: 1
Maas JA, Garnsworthy PC, Flint AP. Modelling responses to nutritional, endocrine and genetic strategies to increase fertility in the UK dairy herd. Vet J 2008
Miller D. Spermatozoal RNA as reservoir, marker and carrier of epigenetic information: implications for cloning. Reprod Domest Anim 2007; 42 (Suppl 2): S2-S9
Miglior F, Muir BL, Van Doormaal BJ. Selection indices in Holstein cattle of various countries. J Dairy Sci 2005; 88: 1255-1263
Hansen LB. Consequences of selection for milk yield from a geneticist´s viewpoint. J Dairy Sci 2000; 83: 1145-1150
Gutierrez CG, Gong JG, Bramley TA, et al. Selection on predicted breeding value for milk production delays ovulation independently of changes in follicular development, milk production and body weight. Anim Reprod Sci 2006; 95: 193-205
Carlén E, Strandberg E, Roth A. Genetic parameters for clinical mastitis, somatic cell count, and milk production traits in the first three lactations of Swedish Holstein cows. J Dairy Sci 2004; 87: 3062-3080
Svensson C, Nyman AK, Persson Waller K, et al. Effects of housing, management, and health of dairy heifers on first lactation udder health in southwest Sweden. J Dairy Sci 2006; 89: 1990-1999
Emanuelson U, Danell B, Philipsson J. Genetic parameters for clinical mastitis, somatic cell counts, and miIk production estimated by multiple-trait restricted maximum likelihood. J Dairy Sci 1988; 71: 467-476
Philipsson J, Lindhe B. Experiences of including reproduction and health traits in Scandinavian dairy cattle breeding programmes. Livest Prod Sci 2003; 83: 99-112
Veerkamp RF, Beerda B, van der Lende T. Effects of genetic selection for milk yield on energy balance, levels of hormones, and metabolites in lactating cattle, and possible links to reduced fertility. Livest Prod Sci 2003; 83: 257-275
Weigel KA. Prospects for improving reproductive performance through genetic selection. Anim Reprod Sci 2006; 96: 323-330
Windig JJ, Calus MPL, Veerkamp RF. Influence of herd environment on health and fertility and their relationship with milk production. J Dairy Sci 2005; 88: 335-347
Windig JJ, Calus MPL, Beerda B, Veerkamp RF. Genetic correlations between milk production and health and fertility depending on herd environment. J Dairy Sci 2006; 89: 1765-1775
Wade GN, Jones JE. Neuroendocrinology of nutritional infertility. Am J Physiol Regul Integr Comp Physiol. 2004; 287: 1277-1296
Agnew RE, Yan T, Murphy JJ, et al. Development of maintenance requirement and energetic efficiency for lactation from production data of dairy cows. Livest Prod Sci 2003; 82: 151-162
Knight CH, Beever DE, Sorensen A. Metabolic loads to be expected from different genotypes under different systems. Metabolic stress in dairy cows. British Soc Anim Sci 1999; Occas Publ 24: 27-36
Veerkamp RF. Selection for economic efficiency of dairy cows using information on liver weight and feed intake: A review. J Dairy Sci 1998; 81: 1109-1119
Grummer RR. Strategies to improve fertility of high yielding dairy farms: Management of the dry period. Theriogenology 2007; S68: S281-S288
Ingvartsen KL, Dewhurst RJ, Friggens NC. On the relationship between lactational performance and health: is it yield or metabolic imbalance that causes diseases in dairy cattle? A position paper. Livest Prod Sci 2003; 83: 277-308
Hjertén J. [Relationships between body condition score, subcutaneous fat, live weight and reproduction in Swedish Holstein and Swedish Red and White Cattle]. In Swedish. Summary in English. MSc thesis 281, SLU, Dept of Animal Breeding and Genetics, Uppsala, Sweden 2006; 35 pp
Collard BL, Boettcher PJ, Dekkers JCM, et al. Relationships between energy balance and health traits of dairy cattle in early lactation. J Dairy Sci 2000; 83: 2683- 2690
Jensen MB, Munksgaard L, Pedersen LJ, Ladewig J, Matthews L. Operant conditioning as a method to assess lying motivation in dairy heifers. Proc 36th Int ISAE Congr, The Netherlands August 6-10 2002, p. 69
Lidfors L, Isberg L. Intersucking in dairy cattle- review and questionnaire. Appl Anim Behav Sci 2002; 42: 87-94
Dobson H, Smith RF, Royal MD, et al. The high-producin dairy cow and its reproductive performance. Reprod Domest Anim 2007; 42 (Suppl 2): 17-23
Rajala-Schultz PJ, Gröhn YT. Culling of dairy cows. Part I. Effects of diseases on culling in Finnish Ayrshire cows. Prev Vet Med 1999a; 41: 195–208
Rajala-Schultz PJ, Gröhn YT. Culling of dairy cows. Part II. Effects of diseases and reproductive performance on culling in Finnish Ayrshire cows. Prev Vet Med 1999b; 41, 279–294
Rajala-Schultz PJ, Gröhn YT. Culling of dairy cows. Part III. Effects of diseases, pregnancy status and milk yield on culling in Finnish Ayrshire cows. Prev Vet Med 1999c; 41, 295–309
Onyiro OM, Brotherstone S. Genetic analysis of locomotion and associated conformation traits of Holstein-Friesian dairy cows managed in different housing systems. J Dairy Sci 2008; 1:322-328
Manske T. Hoof lesions and lameness in Swedish dairy cattle; prevalence, risk factors, effects of claw trimming and consequences for productivity. PhD-Thesis, SLU, Skara, 2002
Søgstad AM, Østeras O, Fjeldaas T. Bovine claw and limb disorders related to reproductive performance and production diseases. J Dairy Sci 2006; 89: 2519-2528
Telezhenko E, Bergsten C. Influence of floor type of the locomotion of dairy cows. Appl Anim Behav Sci 2005; 93: 183-197
Cook N, Nordlund K. Behavioural needs of the transition cow and considerations for special needs facility design. Vet Clin North Am, Food Anim Pract 2004; 20: 495-520
Von Keyserlingk MA, Olenick D, Weary DM. Acute behavioural effects of regrouping dairy cows. J Dairy Sci 2008; 91: 1011-1116
Beilharz RG. Quantitative genetics and evolution: is our understanding of genetics sufficient to explain evolution? J Anim Breed Gen 1993 110: 161-170
Lindqvist C. Domestication effects on foraging behaviour – consequences for adaptability in chickens. Linköping Studies in Science and Technology. 2008, Dissertation No. 1164
De Vries MJ, Veerkamp RF. Energy balance of dairy cattle in relation to milk production variables and fertility. J Dairy Sci 2000; 83: 62-69
Butler WR. Energy balance relationships with follicular development, ovulation and fertility in postpartum dairy cows. Livest Prod Sci 2003; 83: 211-218
Llewellyn S, Fitzpatrick R, Kenny DA, et al. Effect of negative energy balance on the insulin-like growth factor system in pre-recruitment ovarian follicles of post partum dairy cows. Reproduction 2007; 133: 627-639
Leroy JL, Vanholder T, Mateusen B, et al. Non-sterified fatty acids in follicular fluid of dairy cows and their effect on developmental capacity of bovine oocytes in vitro. Reproduction 2005; 130: 485-495
Vanholder T, Leroy JL, van Soom A, et al. Effect of non-esterified fatty acids on bovine granulosa cell steroidogenesis and proliferation in vitro. Anim Reprod Sci 2005; 87: 33-44
Patton J, Kenny DA, McNamara S, et al. Relationships among milk production, energy balance, plasma analyses, and reproduction in Holstein-Friesian cows. J Dairy Sci 2007; 90: 649-658
Alban L, Agger JF. Welfare in Danish dairy herds 1. Disease management routines in 1983 and 1994. Acta vet Scand 1996; 37: 49-63
Alban L, Agger JF. Welfare in Danish dairy herds 2. Housing systems and grazing procedures in 1983 and 1994. Acta vet Scand 1996; 37: 65-77
Platz S, Ahrens F, Bendel J, et al. What happens with cow behaviour when replacing concrete slatted floor by rubber coating: A case study. J Dairy Sci 2008; 91: 999-1004
Bergsten C. Effects of conformation and management system on hoof and leg diseases and lameness in dairy cows. Vet Clin North Am Food Anim Pract 2001; 17: 1-23
Roche JF. The effect of nutritional management of the dairy cow on reproductive efficiency. Anim Reprod Sci 2006; 96: 282-296
Rukkwamsuk T, Kruip TA, Wensing T. Relationship between overfeeding and overconditioning in the dry period and the problems of high producing dairy cows during the postparturient period. Vet Q 1999; 21: 71-77
Smith BI, Risco CA. Predisposing factors and potential causes of postpartum metritis in dairy cattle. Comp Contin Educ Pract Vet 2002; 24: S74-S80
Stronge AJH, Sreenan JM, Diskin MG, et al. Post insemination milk progesterone concentration and embryo survival in dairy cows. Theriogenology 2005; 64: 1212-1224
Al-Katanani YM, Webb DW, Hansen PJ, Factors affecting seasonal variation in 90-day nonreturn rate to first service in lactating Holstein cows in a hot climate. J Dairy Sci 1999; 82: 2611-2616
Snijders SE, Dillon PG, O'Farrell KJ, et al. Genetic merit for milk production and reproductive success in dairy cows. Anim Reprod Sci 2001; 65:17-31
Sartori R, Sartori-Bergfelt R, Mertens SA, et al. Fertilization and early embryonic development in heifers and lactating cows in summer and lactating and dry cows in winter. J Dairy Sci 2002; 85: 2803-2812
Rodriguez-Martinez H, Barth AD. In vitro evaluation of sperm quality related to in vivo function and fertility. In: Reproduction in Domestic Ruminants VI. Edited by JI Juengel, JF Murray, MF Smith. Nottingham University Press, Nottingham, UK, 2007; 39-54 (Soc Reprod Fert 2007; 64: 39-54)
Ostermeier GC, Goodrich RJ, Moldenhauer JS, et al. A suite of novel human spermatozoal RNAs. J Androl 2005; 26: 70-74
Miller D, Ostermeier GC. Spermatozoal RNA: Why is it there and what does it do? Gynecol Obstet Fertil 2006; 34: 840-846
Lalancette C, Thibault C, Bachand I, et al. Transcriptome analysis of bull semen with extreme nonreturn rate: Use of suppression-substractive hybridization to identify functional markers for fertility. Biol Reprod 2008; 78: 618-635
Roche JF, Mackey D, Diskin MD. Reproductive management of postpartum cows. Anim Reprod Sci 2000; 60-61: 703-712
Lucy MC. Mechanisms linking nutrition and reproduction in postpartum cows. Reproduction 2003; S61: S415-S417
Reksen O, Grohn YT, Havrevoll O, et al. Relationships among milk progesterone, concentrate allocation, energy balance, milk yield and conception rate in Norwegian cattle. Anim Reprod Sci 2002; 73: 169-184
Inskeep EK. Preovulatory, postovulatory, and postmaternal recognition effects of concentrations of progesterone on embryonic survival in the cow. J Anim Sci 2004; 82: E24-E39
Lopez H, Satter LD, Wiltbank MC. Relationship between level of milk production and estrous behavior of lactating dairy cows. Anim Reprod Sci 2004; 81: 209-223
Vanholder T, Opsomer G, de Kruif A. Aetiology and pathogenesis of cystic ovarian follicles in dairy cattle: a review. Reprod Nutr Dev 2006; 46: 105-119
Wiltbank M, Lopez H, Sartori R, et al. Changes in reproductive physiology of lactating dairy cows due to elevated steroid metabolism. Theriogenology 2006; 65: 17-29