Selenium and selenoproteins in poultry production
Selenium sources and selenoproteins in practical poultry production
Nutritional needs of food animals must be met by provision of nutrients from plants, soil, and even from prey animals. Many of the potential nutrients are minerals that exist in chemical complexes that are not readily available.
This condition is advantageous for animals because many of the minerals can be toxic, but nutritionists have demonstrated that a large number of minerals, either as macronutrients or as micronutrients, are required for normal growth and development of animals including humans. The macronutrients consist of minerals such as calcium, phosphorus, sodium, and potassium because these are involved in structural integrity of the body and in homeostatic mechanisms.
Micronutrients, often called trace nutrients, include minerals such as magnesium, manganese, zinc, iron, copper, and there are ultramicronutrient minerals such as molybdenum, selenium, iodine, cobalt, chromium and even vanadium. The trace minerals as either micro- or ultramicronutrients function as parts of proteins, hormones, enzymes, or as co-factors that activate specific enzymes.
Nature’s provision of nutrients to the inhabitants of the earth is a process that can be very variable.
Although common foodstuffs such as corn, wheat, barley, soybeans, or oats are now grown around the world, they are not the same world wide. In the case of foodstuffs that provide selenium, some can be enriched as the result of high concentrations of selenium in the soil. In contrast, some foodstuffs can have very low levels of selenium because the soil in which they are grown had very low concentrations.
This situation requires that supplemental selenium be provided via the manufactured feed to ensure optimal performance of food production animals.
Among the ultramicronutrients, selenium may hold the distinction of being the most difficult to understand. It can exist in four valence states: -2 (hydrogen selenide, sodium selenide, dimethyl selenium, trimethyl selenium, and selenoamino acids such as selenomethionine), 0 (elemental selenium), +4 (selenium dioxide, selenous acid, and sodium selenite), and +6 (selenic acid and sodium selenate), and depending upon its valence state and water solubility, the gastrointestinal absorption rate can be affected. Selenium can be toxic in valence states –2, +4 and +6, but in appropriate trace levels in feed or drinking water, selenium in valence states –2, +4 and +6 can also serve in several essential roles in maintenance of the body’s homeostatic condition.
Forms and availability of selenium used in animal production
Selenium supplementation of poultry rations is now a routine procedure. Since 1974, when the US Food and Drug Administration (FDA) approved selenium as a feed supplement, sodium selenite has become the traditional source for dietary supplemental selenium for poultry and livestock (Leeson and Summers, 1991). Other inorganic sources of selenium are sodium selenate and calcium selenate (Echevarria et al., 1988a,b).
Poultry obtain all of their selenium through their feed. The commonly used feed ingredients, such as cereal products (Burk, 1976) and meals of fish, poultry and meat (Levander 1986; Cai et al., 1995), contain selenium almost exclusively as organic compounds such as the naturally occurring selenoamino acids (selenomethionine and selenocysteine). The selenoamino acids are incorporated into protein, principally as selenomethionine and selenocysteine, and constitute 50 to 80% of the total selenium in plants and grains (Butler and Peterson, 1967).
Estimates of bioavailability of selenium for poultry vary considerably depending upon which criteria are used for the evaluation. Scott and Thompson (1971) determined that there was a linear relationship in tissue deposition of dietary inorganic selenium up to 0.3 mg/kg. Cantor et al. (1975a,b) have reported diverse and variable results when exudative diathesis or pancreatic fibrosis were used to evaluate selenium bioavailability.
In both studies, plant-based selenium sources were superior to inorganic sources, but there were low protective values associated with plantbased organic selenium in terms of protection against development of exudative diathesis (Cantor et al., 1975a). In contrast, Osman and Latshaw (1976) reported that selenomethionine was at least equivalent to sodium selenite in the protection of the chick against exudative diathesis. Cantor et al., (1975b) reported high availability of selenomethionine in protection of the chick against pancreatic fibrosis.
Echevarria et al. (1988a,b) indicated that sodium selenite, sodium selenate, and calcium selenate are absorbed equally well by chicks based upon tissue distributions of selenium after short term feeding trials with high levels of the inorganic selenium sources. Echevarria et al. (1988a,b) demonstrated that selenium concentrations ranged from highest to lowest in kidney, liver, muscle, and plasma, respectively.
Increasing levels of dietary inorganic selenium were associated with greater tissue concentrations. The increasing tissue concentrations of selenium were time dependent with liver showing weekly increases in selenium concentrations whereas selenium reached a plateau within seven days of feeding in kidney, muscle, and plasma. In a study conducted with rats, Vinson and Bose (1987) found that organic selenium from yeast was more available to blood and liver proteins than from either sodium selenite or an amino acid-selenium chelate (Table 1).
Table 1.Bioavailability of selenium forms in the inorganic sodium selenite, amino acid chelate, and organic selenomethionine in yeast (Vinson and Bose, 1987).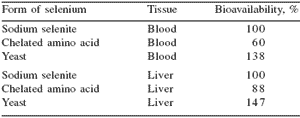
The availability of selenium from different sources such as selenomethionine or inorganic selenium from selenite or selenate may be affected by numerous factors. Cantor and Johnson (1985) concluded that selenium was made more available in diets that were low in protein, possibly as a result of decreased total methionine in the diet.
The kind and quality of fish meal that has been used in poultry diets also influences the availability of selenium for poultry (Miller et al., 1972; Whitacre and Latshaw, 1982). Furthermore, intestinal absorption of selenium in foodstuffs varies by species, and bioavailability varies according to the form that is eaten (Gabrielsen and Opstevedt, 1980; Douglas et al., 1981; Ringdal et al., 1985; Wen et al., 1997; Schen et al., 1997). Schen et al. (1997) reported that selenomethionine and sodium selenate are more diffusible than selenocysteine and sodium selenite, explaining the higher absorption rate for selenomethionine and sodium selenate.
However, utilization of selenium from sodium selenate can be quite variable because a significant fraction may be lost in urine before it can be incorporated into body tissues (Xia et al., 1992).
Bioavailability of selenium has been estimated by a tissue residue approach by numerous investigators.
Miller et al. (1972) found selenomethionine to be more available to chicks than selenium from sodium selenite or selenium from fish products. Cantor et al. (1982) observed significantly more selenium from selenomethionine in muscles of turkey poults than selenium from inorganic selenium sources.
However, Cantor and Tarino (1982) showed that availability of selenium from selenite was greater than from selenomethionine when plasma glutathione peroxidase (GSH-Px) activity was used an indicator. Moksnes and Norheim (1986) also demonstrated greater tissue concentrations of selenium and GSH-Px activity in selenomethionine-fed chickens compared with sodium selenite-fed chickens. There is no question that selenomethionine will increase tissue concentrations of selenium as compared with selenium deposition from inorganic selenium.
When the FDA approved selenium supplements for poultry and swine in 1974, the animal industries settled on the use of inorganic selenium forms rather than the organic selenoamino acids. The primary reason for that decision was based on cost of the selenium supplements and lack of information on selenomethionine.
The inorganic forms were cheap and the organic forms were expensive. Without a doubt, the use of inorganic selenium supplements in feeds has improved the performance of all classes of commercial poultry. In modern high-yielding poultry, which have higher metabolic rates and different nutritional needs compared with poultry from 30 years ago, there is a need to reassess nutrient requirements.
Inorganic selenium has some problems associated with its use. Among those problems are the minimal levels of selenium in meat proteins and the potential for toxicity if too high a dietary level of inorganic selenium is provided to chickens. Thus, a need to revisit organic selenium as a feed supplement for poultry is apparent.
After many years of laboratory and field research, a source for natural organic selenium (Sel-Plex®, Alltech Inc., Nicholasville, KY), was approved for use in the poultry industry in the US by the FDA (Federal Register, 2000; 2002).
Sel-Plex® provides a broad spectrum of selenium compounds (Kelly and Power, 1995), but selenomethionine in the seleniumenriched yeast cellular protein component is the primary form of selenium in Sel-Plex®. The organic selenium profile in Sel-Plex® is similar to the organic selenium profile in plants and grains (Kelly and Power, 1995). The organic selenium in Sel-Plex® is readily available and will be absorbed actively (Mahan, 1994; 1995) from the intestine via the Na+- dependent methionine transport system (Spencer and Blau, 1962) while sodium selenite is absorbed passively by diffusion from the intestinal tract (McConnell and Cho, 1965; Schrauzer, 2001).
Even though organic selenium in Sel-Plex® is a superior source of selenium for poultry production (Edens, 1996; 2002; Edens and Sefton, 2002; Edens et al., 2002), sodium selenite is still used as the principal source of selenium in animal feeds. This seems paradoxical because sodium selenite has a documented pro-oxidant influence in all animals tested including humans (Hafeman et al., 1974; Csallany and Menken, 1986; Spallholz, 1997; Terada et al., 1999).
Selection of the best form of organic selenium as a feed supplement
Based on natural availability of organic selenium in plant–based foodstuffs and its superior bioavailability, it should be easy to decide to provide organic feed supplements, rather than the pro-oxidative sodium selenite, to food producing animals.
However, the choices among selenium supplements available to feed compounders and commercial nutritionists are rather large and diverse in their composition. Schrauzer (2001) has reported that there are quality concerns associated with many selenium supplements.
Some yeast products do not contain organic selenium.
Instead they are made with inorganic sodium selenite or sodium selenate. Some feed supplements do not tell which form of selenium is provided. Some products are made with selenium proteinates or selenoamino acid chelates. Schrauzer also pointed out that vitamin supplements for humans are often formulated with vitamin C and that over time, the selenium is reduced to elemental selenium and is not available.
Thus, it is clear that the compounders and commercial nutritionists have had a difficult task in making an informed decision concerning the appropriate product to use for supplementation of organic selenium. There are many companies around the world that produce organic selenium yeast products. Each manufacturer has claimed superior performance of food animals fed their product.
However, little is known about speciation of selenium in selenized yeast products. In a recently published article, investigators, using state-of-the-art methodologies, studied the fractionation of selenium in yeast into different classes of chemical species and determined the true speciation of protein-incorporated selenium in individual yeast proteins characterized by unique amino acid sequences (Encinar et al., 2003).
In this study, they used three commercial yeast products. They quantified water soluble selenium, water insoluble polysaccharide-bound selenium, water-insoluble protein-bound selenium, residual protein-bound selenium, and residual hydrolysable selenium (Table 2). Based on these percentage values, it is apparent that the profile of selenocompounds differs markedly, which indicates that all sources of ‘selenium yeast’ are not alike.
Thus, as one begins to think about and then select a yeast product for dietary organic selenium supplementation for food producing animals, it is important to know about the availability of the selenium in the product.
Table 2. Selenium species in three selenized yeast products1.
1Percentage values based on Encinar et al., 2003
Why is selenium an essential nutrient?
The essentiality of selenium as an ultramicronutrient in the daily nutrition of mammals was demonstrated by Schwarz and Foltz (1957). Their discovery that selenium deficient rats would suffer hepatic necrosis was rapidly followed by the discovery that selenium was essential in poultry (Schwarz et al., 1957).
At Lederle Laboratories, Pearl River, New York, Patterson et al. (1957) made an independent observation that exudative diathesis in chickens was prevented when selenium was added to the feed. It was not until 1973 that the beneficial effects of selenium could be ascribed a biochemical function when Rotruck et al. (1973) and Flohé et al. (1973) discovered that it was an essential component of an important antioxidant enzyme, glutathione peroxidase (GSH-Px). Continued study of GSH-Px has shown that there are at least six isoenzyme forms in various organs and tissues in mammalian species (Table 3).
Corresponding evidence is not available in avian species, but empirical data from studies with selenium in poultry suggest that several functional GSH-Px isoenzymes and other selenoproteins are also present in avian species. This would suggest that at least for GSH-Px and some of the other selenoproteins, the active selenocysteine site on the enzyme/protein has been conserved.
Conservation of active sites may not be limited to vertebrate animals. Indeed, in a parasite, the blood fluke Schistasoma mansoni, phospholipidhydroperoxide GSH-Px has been discovered (Maiorino et al., 1996). Thus, selenium has a special function in antioxidant control mechanisms in animal species.
Mertz (1987) suggested that selenium is unique among the essential trace elements especially in the manner in which its deficiency is expressed. Mertz (1987) stated that a selenium deficiency in mammals was likely to be manifest in second generation progeny.
Selenium deficient rats with clinical signs die within a few weeks if they become vitamin E deficient (Schwarz, 1951), but if rats in the first generation of selenium deficiency are force-fed high levels of fat, they die within a few hours (Schwarz, 1954). These observations by Schwarz and by Mertz pointed to the complex interaction between selenium and vitamin E and established that there was a need for adequate levels of both in diets of humans and food animals.
A survey of the literature readily reveals that selenium, through the selenoproteins, influences numerous physiological and biochemical functions that are necessary for maintenance of homeostasis in all animals.
Edens (1996) listed a number of problems associated with selenium deficiency. Selenium deficiency can manifest itself in many diseases and dysfunctions such as liver necrosis, muscular dystrophy, microangiopathy, exudative diathesis, pancreatic fibrosis, poor feathering, retained placenta, mastitis, cystic ovaries, general unthriftiness, Keshan disease, Kashin-Beck disease, cancer, numerous heart diseases, immune deficiencies, reduced fecundity, and many others and can affect humans and food production animals alike.
These listed problems show us that no animal is exempt from selenium deficiency syndromes. Therefore, one can confidently state that selenium is essential and must be provided if natural intake is below optimal requirements.
Selenium in body proteins
Schrauzer (2003) reported that selenomethionine is incorporated into body proteins in place of methionine, providing a means for reversible storage of selenium in organs and tissues. This property of selenomethionine is not shared with any other selenoamino acid, which points to a specific physiological function of selenomethionine (Schrauzer, 2003).
In fact, Schrauzer (2003) has asserted that all needed metabolic forms of selenium can be produced from selenomethionine, and it, therefore, meets all the criteria for an essential amino acid. The situation associated with the use of inorganic selenium is different because inorganic selenium from selenite or selenate results in very limited nonspecific insertion of selenocysteine into protein but serves as a substrate for synthesis of selenocysteine for specific insertion into selenoproteins (Hawkes et al., 1985a,b; 2003).
Selenium from sodium selenite or selenate might also form a selenotrisulfide (-S-Se-S-) bond (Ilian and Whanger, 1989) that is subject to rapid oxidation and release from protein. The selenotrisulfide bond is not formed with selenomethionine.
Cummins and Martin (1967) and Latshaw and Osman (1976) have demonstrated that selenium derived from selenite was easily released from animal protein subjected to alkaline dialysis whereas selenium from selenomethionine was retained as a part of the protein.
Ort and Latshaw (1978) have shown that even when toxic levels of selenomethionine or sodium selenite are fed to laying hens, egg levels and body tissue levels return to normal within two to four weeks after cessation of feeding toxic levels of selenite selenium, but a longer time is required for birds fed selenomethionine.
Selenium from selenite, therefore, appears to interact loosely with the cysteine thiol group and selenium in selenomethionine is molecularly integrated into protein. The stored form of organic selenium as selenomethionine is in a non-functional state, i.e. not used immediately for formation of biologically functional selenoproteins (Mahan, 1994; 1995).
Since selenium in selenomethionine is better retained than inorganic selenium, the ultimate metabolism of organic selenium with liberation of selenide that enters the pathway for selenocysteine synthesis for incorporation into functional selenoproteins appears to be more efficient.
As a consequence of the large pool of stored selenium in protein, in times of oxidative stress, body protein can be degraded rapidly providing more than adequate concentrations of organic selenium that can be used for synthesis of specific selenoproteins.
Constant provision of a readily available source of organic selenium from body proteins for synthesis of selenoproteins is a unique and continuous process.
Mitch and Goldberg (1996) have shown that most proteins are degraded within a few hours after their synthesis. Functional proteins such as enzymes have a more rapid degradation rate than do structural proteins such as muscle protein. Consumption of diets containing organic selenium results in a high rate of selenomethionine incorporation in muscle protein.
As the muscle protein is replaced, the stored selenomethionine is released into the free amino acid pool from which it is made available for selenoprotein synthesis. In times of oxidative stress, the proteasome, a cytosolic organelle, increases the rate of protein degradation and increases amino acid availability for synthesis of other proteins such as the selenoproteins.
In fast growing animals such as the high-yielding commercial broiler lines, it is important to have a readily available source of selenium to be used in selenoprotein synthesis.
Selenium incorporation into selenoproteins
The interest in the antioxidant properties of selenium has been facilitated by the fact that the active site(s) on the various selenoproteins, which often act in antioxidant systems, contain selenocysteine- the proclaimed 21st amino acid. The locus of selenocysteine in selenoproteins involves use of a stem-loop structure in the 3´ non-translated region of the mRNA that designates selenocysteine insertion at the UGA codon instead of chain termination (Berry et al., 1991a,b; 1993).
In order for selenocysteine to be incorporated into specific selenoproteins, there is a requirement for selenocysteine-ß-lyase reaction with free selenocysteine that causes the release of selenide in the presence of reducing agents (Sunde, 1990; Burk, 1991). Another selenoenzyme, selenophosphate synthetase, using selenide and serine as substrates, phosphorylates selenide to form selenophosphate. The selenophosphate is made available to a unique seryltRNASEC that is recognized by selenocysteine synthetase.
Under the influence of selenocysteine synthetase, that converts seryl-tRNASEC to selenocysteyl-tRNASEC, the selenium in selenophosphate is co-translationally incorporated into selenocysteine. Then, selenocysteyltRNA[ Ser]SEC, which recognizes the specific UGA codon in the selenoprotein-mRNA, inserts the new, cotranslationally synthesized selenocysteine into the specific selenoprotein (Burk, 1991).
The base triplet UGA that normally functions as a stop codon (Amberg et al., 1996) encodes this process of selenocysteine insertion at its appropriate site in the peptide. The selenocysteine insertion also requires a specific mRNA, an elongation factor, GTP, and the selenocysteine insertion sequence (SECIS) that all interact at the ribosome to read the UGA selenocysteine codon (Low and Berry, 1996).
Thus, organic selenium must be converted from its original organic form (-2 valence) to the inorganic selenide (also -2 valence) form then back to an organic form (-2 valence) to fulfill its biological function (Arthur, 1997). This conversion is crucial with regard to synthesis of selenoproteins because it has been reported that 30 to 80% of the selenium in the body may be selenocysteine (Hawks et al., 1985a).
Nevertheless, selenomethionine is a highly available substrate for many proteins and can substitute nonspecifically for methionine in their structure (Daniels, 1996). In cases of selenomethionine supplementation to feeds, it can be demonstrated that 40 to 50% of total body selenium, as selenomethionine, can be found in muscle protein (Daniels, 1996). Natural selenocysteine also can be substituted non-specifically for cysteine in many proteins, but it is not incorporated directly into specific selenoproteins (Sunde, 1990; Daniels, 1996).
Animals cannot synthesize selenomethionine, the primary selenoamino acid, directly from selenite or selenate forms of inorganic selenium (Cummins and Martin, 1967; Sunde, 1990). Selenoamino acids are acquired naturally from plants and grains and animal protein eaten by the animals. Selenocysteine can be found in animals fed inorganic selenium such as selenite and selenate. In animals consuming selenium rich foodstuffs, selenomethionine is present in high concentrations and is easily converted to selenocysteine via cystethionase (Esaki et al., 1981).
Selenium functions through selenoproteins
In mammals there are at least 18 known (Hatfield and Gladyshev, 2003) and possibly more genes that encode selenoproteins (Behne et al., 1997). Recent evidence strongly suggests that there are at least 25 selenoproteins in the mammalian selenoproteome (Kryukov et al., 2003), and this would suggest more genes that encode selenoproteins.
The known selenoproteins have numerous functions, but many of the selenoproteins still have unknown functions (Table 3). Furthermore, it is not known if all animals possess equivalent proteins. An excellent review of the functions of selenoproteins in farm animals was presented by Jacques (2001) who pointed out that selenium must exert its greatest influences through selenoproteins.
In poultry there has been limited research to ascertain the presence and function(s) of the various selenoproteins. It appears from the few publications on avian selenoproteins that their functions are generally similar to the corresponding protein(s) in mammals (Petovich and Podorozhnaia, 1981; Ilian and Whanger, 1989; Snityns’kyi and Antoniak, 1994).
Of the selenoproteins studied in chickens, it has been reported that there are two deiodinases: Type I that functions in the liver to convert thyroxin (T4) to triiodothyronine (T3), and Type III that functions to convert T3 to rT3 and T3 to T2 (May, 1989; Van der Geyten et al., 1997). There are numerous reports on GSH-Px activity(ies) in chickens, and those reports suggest that the GSH-Px enzymes function similarly to those found in mammals because of their specificity for reduction of hydrogen peroxide and lipid peroxides (Stadtman, 1980; Pablos et al., 1995a,b; 1998).
Recent evidence has shown that chickens express phospholipid-hydroperoxide GSH-Px (Kong et al., 2003), supporting observations of its presence in several chicken and turkey tissues (Surai et al., 1998a, b). The chicken is different from most mammals because it also expresses a monomeric GSH-Px in its tissues (Miyazaki and Motoi, 1992; 1996).
Finally, thioredoxin reductase (TrxR) activity has been reported for the chicken (Smith and Levander, 2001; 2002), and it was determined that the activity of chicken TrxR is very much less than mammalian TrxR. Tissue distribution of chicken TrxR has not been determined yet, but preliminary data will be presented herein. The presence of other selenoproteins can be inferred from current and previous studies; but based on the literature, it is possible that the chicken will express selenoproteins similar to mammals.
Edens (2001) reported that broiler chickens given a Sel-Plex® supplement in the feed were more resistant to enteropathogenic E. coli enteric infection than chickens that had no supplemental selenium. The broilers given Sel-Plex® had body weights and feed conversions that were similar to the birds with no supplemental selenium and not challenged with the bacteria (Table 5). Mortality of E. coli-challenged Sel-Plex®-fed broilers was 59% less than mortality in E. coli-challenged broilers given no supplemental selenium (Table 5).
Additionally, the GSH/GSSG ratios in broilers given Sel-Plex® were less than those that were not given supplemental selenium (Table 6). This suggested that the GSH/GSH-Px system was working more efficiently in broilers fed Sel-Plex®.
This conclusion was supported by the observation that there was less heat shock protein (hsp) 70 in the intestines of Sel-Plex®-fed birds compared with those given no supplemental selenium (Figure 1).
Sulfur-containing amino acids (cysteine and methionine) are susceptible to oxidation. The sulfhydryl groups of cysteine residues of proteins are normally maintained in a reduced state or they can be oxidized by thioredoxins in the cytoplasm (Okamoto et al., 1999). During oxidative stress, thiol-disulfide exchange occurs between cysteine residues in protein and oxidized glutathione (GSSG).
This process serves as a redox-dependent regulator of various protein functions (Fratelli et al., 2002).
The oxidized sulfhydryl in methionine can be reduced by the methionine sulfoxide reductases (Grimaud et al., 2001) and might serve in an antioxidative capacity (Levine et al., 1996; Stadtman et al., 2002). These mechanisms are interrelated because the heat shock proteins elevate reduced glutathione levels by promoting an increase in glucose-6-phosphodehydrogense activity (Preville et al., 1999).
The heat shock proteins are normally induced during oxidative stress and protect sensitive sites on proteins. In the results of two studies presented here, we concluded that broilers given Sel-Plex® and exposed to either heat stress (Table 4) or enteric an E. coli challenge (Figure 1) performed better because they had lower concentrations of liver hsp70 than broilers given sodium selenite or no supplemental selenium (Edens, 2001).
If the hypothesis presented here is correct, the selenium in selenomethionine and selenocysteine in proteins might be less reactive with oxidized GSSG, and the GSSG would be reduced more readily by glutathione reductase as postulated by Mahmoud and Edens (2003). As a result, when organic selenium is fed to broilers, less heat shock protein would be induced because the cell cytoplasm would be maintained in a more reduced status even in the face of stressors that promote oxidative distress (Table 4).
Table 3.Selenoproteins and their role in the maintenance of homeostasis in animals.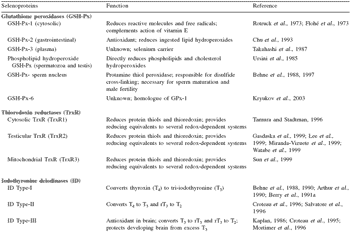

To enlarge the image, click here
Table 4.Influence of selenium source on blood and liver total glutathione (TGSH), reduced glutathione (GSH), oxidized GSH (GSSG), GSH/ GSSG, glutathione peroxidase (GSH-Px), glutathione reductase (GR), and hsp70 in chickens before and after mild heat distress.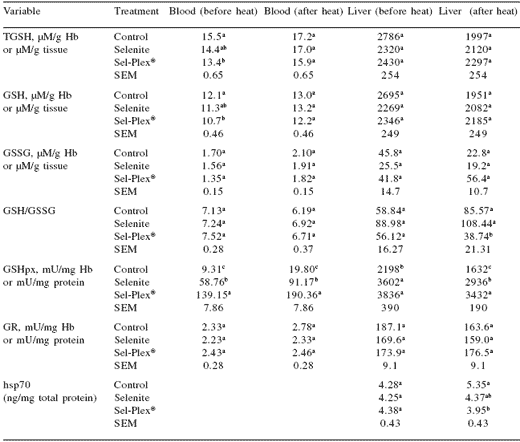
a,b,cIn a column, means with unlike superscripts within a variable differ significantly (P≤0.05).
Table 5. Performance of 42 days old male broiler chickens given a dietary supplement of Sel-Plex® and challenged with an enteropathogenic E. coli.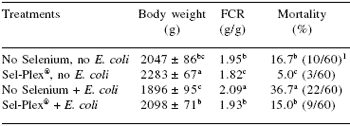
a,b,c In a column, means with unlike superscripts differ (P≤0.05). 1Numbers in parentheses represents total mortality per treatment.
The data from these two different experiments suggest that in three tissues there might be three or four different GSH-Px, specifically the classical GSH-Px-1, GSH-Px-2, GSH-Px-3 and monomeric GSH-Px. Although, the data cannot predict which isoform was active/not active, it was apparent that the GSH/GSH-Px system in the chicken is highly responsive to selenium both as sodium selenite and as organic selenomethionine in yeast protein.
Table 6. Influence of Sel-Plex® on intestinal total glutathione (μM/g), oxidized glutathione (μM/g), reduced glutathione (μM/g), and the reduced:oxidized glutathione ratios in broiler chickens given a dietary supplement of organic selenium and challenged with an enteropathogenic E. coli.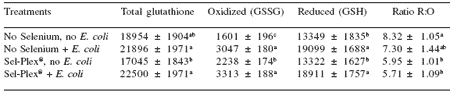
a,b,c In a column, means with unlike superscripts differ significantly (P≤0.05).
Figure 1.Intestinal heat shock protein 70 response to enteropathogenic E. coli (EC) infection in broiler chickens with and without supplemental Sel-Plex® .
Without a highly functional selenium-dependent antioxidant system, the modern broiler chicken would not be as productive. As an illustration, Edens (2001) presented performance data of broilers grown to 42 days of age. In that experiment Sel-Plex®-fed broilers had body weight improvement of 20 g over selenite supplemented broilers and a 50 g improvement over broilers with no selenium supplementation. Feed conversion in Sel-Plex®-fed broilers was improved 3 points over selenite-fed birds and 9 points over birds with no supplemental selenium.
Monodeiodinase and triiodothyronine: influences on diverse functions and poultry performance
The thyroid hormone, triiodothyronine (T3), has many important functions in all animals. In poultry, T3 has the potential to affect the efficiency of conversion of food into energy and metabolic heat production.
The mitochondria, the cell’s power generators, are affected directly by T3. Rapid protein synthesis, mitochondrial gene transcription, and synthesis of proteins from genetic information are affected by T3.
These processes cause turnover of body proteins, increase free fatty acids for energy metabolism, and increase oxygen usage by cells in the body.
Additionally, the cardiovascular system is stimulated to a higher level of activity to meet the tissue demands for increased oxygen caused by higher levels of T3.
Body temperature regulation is also partly under the control of circulating T3 that influences mitochondrial activity in muscles. The circulating T3 level is highly responsive to caloric intake and external temperature and is necessary for cold adaptation. If caloric intake increases or external temperature decreases, there is an increase in circulating T3.
Conversely, with starvation T3 decreases along with increased thyroxin (T4). The development and growth of animals is directly affected by T3 binding to thyroid receptors (TR) that bind to responsive elements on the nuclear DNA triggering gene transcription. This T3 function in all animals is extremely important in the developing embryo for growth and hatching efficiency and in the post-hatch chicken for growth.
Recently, a report was given that sodium selenite actually inhibited the development of the T3-TR bond, but organic selenium did not inhibit that relationship (Brtko et al., 1997).
Involvement of thyroid hormones in feathering has long been recognized (Radi and Warren, 1938; Boone et al., 1950). Feeding thyroprotein increases feathering rate in slow feathering birds. In thyroprotein there is a predominance of thyroxin (T4), but in poultry, triiodothyronine (T3) is the most active thyroid hormone. It is known that T3 is intimately involved in feather development.
A small quantity of T3 is produced in the thyroid, but the greatest quantity is converted from T4 to T3 in the liver by the seleniumdependent type I 5′-deiodinase enzyme (Berry et al., 1991a) that predominates in the chicken (May, 1989).
Experiments have been conducted to compare selenium from sodium selenite and from Sel-Plex® on feathering in broiler chickens (Edens, 1996; 2001).
It was determined that organic selenium induced more rapid feathering in auto-sexing, slow feathering male chickens and in their normal feathering sisters (Edens, 2001). An evaluation of thyroid hormone levels in broiler chickens was made in an attempt to relate thyroid hormones to feathering (Edens, 2001).
When organic selenium in Sel-Plex® was fed to broilers, the circulating percentage concentrations of T4 were lower and T3 concentrations were higher than in broilers given sodium selenite (Table 7). These observations suggest that organic selenium is more important than inorganic selenium in the early feathering of chickens and possibly other birds.
Table 7.Mean serum thyroxin (T4, ng/mL), triiodothyronine (T3, ng/mL), and percentage levels in Sel-Plex®-fed, Sel-Plex® + selenitefed and no supplemental selenium compared with the control (selenite) over a six week growing period.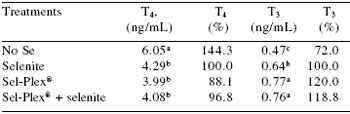
a,b,c In a column, means with unlike superscripts differ (P≤0.05).
Edens (2001) reported that body weight of broilers fed organic selenium in Sel-Plex® was improved (70 g/bird and 20 g/bird, respectively) over that of broilers fed no supplemental selenium and those fed sodium selenite.
Feed conversions of broilers fed organic selenium in Sel-Plex® was also improved (9 points and 3 points, respectively) over that of broilers fed no supplemental selenium and those fed sodium selenite. Part of the improvement in performance was due to the status of the metabolically active thyroid hormone in the broilers fed Sel-Plex® (Table 7).
These observations were supported by the report of Jianhua et al. (2000), who examined hepatic 5´-deiodinase activity and thyroid hormone dynamics in broilers fed selenium deficient or selenium (selenite) adequate diets. They concluded that selenium deficiency depressed growth of broilers by inhibiting 5´- deiodinase activity, which in turn decreased the plasma concentration of T3.
The role of selenium in the immune system of chickens has received little interest to date. We have noted that fast growing, high yielding broilers fed selenite at 1.2 ppm Se showed early signs of selenium toxicity by exhibiting decreased body weight, increased liver weight, and a compromised immune system as indicated by decreased thymus and bursa of Fabricius weights and abnormally high delayed type hypersensitivity reactions to phytohemagglutinin antigen (PHA-P), but broilers fed Sel-Plex® at 1.2 ppm were not affected (Gowdy and Edens, 2003).
Marsh et al. (1986) fed low levels of sodium selenite to chickens and observed increased growth of lymphoid organs including an increased thymus weight. Gowdy and Edens (2003) found that feeding sodium selenite (0.3 or 0.6 ppm) to control chicks also caused increased thymus weight compared to thymus weight from chicks fed either no selenium or organic selenium, but with infection, only organic selenium (0.3 or 0.6 ppm) maintained thymus weight comparable to weights of non-infected chicks.
Gowdy and Edens (2003) also reported that the chicken Tcell- mediated wing web response to PHA-P in Sel-Plex®-fed broilers was less than the response in sodium selenite-fed birds. Presumably, the lesser delayed type hypersensitivity response associated with Sel-Plex® feeding was associated with increased GSH-Px activity in lymphocytes and granulocytes that migrate to the site of antigen stimulation (Brown et al., 2000).
Arthur et al. (2003) reported that selenium is essential in the activation of all elements of the immune system. The thymus seems to have a special affinity for selenium because it is the location for maturation of T cells that ultimately will control most aspects of the immune system. The thymus, at least in mammals, also contains selenium-dependent Type II deiodinase enzyme, which is used to locally convert T4 to T3 and rT3 to T2.
Thus, it is highly probable that any impairment of this enzyme could have effects on T cells and the immune system in diverse locations throughout the body of an animal. The thymus appears to be exquisitely sensitive to selenium as shown by the results of Gowdy and Edens (2003); and it appears that organic selenium is better tolerated than inorganic selenium in the thymus and possibly mature T cells.
Leng et al. (2003) compared the influence of either sodium selenite or organic selenium sources on immunity in layer chickens. They determined that organic selenium increased levels of tissue selenium more than 2-fold higher than with selenium derived from selenite.
More importantly, Leng et al. (2003) were the first to show that the organic selenium caused an increase in CD3+, CD4+, and CD8+ surface markers on T cells located in several lymphoid structures in young chickens. The CD3+ marker is involved in signal transduction and is found in all T cells. The CD4+ marker is related to helper T cellmediated expansion of the humoral immune response ascribed solely to B cells from the bursa of Fabricius, and the CD8+ T cell marker is associated with cytotoxicity and cell killing functions.
Leng et al. (2003) concluded that organic selenium improved the status of the avian immune system by increasing the rate and ability of immunocompetent cells to respond to antigen challenge. They concluded that chickens fed organic selenium had more protection against potential pathogens than did chickens fed selenite. Chang et al. (1994) studied the influence of sodium selenite and vitamin E on T and B cell markers and lymphocyte proliferation. Lymphocyte proliferation was impaired by vitamin E and selenium deficiency, but the CD4+, CD8+ T cell populations increased slightly with vitamin E and selenium deficiency.
The relationship between the immune system and performance in poultry species is complex. It is generally believed that a heightened status of the active immune system is correlated with decreased performance. In fact, that situation can become established because an active immune system has higher energetic demands and nutrients are recompartmentalized to provide the energy required by the immunoactive cells. Increased demand from immunoactive cells decreases the nutrients that normally would be destined for protein production associated with growth.
In animals, with an immune system conditioned to react quickly to an antigen challenge and clear that antigen more quickly, performance can actually be enhanced due to less time utilized in combating the numerous challenges poultry face every day in their specific environments.
These activities might be enhanced in animals given organic selenium in Sel-Plex® as compared with animals fed inorganic sodium selenite. Evidence presented by Gowdy and Edens (2003) seem to support this hypothesis.
It is apparent that selenium-dependent 5´-deiodinase activity in poultry has a very important role to play.
On the most fundamental basis, embryonic growth, livability and hatchability are dependent on availability of selenium from the dam, and provision of organic selenium to the dam improves all of these parameters for the embryo (Edens, 2002).
Hatchability of chicken and turkey embryos is under the influence of thyroid hormones (Christensen and Biellier, 1982; Christensen, 1985). Christensen et al. (2002) determined that delayed conversion of T4 to T3 was associated with increased embryonic mortality and delayed hatchability.
However, if T3 is elevated prior to certain critical periods, embryonic livability and hatchability can also be decreased (Christensen et al., 2002). Van der Geyten et al. (1997) have reported that deiodinase 1 (D1) and deiodinase 3 (D3) in chicken embryos develop in parallel to the plasma T3 profiles.
D1 increased from day 14 of incubation to hatching. D3 increased from day 14 of incubation until day 17 of incubation then decreased. These enzymatic activities would indicate that there is maximized plasma T3 concentrations at the time of internal pipping and conversion to lung breathing by the embryo.
More recently, an unpublished report from the poultry industry indicated that hatchability of turkey eggs from Sel-Plex® -fed hens had a higher hatch rate if they had to be stored for as long as 10 to 14 days.
As pointed out by Edens (1996; 2001), broiler performance is improved with the use of organic selenium. Much of the explanation for the improved performance has focused on improved antioxidant status in chicks and poults from organic seleniumfed hens. Furthermore, post-slaughter yield is improved with the use of organic selenium, and water retention as indicated by less drip loss is improved.
Because it appears that T3 is very intimately involved in these responses, one must conclude that selenium as organic selenium has a special role to play in the regulation of poultry selenium-dependent monodeiodinases that convert T4 to T3 and rT3 to T2.
Thioredoxin reductase in chickens
Thioredoxin reductase (TrxR) belongs to a superfamily of flavoenzyme disulfide oxidoreductases including glutathione reductase, mercuric ion reductase, dihydrolipoamide reductase, and alkyl hydroperoxide reductase (Williams, 1992). This selenium dependent enzyme has been reported to show increased activity after selenium supplementation to animals as compared with enzyme activity in selenium deficient animals (Hill et al., 1997; Berggren et al., 1999).
However, Ganther and Ip (2001) have reported that monomethylated selenium in vivo did not increase TrxR activity and that high concentrations of selenium in a cell-free system inhibited TrxR. Wu et al. (2003) reported that there was an inverse relationship between dietary selenium and the activity and expression of TrxR in rat aorta.
Wu et al. (2003) suggested that the mechanisms regulating transcription of GSH-Px and TrxR in the aorta are different. Wu and colleagues suggested that GSH-Px expression and activity are directly related to selenium intake, but TrxR activity is mediated mostly by generation of ROM. In long-term selenium deficiency studies by Wu et al. (2003), TrxR activity actually increased, selenium repletion of the rats caused a decrease in TrxR activity, and selenium adequate animals had the lowest activity of TrxR.
The decreased activity of TrxR in the aorta of selenium-repleted animals might be a tissue specific response, because in other organs/tissues selenium deficiency caused a decrease in TrxR and GSH-Px activities (Berggren et al., 1999).
Berggren et al. (1999) stated that TrxR behaved in a manner very different from other selenoenzymes in that it increased its activity with excess sodium selenite and that the increased activity was transitory eventually returning to lower activities even with continued high dosing of selenium. Therefore, these observations might indicate that TrxR is induced as a safety precaution when its selenocysteine is affected by severe oxidative stress (Gladyshev et al., 1999).
In mammalian fast growing transformed tumor cells with high generation of ROM, thioredoxin and TrxR activities are much higher than in normal cells (Liu and Stadtman, 1997; Gladyshev et al., 1998; Ganther, 1999). There is an inverse relationship between TrxR and GSH-Px activity in tumor cells as compared with normal tissues or cell lines (Gladyshev et al., 1998).
Thioredoxin reductase functions to stabilize disulfide bonds (-S-S-), free sulfhydryl groups (-SH), and to reduce thioredoxin. In the extracellular environment, TrxR interacts with disulfide bonds on cell surfaces and extracellular proteins. In the cytosol of the cell, TrxR primarily stabilizes the sulfhydryl groups and reduces thioredoxin.
Thioredoxin is required inside the cell to provide reducing equivalents for many different substrates, primarily proteins that are vital to the survival of the cell. When thioredoxin undergoes oxidation, it is able to transfer reducing power to cellular proteins through TrxR, and TrxR, using electrons from NADPH, then reduces thioredoxin. All three mammalian TrxRs are selenium-dependent flavoproteins (Tamura and Stadtman, 1996).
Thioredoxin, the primary substrate for thioredoxin reductase, is expressed differentially in chickens as a product of a single copy gene and is similar to thioredoxins in other species (Jones and Luk, 1988).
Thioredoxin is a high capacity electron donor for reductive enzymes that include ribonucleotide reductase, thioredoxin peroxidase, and through thiol/ disulfide exchange it reduces cysteine residues in transcription factors to increase their binding to DNA thereby influencing gene transcription (Mustacich and Powis, 2000; Powis and Montfort, 2001).
Thioredoxin also functions as a cell growth factor and inhibits apoptosis. Arner and Holmgren (2000) report that thioredoxin reduces hydroperoxides, ascorbate and selenite. Ascorbate is the biochemical link between vitamin E and selenium because it has been shown to recycle tocopheroxyl to tocopherol in vitro (Burk and Hill, 1999). The thioredoxin-thioredoxin reductase system, which maintains free sulfhydryls, and the GSH/GSH-Px system, which is a primary antioxidant system, work together to regulate a low intracellular redox potential (Arner and Holmgren, 2000).
The TrxR work in chickens has just begun, and preliminary observations suggest that chicken TrxR may be different from mammalian TrxR (Gowdy and Edens, unpublished). Some of our preliminary observations on TrxR are presented in Tables 8-10, representing the first observations on tissue and cellular distributions in chicken and the influence of different selenium sources on its activity in various tissues.
Organ distribution and activity of TrxR in 3-week old broiler cockerels from our current studies are presented in Table 8. Similar to the observations made by Berggren et al. (1999) with the rat, we observed that broiler chickens which were not provided supplemental selenium in their diets had significantly lower TrxR activities than broilers given either sodium selenite or Sel-Plex®.
A second observation in this distribution study was that the selenium source was important in regulating TrxR activity. With few exceptions, TrxR activity was higher in Sel-Plex®- fed cockerels than in sodium selenite-fed or combination (selenite + Sel-Plex®)-fed birds. In comparison to mammals, chickens have low TrxR activities (Smith and Levander, 2001), and the low activities found in these chickens reflect the fact that TrxR levels in normal tissues are very low (Liu and Stadtman, 1997).
Table 8.Thioredoxin reductase activity in chicken tissues from 3- week-old male broilers fed supplemental levels of selenite, Sel- Plex®, a combination of 0.15 ppm selenite + 0.15 ppm Sel-Plex®, or no supplemental selenium 1.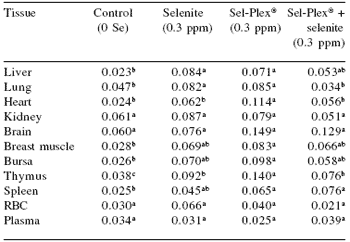
1TrxR activity (at 412 nm) was determined using the DTNB assay that measures the NADPH-dependent reduction of the disulfide bond in 5,5´-dithiobis(2-nitrobenzoic acid). Calculated results are based on yield of 2 moles of 2-nitro-5-thiobenzoate per mole of NADPH consumed. Results are given as the μmol of NADPH oxidized per minute per mg of total protein.
a,b,cIn a row, means with unlike superscripts differ significantly (P≤0.05).
The subcellular distribution of TrxR in chicken hepatic cells is presented in Table 9. The relatively high TrxR activity in a liver homogenate (primarily cytoplasmic) indicates the importance of the cellular content of the enzyme. When different fractions of the cell were analyzed for TrxR activity, the nuclear pellet and the mitochondrial lysate had the highest activities followed by post-mitochondrial supernatant, mitochondrial membranes, post-nuclear supernatant, and the mitochondrial pellet.
Rigobello et al. (1998) reported that total TrxR activity of the mitochondrial pellet was higher than the mitochondrial matrix (lysate) in rat liver cells, but when specific activity was determined, the matrix TrxR activity was higher than that in the mitochondrial pellet, similar to the observation made in this investigation with chickens (Table 9). The post-nuclear and post-mitochondrial supernatant TrxR activities were comparable to the cytosolic activity because those post-supernatants are comprised of the cytosol.
The observations with subcellular distribution of TrxR in chickens are similar to observations made in rat (Rozell et al., 1985;1988) and human tissue (Ejima et al., 1999; Chen et al., 2002). Rozell et al. (1985;1988) concluded that the distribution of thioredoxin and TrxR to subcellular structures such as the endoplasmic reticulum, secretory granules, plasma membrane, and at the subplasma membrane was consistent with the functions in protein processing, secretion, and formation of nascent protein disulfides.
Chen et al. (2002) found high concentrations of TrxR in the mitochondria, lysosome, microsome and cytosol in human liver, and Ejima et al. (1999) found high TrxR activity (90%) in the cytosol of the placenta cells and about 10% of the total cellular activity in mitochondria. An important deduction from these observations is that TrxR plays a significant role in the nucleus and in the mitochondria. The role of TrxR in the nucleus should be apparent when it is understood that thioredoxin is used to enhance transcription of DNA (Mustacich and Powis, 2000; Powis and Montfort, 2001).
Table 9.Subcellular distribution of chicken thioredoxin reductase activity (μmol NADPH/min/mg total protein) in liver cells.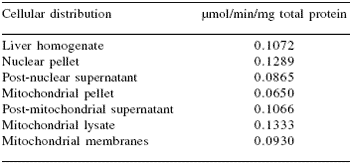
TrxR activity (at 412 nm) was determined using the DTNB assay that measures the NADPH-dependent reduction of the disulfide bond in 5,5´-dithiobis(2-nitrobenzoic acid). Results are given as the μmol of NADPH oxidized per minute per mg of total protein.
We also wanted to determine the influence of feeding high levels of sodium selenite and Sel-Plex® on activity of TrxR in chicken liver homogenates (Table 10). This was part of a larger study in which the toxicity of Sel-Plex® was being evaluated in chickens (Gowdy and Edens, 2003).
In that study, we determined that as little as 1.2 ppm of sodium selenite was sufficient to allow signs of toxicity to develop in high-yield broiler chickens. At 5 ppm sodium selenite caused severe growth depression, and at 15 ppm all of the chickens fed sodium selenite had experienced severe growth depression and nearly all had died by two weeks of age.
Contrary to those observations, there was no treatment-associated mortality in the Sel-Plex®-fed groups and body weights were comparable to controls fed 0.3 ppm Se. At two weeks of age, the chickens were killed by carbon dioxide asphyxiation and liver samples were collected from each bird and analyzed for TrxR (Table 10).
It was interesting to observe that chickens given no supplemental selenium had the lowest liver TrxR activity. Feeding either sodium selenite or Sel-Plex® from 0.3 ppm to 15 ppm induced higher activities of TrxR, and Sel-Plex® at 15 ppm Se induced the highest activity of TrxR. This observation was similar to that of Berggren et al. (1999) who also reported selenium (as selenite) to be capable of inducing in vivo higher TrxR activity in rats. Thus, chickens and rats respond comparablely with selenium-mediated induction of TrxR.
Table 10. Thioredoxin reductase activity (μmol NADPH/min/mg total protein) from chicken liver homogenates from 3 week old male broilers fed high levels of selenite or Sel-Plex® 1.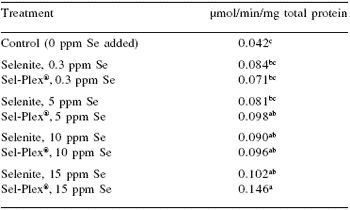
1TrxR activity (at 412 nm) was determined using the DTNB assay that measures the NADPH-dependent reduction of the disulfide bond in 5,5´-dithiobis(2-nitrobenzoic acid). Calculated resulted are based on yield of 2 moles of 2-nitro-5-thiobenzoate per mol of NADPH consumed. Results are given as the μmol of NADPH oxidized per minute per mg of total protein. a,b,cMeans with unlike superscripts differ significantly (P≤0.05).
The importance of thioredoxin reductase in living systems, such as the growing chicken, continues to be revealed.
Currently, it is believed that TrxR functions to reduce thioredoxin and maintain a readily available supply of the reduced thioredoxin, which has numerous functions associated with growth and development.
Second, TrxR functions to maintain cytosolic protein sulfhydryls and disulfide bonds in a reduced state.
Third, TrxR has antioxidant properties that come into play when the redox potential becomes more negative due to rapid and large production of ROM in cells. In that capacity, TrxR acts in a manner similar to GSH-Px as an antioxidant against ROM.
Without these general functions, animals such as chickens and other food animal species would suffer from oxidative stress and become less productive.
Summary
• Selenium is essential in all animals.
• Organic selenium is the preferred form and has higher bioavailability than inorganic selenium. • Sodium selenite is a pro-oxidant but selenomethionine is not.
• Feed supplements providing organic selenium are not the same; exercise caution in making selection.
• Selenium is unique among all trace minerals, requiring its own mRNA and SECIS.
• Selenomethionine in body protein provides reversible storage of selenium in tissues and organs.
• Selenomethionine is integrated into body protein, but inorganic selenium is loosely associated with thiols via a labile bond.
• Selenium must function through specific selenoproteins (25 known) encoded by at least 18 genes.
• Poultry express selenoproteins, but all avian selenoproteins are not yet described.
• Poultry are known to express selenium-dependent GSH-Px, iododeiodinase, and TrxR.
• Performance as indicated by body weight, livability, feed conversion, meat yield, and meat quality in poultry is influenced significantly by selenoproteins.
• Physiological functions as diverse as body temperature regulation, immunity, resistance to oxidative stress, reproduction, thyroid function, growth, development, all endocrine functions, digestive processes associated with pancreatic function, and more are influenced by selenium status in poultry.
References
Allen, C.B., G.M. Lacourciere and T.C. Stadtman. 1999. Responsiveness of selenproteins to dietary selenium. Annu. Rev. Nutr. 19:1-16.
Amberg, R., T. Mizutani, X.Q. Wu and H.J. Gross. 1996. Selenocysteine synthesis in mammalia: an identity switch from tRNA(Ser) to tRNA(SEC). J. Mol. Biol. 263:8-19.
Arner, E.S.J. and A. Holmgren. 2000. Physiological functions of thioredoxin reductase. Eur. J. Biochem. 267:6102-6109.
Arthur, J.R. 1997. Non-glutathione peroxidase functions of selenium. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 13th Annual Symposium (K.A. Jacques and T.P Lyons, eds). Nottingham University Press, UK, pp. 143-154.
Arthur, J.R., R.C McKenzie and G.J. Beckett. 2003. Selenium in the immune system. J. Nutr. 133:1457S-1459S.
Arthur, J.R., F. Nicol and G.J. Beckett. 1990. Hepatic iodothyronine 5’-deiodinase. The role of selenium. Biochem. J. 272:537-540.
Avanzo J.L., C.X. Mendonca, S.M. Pugine and M.C. Cesar. 2001. Effect of vitamin E and selenium on resistance to oxidative stress in chicken superficial pectoralis muscle. Comp. Biochem. Physiol. C 129:163-173.
Behne, D., H. Hilmert, S. Scheid, H. Gessner and W. Elger. 1988. Evidence for specific selenium target tissues and new biologically important selenoproteins. Biochim. Biophys. Acta 966:12-21.
Behne, D., A. Kyriakopoulos, H. Meinhold and J. Köhrle. 1990. Identification of type-I iodothyronine 5’-deiodinase as a selenoenzyme. Biochem. Biophys. Res. Commun. 173:1143-1149.
Behne, D., A Kyriakopoulos, M. Kalcklösch, C. Weiss-Nowak, K. Pfeifer, H. Gessner and C. Hammel. 1997. Two new selenoproteins found in the prostatic glandular epithelium and in the spermatid nuclei. Biomed. Environ. Sci. 10:340- 345.
Berggren, M.M., J.F. Mangin, J.R. Gasdaska and G. Powis. 1999. Effect of selenium on rat thioredoxin reductase activity. Biochem. Pharmacol. 57:187- 193.
Berry, M.J., L. Banu and P.R. Larsen. 1991a. Type I iodothyronine deiodinase is a selenocysteinecontaining enzyme. Nature 349:438-440.
Berry, M.J., L. Banu, Y. Chen, S.J. Mandel, J.D. Kieffer, J.W. Harney and P.R. Larsen. 1991b. Recognition of UGA as a selenocysteine codon in Type I deiodinase requires sequences in the 3’ untranslated region. Nature 353:273-276.
Berry, M.J., L. Banu, J.W. Harney and P.R. Larsen. 1993. Functional characterization of the eukaryotic SECIS elements which direct selenocysteine insertion at UGA codons. EMBO J. 12:3315-3322.
Boone, M.A., J.A. Davidson and E.P. Reineke. 1950. Thyroid studies in fast- and slow- feathering Rhode Island Red chicks. Poult. Sci. 29:195-200.
Brown, K.M., K. Pickard, F. Nicol, G.J. Beckett, G.G. Duthie and J.R. Arthur. 2000. Effects of organic and inorganic selenium supplementation on selenoenzyme activity in blood lymphocytes, granulocytes, platelets and erythrocytes. Clin. Sci. 98:593-599.
Brtko, A., J. Bransova, P. Filipcik, S. Hudecova and A. Brtkova. 1997. Nuclear receptors-ligand inducible transcription factors: effects of selenium. Biomark. Env. 1:6-7.
Burk, R.F. 1976. Selenium in Man. In: Trace Elements in Human Health and Disease (A.S. Prasad and D. Oberleas, eds). Academic Press, New York, pp. 105- 133.
Burk, R.F. 1991. Molecular biology of selenium with implications for its metabolism. FASEB J. 5:2274- 2279.
Burk, R.F. and K.E. Hill. 1999. Orphan selenoproteins. BioEssays 21 (3):231-237.
Butler, G.W. and P.J. Peterson. 1967. Uptake and metabolism of inorganic forms of selenium-75 by Spiroldela oligorrhiza. Aust. J. Biol. Sci. 20: 77.l
Cai, X., E. Block, P. C. Uden, X. Zhang, B.D. Quimby and J.J. Sullivan. 1995. Allium Chemistry: Identification of selenoamino acids in ordinary and selenium-enriched garlic, onion and broccoli using gas chromatography with atomic emission detection. J. Agric. Food Chem. 43:1754-1757.
Cantor, A.H. and T.H. Johnson. 1985. Influence of dietary protein sequence and selenium upon development of pullets. Poult. Sci. 64(Suppl. 1):75 (Abstr.).
Cantor, A.H. and J.Z. Tarino. 1982. Comparative effects of inorganic dietary sources of selenium on selenium levels and selenium-dependent glutathione peroxidase activity in blood of young turkeys. J. Nutr. 112:2187-2196.
Cantor, A.H., M.L. Scott and T. Noguchi. 1975a. Biological availability of selenium in foodstuffs and selenium compounds for prevention of exudative diathesis in chicks. J. Nutr. 105:96-105.
Cantor, A.H., M.L. Langevin, T. Naguchi and M.L. Scott. 1975b. Efficacy of selenium in selenium compounds and foodstuffs for prevention of pancreatic fibrosis in chicks. J. Nutr. 105:106-111.
Cantor, A.H., P.D. Moorhead and M.A. Musser. 1982. Comparative effects of sodium selenite and selenomethionine upon nutritional muscular dystrophy, selenium-dependent glutathione peroxidase and tissue selenium concentrations of turkey poults. Poult. Sci. 61:478-484.
Catellano, S., S.V. Novoselov, G.V. Kryukov, A. Lescure, E. Blanco, A. Krol, V.N. Gladyshev and R. Guigo. 2004. Reconsidering the evolution of eukaryotic selenoproteins: a novel nonmammalian family with scattered phylogenetic distribution. EMBO Reports 5:71-77.
Chen, C.J. Zhao, P. Zhang and Z. Chai. 2002. Speciation and subcellular location of Se-containing proteins in human liver studied by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and hydride generation-atomic fluorescence spectrometric detection. Anal. Bioanal. Chem. 372:426-430.
Chang, W.P., J.S. Hom, R.R. Dietert, G.F. Combs, Jr. and J.A. Marsh. 1994. Effect of dietary vitamin E and selenium deficiency on chicken splenocyte proliferation and cell surface marker expression. Immunopharmacol. Immunotoxicol. 16:203-223.
Christensen, V.L. 1985. Supplemental thyroid hormones and hatchability of turkey eggs. Poult. Sci. 64:2202-2210.
Christensen, V.L., G. S. Davis and K. E. Nestor. 2002. Environmental incubation factors influence embryonic thyroid hormones. Poult. Sci. 81:442- 450.
Christensen, V.L. and H.V. Biellier. 1982. Physiology of turkey embryos during pipping and hatching. IV. Thyroid function in embryos from selected hens. Poult. Sci. 61:2482-2488.
Chu, F.F., J.H. Doroshow and R.S. Esworthy. 1993. Expression, characterization and distribution of a new cellular selenium-dependent glutathione peroxidase, GSHpx-GI. J. Biol. Chem. 268:2571- 2576.
Croteau, W., S.L. Whittemore, M.J. Schneider and D.L. St. Germain. 1995. Cloning and expression of a cDNA for a mammalian type-III iodothyronine deiodinase. J. Clin. Invest. 270:16569-14575.
Croteau, W., J.C. Davey, V.A. Galton and D.L. St. Germain. 1996. Cloning of the mammalian type- II iodothyronine deiodinase. J. Clin. Invest. 98:405- 417.
Csallany, A.S. and B.Z. Menken. 1986. Effect of dietary selenite on hepatic organic solvent-soluble lipofuscin pigments. J. Am. Coll. Toxicol. 5:79- 85.
Cummins, L.M. and J.L. Martin. 1967. Are selenocystine and selenomethionine synthesized in vivo from sodium selenite in mammals? Biochem. 6:3162-3168.
Daniels, L.A. 1996. Selenium metabolism and bioavailability. Biol. Trace Elem. Res. 54: 185- 199.
Douglas, J.S., V.C. Morris, J.H. Soares and O.A. Levander. 1981. Nutritional availability to rats of selenium in tuna, beef kidney and wheat. J. Nutr. 111:2180-2187.
Echevarria, M.G., P.R. Henry, C.B. Ammerman, P.V. Roa and R.D. Miles. 1988a. Estimation of the relative bioavailability of inorganic selenium sources for poultry. 1. Effect of time and high dietary selenium on tissue selenium uptake. Poult. Sci. 67:1295-1301.
Echevarria, M.G., P.R. Henry, C.B. Ammerman, P.V. Roa and R.D. Miles. 1988b. Estimation of the relative bioavailability of inorganic selenium sources for poultry. 2. Tissue uptake of selenium from high dietary selenium concentrations. Poult. Sci. 67:1585-1592.
Edens, F.W. 1996. Organic selenium: From feathers to muscle integrity to drip loss. Five years onward: No more selenite! In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 12th Annual Symposium (K.A. Jacques and T.P Lyons, eds). Nottingham University Press, UK, pp. 165-185.
Edens, F.W. 2001. Involvement of Sel-Plex in physiological stability and performance of broiler chickens. In: Science and Technology in the Feed Industry, Proceedings of Alltech’s 17th Annual Symposium (K.A. Jacques and T.P Lyons, eds). Nottingham University Press, UK, pp. 349-376.
Edens, F.W. 2002. Practical Applications for Selenomethionine: Broiler Breeder Reproduction. In: Nutritional Biotechnology in the Feed and Food Industry, Proceedings of Alltech’s 18th Annual Symposium (K.A. Jacques and T.P Lyons, eds). Nottingham University Press, UK, pp. 29-42.
Edens, F.W. and A.E. Sefton. 2002. Selenomethionine supplementation to diets of broiler breeders improves performance. Poult. Sci. 80(Suppl.1):91.
Edens, F.W., C.R. Parkhurst and A.E. Sefton. 2002. Selenomethionine from selenized yeast improves spermatozoal integrity. Poult. Sci. 80(Suppl.1):90.
Ejima, K., H. Nanri, N. Toki, M. Kashimura and M. Ikeda. 1999. Localization of thioredoxin reductase and thioredoxin in normal human placenta and their protective effect against oxidative stress. Placenta. 20:95-101.
Encinar, J.R., M. Sliwka-Kaszynska, A. Polatajko, V. Vacchina and J. Szpunar. 2003. Methodological advances for selenium speciation analysis in yeast. Analyt. Chim. Acta 500:171-183.
Esaki, N., T. Nakamura, H. Tanaka, T. Suzuki, Y. Morino and K. Soda. 1981. Enzymatic synthesis of selenocysteine in rat liver. Biochem. 20:4492- 4496.
Federal Register, June 6, 2000. Food additive permitted in feed and drinking water: Selenium yeast. Fed. Reg. 65(109):35823-35824.
Federal Register, July 17, 2002. Food additive permitted in feed and drinking water: Selenium yeast. Fed. Reg. 67(137):46850-46851.
Flohé, L., W.A. Gunzler and H.H. Schock. 1973. Glutathione peroxidase. A selenoenzyme. FEBS Lett. 32:132-134.
Fratelli, M., H. Demol, M. Puype, S. Casagrande, I. Eberini, M. Salmona, V. Bonetto, M. Mengozzi, F. Duffieux, E. Miclet, A. Bachi, J. Vanderkerckhove, E. Gianazza and P. Ghezzi. 2002. Identification by redox proteomics of glutathionylated proteins in oxidatively stressed human T lymphocytes. Proc. Natl. Acad. Sci., USA 99:3505-3510.
Gabrielsen, J.O. and J. Opstevedt. 1980. Availability of selenium in fish meal in comparison with soybean meal, corn gluten meal and selenomethionine relative selenium in sodium selenite for restoring glutathione peroxidase activity in selenium depleted chicks. J. Nutr. 110:1096- 1100.
Ganther, H.E. 1999. Selenium metabolism, selenoproteins and mechanisms of cancer prevention: complexities with thioredoxin reductase. Carcinogenesis. 20:1657-1666.
Ganther, H.E. and C. Ip. 2001. Thioredoxin reductase activity in rat liver is not affected by supranutritional levels of monmethylated selenium in vivo and is inhibited only by high levels of selenium in vitro. J. Nutr. 131:310-304.
Gasdaska, P.Y., M.M. Berggren, M.L. Berry and G. Powis. 1999. Cloning, sequencing and functional expression of a novel human thioredoxin reductase. FEBS Letters 442:105-111.
Gladyshev, V.N., Q-A. Sun, Y.L. Wu, F. Zappacosta, K-T. Jeang, B.J. Lee and D.L. Hatfield. 1999. Redox regulation of cell signaling by selenocysteine in mammalian thioredoxin reductases. FASEB J. 13:A1579.
Gladyshev, V.N., V.M. Factor, F. Housseau and D.L. Hatfield. 1998. Contrasting patterns of regulation of the antioxidant selenoproteins, thioredoxin reductase and glutathione peroxidase, in cancer cells. Biochem. Biophys. Res. Commun. 251:488- 493.
Gowdy, K.M. and F.W. Edens. 2003. Organic selenium affects broiler responses to immunostimulation. Poult. Sci. 82(Suppl. 1):10.
Grimaud, R., B. Ezraty, J.K. Mitchell, D. Lafitte, C. Briand, P.J. Derrick and F. Barras. 2001. Repair of oxidized proteins. Identification of a new methionine sulfoxide reductase of type B:hCBS1. FEBS Lett. 527:91-100.
Hafeman, D.G., R.A. Sunde and W.G. Hoekstra. 1974. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J. Nutri. 104: 580-587.
Hatfield, D.L. and V.N. Gladyshev. 2003. How selenium has altered our understanding of the genetic code. Mol. Cell. Biol. 22:3565-3576.
Hawkes, W.C., E.C. Wilhelmsen and A.L. Tappel. 1985a. Abundance and tissue distribution of selenocysteine-containing proteins in the rat. J. Inorg. Biochem. 23:77-92.
Hawkes, W.C., E.C. Wilhelmsen and A.L. Tappel. 1985b. Subcellular distribution of seleniumcontaining proteins in the rat. J. Inorg. Biochem. 25:77-93.
Hawles, W.C., F.Z. Alkan and L. Oehler. 2003. Adsorption, distribution and excretion of selenium from beef and rice in healthy North American men. J. Nutr. 133:3434-3442.
Herrman, J.L. 1977. The properties of a rat serum protein labeled by the injection of sodium selenite. Biochim. Biophys. Acta 500:61-70.
Hill, K.E., G.W. McCollum, M.E. Boeglin and R.F. Burk. 1997. Thioredoxin reductase activity is decreased by selenium deficiency. Biochem. Biophys. Res. Commun. 234:293-295.
Ilian, M.A. and P.D. Whanger. 1989. In vitro metabolism of 75Se-selenite and 75Seselenomethionine in chick blood. J. Trace Elem. Electrolytes Health Dis. 3:9-16.
Jacques, K.A. 2001. Selenium metabolism in animals: the relationship between dietary selenium form and physiological function. In: Science and Technology in the Feed Industry, Proceedings of Alltech’s 17th Annual Symposium (K.A. Jacques and T.P Lyons, eds). Nottingham University Press, UK, pp. 319-348.
Jianhua, H., A. Ohtsuka and K. Hayashi. 2000. Selenium influences growth via thyroid hormone status in broiler chickens. Brit. J. Nutr. 84:727- 732.
Jones, S.W. and K-C. Luk. 1988. Isolation of a chicken thioredoxin cDNA clone. J. Biol. Chem. 263:9607-9611.
Kaplan, M.M. 1986. Regulatory influences on iodothyronine deiodinase in animal tissues. In: Thyroid Hormone Metabolism (G. Hennemann, ed). Marcel Dekker, New York, pp. 231-251.
Kelly, M.P. and R.F. Power. 1995. Fractionation and identification of the major selenium compounds in selenized yeast. J. Dairy Sci. 78(Suppl. 1):237.
Kong, B.W., H. Kim and D.N. Foster. 2003. Cloning and expression analysis of chicken phospholipidhydroperoxide glutathione peroxidase. Anim. Biotechnol. 14:19-29.
Kryukov, G.V., S. Castellano, S.V. Novoselov, A.V. Lobanov, O. Zehtab, R. Gulgó and V.N. Gladyshev. 2003. Characterization of mammalian selenoproteomes. Science 300:1439-1443.
Kyriakopoulos, A., C. Hammel, H. Gessner and D. Behne. 1996, Characterization of an 18 kDaselenium containing protein in several tissues of the rat. Am. Biotech. Lab. 14:22.
Latshaw, J.D. and M. Osman. 1974. A selenium and vitamin E responsive condition in the laying hen. Poult. Sci. 53:1704-1708.
Lee, S.R., J.R. Kim, K.S. Kwon, H.W. Yoon, R.L. Levine, A. Ginsberg and S.G. Rhee. 1999. Molecular cloning and characterization of a mitochondrial selenocysteine-containing thioredoxin reductase from rat liver. J. Biol. Chem. 274:4722-4734.
Leeson, S. and J.D. Summers. 1991. Commercial Poultry Nutrition. University Books, Guelph, Ontario, Canada.
Leng, L., R. Bobcek, S. Kuricová, K. Boldizárová, L. Grešáková, Z. Ševcíková, V. Révajová, M. Levkutová and M. Levkut. 2003. Comparative metabolic and immune responses of chickens fed diets containing inorganic selenium and Sel-PlexTM organic selenium. In: Nutritional Biotechnology in the Feed and Food Industry, Proceedings of Alltech’s 19th Annual Symposium (K.A. Jacques and T.P Lyons, eds). Nottingham University Press, UK, pp. 131-145.
Lescure, A., D. Gautheret, D.P. Carbon and A. Krol. 1999. Novel selenoproteins identified in silco and in vivo by using a conserved RNA structural motif. J. Biol. Chem. 274:38147-38154
Lescure, A., D. Gautheret, D. Fagegaltier, P. Carbon and A. Krol. 2000. From RNA structure to the identification of new genes: The example of selenoproteins. J. Hlth. Sci. 46:405-408.
Levander, O. 1986. Selenium. In: Trace Elements in Human and Animal Nutrition (W. Mertz, ed). Academic Press, Orlando, FL, pp. 209-280.
Levine, R.L., L. Mosoni, B.S. Berlett and E.R. Stadtman. 1996. Methionine residues as endogenous antioxidants in proteins. Proc. Natl. Acad. Sci., USA 93:15036-15040.
Liu, S-Y. and T.C. Stadtman. 1997. Heparin-binding properties of selenium-containing thioredoxin reductase from HeLa cells and human lung adenocarcinoma cells. Proc. Natl. Acad. Sci. USA 94:6138-6141.
Low, S.C. and M.J. Berry. 1996. Knowing when not to stop: Selenocysteine incorporation into eukaryotes. TIBS 21:203-208.
Low, S. C., J.W. Harney and M.J. Berry. 1995. Cloning and functional characterization of human selenophosphate synthatase, an essential component of selenoprotein synthesis. J. Biol. Chem. 270:21659-21664.
Mahan, D.C. 1994. Organic selenium sources for swine. How do they compare with inorganic selenium sources? In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 10th Annual Symposium (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, UK, pp. 323-333.
Mahan, D.C. 1995. Selenium metabolism in animals: What role does selenium yeast have? In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 11th Annual Symposium (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, UK, pp. 257-267.
Mahmoud, K.Z. and F.W. Edens. 2003. Influence of selenium sources on age-related and mild heat stressrelated changes of blood and liver glutathione redox cycle in broiler chickens (Gallus domesticus). Comp. Biochem. Physiol. B 136:921-934.
Maiorino, M., C. Roche, M. Kiess, K. Koenig, D. Gawlik, M. Matthes, E. Naldini, R. Pierce and L. Flohe. 1996. A selenium-containing phospholipidhydroperoxide glutathione peroxidase in Schistosoma mansoni. Eur. J. Biochem. 238:838- 844.
Marsh, J.A., G.F. Combs, Jr, M.E. Whitacre and R.R. Dietert. 1986. Effect of selenium and vitamin E dietary deficiencies on chick lymphoid organ development. Proc. Soc. Expt. Biol. Med. 182:425- 436.
May, J.D. 1989. The role of the thyroid in avian species. Crit. Rev. Poult. Biol. 2: 171-189. McConnell, K.P. and G. Cho. 1965. Transmucosal movement of selenium. Am. J. Physiol. 208:1191- 1195.
Mertz, W. 1987. Selenium from a distance. In: Selenium in Biology and Medicine. Part A (G.F. Combs, Jr., J.E. Spallholz, O.A. Levander and J.E. Oldfield, eds). Van Nostrand Reinhold Company, New York, New York, pp. 3-8.
Miller, D., J.H. Soares, Jr., P. Baursfeld, Jr. and S.L. Cuppett. 1972. Comparative selenium retention by chicks fed sodium selenite, selenomethionine, fish meal and fish solubles. Poult. Sci. 51:1669-1673.
Miranda-Vizuete, A., A.E. Damdimopoulos, J.R. Pedrajas, J.A. Gustafsson and G. Spyrou. 1999. Human mitochondrial thioredoxin reductase. Eur. J. Biochem. 261:405-412.
Mitch, W.E. and A.L. Goldberg. 1996. Mechanism of muscle wasting- The role of the ubiquitinproteosome pathway. N. Engl. J. Med. 335:1897- 1905.
Miyazaki, S. and Y. Motoi. 1992. Tissue distribution of monomeric glutathione peroxidase in broiler chicks. Res. Vet. Sci. 53:47-51.
Miyazaki, S. and Y. Motoi. 1996. Purification and characterization of chicken liver monomeric glutathione peroxidase. Br. Poult. Sci. 37:651-660.
Moksnes, K. and G. Norheim. 1986. A comparison of selenomethionine and sodium selenite as a supplement in chicken feeds. Acta Vet. Scand. 27:103-114.
Molinero, P., C. Osuna and J.M. Guerrero. 1995. Type II thyroxine 5’-deiodinase in rat thymus. J. Endocrinol. 146:105-111.
Mortimer, R.H., J.P. Galligan, G.R. Cannell, R.S. Addison and M.S. Roberts. 1996. Maternal to fetal thyroxine transmission in the human term placenta is limited by inner ring deiodination. J. Clin. Endocrinol. Metab. 81:2247-2249.
Moskovitz, J.S. Bar-Noy, W.M. Williams, J. Requena, B.S. Berlett and E.R. Stadtman. 2001. Methionine sulfoxide reductase (MsrA) is a regulator of antioxidant defense and lifespan in mammals. Proc. Natl. Acad. Sci. USA 98:12920- 12925.
Motsenbocker, M.A. and A.L. Tappel. 1982. A selenocysteine-containing selenium transport protein in rat blood. Biochim. Biophys. Acta 719:147-153.
Mustacich, D. and G. Powis. 2000. Thioredoxin reductase. Biochem. J. 346:1-8.
Okamoto, K., H. Tanaka, H. Ogawa, Y. Makino, H. Eguchi, N. Yoshikawa, L. Poellinger, K. Umesono and I. Makino. 1999. Redox dependent regulation of nuclear import of the glucocorticoid receptor. J. Biol. Chem. 274:10363-10371.
Ort, J.F. and J.D. Latshaw. 1978. The toxic level of sodium selenite in the diet of laying chickens. J. Nutr. 108:1114-1120.
Osman, M. and J.F. Latshaw. 1976. Biological potency of selenium from sodium selenite, selenomethionine and selenocysteine in the chick. Poult. Sci. 55:987-984.
Pablos, M.I., M.T. Agapito, R. Gutierrez, J. M. Recio, R.J. Reiter, L.R. Barlow-Walden, D. Acuna- Castroviejo and A. Menendez-Pelaez. 1995a. Melatonin stimulates the activity of the detoxifying enzyme glutathione peroxidase in several tissues of the chick. J. Pineal Res. 19:111-115.
Pablos, M.I., J.I. Chuang, R.J. Reiter, G.G. Ortiz, W.M. Daniels, E. Sewerynek, D. Melchiorri and B. Poeggeler. 1995b. Time course of melatonininduced increase in glutathione peroxidase activity in chick tissues. Biol. Signals 4:325-330.
Pablos, M.I., R.J. Reiter, G.G. Ortiz, J.M. Guerrero, M.T. Agapito, J-I. Chuang and E. Sewerynek. 1998. Rhythms of glutathione peroxidase and glutathione reductase in brain of chick and their inhibition by light. Neurochem. Int. 32:69-75.
Patterson, E.L., R. Milstrey and E.L.R. Stokstad. 1957. Effect of selenium in preventing exudative diathesis in chicks. Proc. Soc. Exp. Biol. Med. 95:617-620.
Petrovich, Iu’A. and R.P. Podorozhnaia. 1981. Selenoenzymes and other selenoproteins, their biological role. Usp. Sovrem. Biol. 91:127-144 (in Russian).
Powis, G. and W.R. Montfort. 2001. Properties and biological activities of thioredoxins. Annu. Rev. Biophys. Biomo. Struct. 30:421-455.
Previle, X., F. Salvemini, S. Giraud, S. Chaufour, C. Paul, G. Stephen, M.V. Ursini and A.P. Arrigo. 1999. Mammalian small stress proteins protect against oxidative stress through their ability to increase glucose-6-phosphate dehydrogenase activity and by maintaining optimal cellular detoxifying machinery. Exp. Cell Res. 247:61-78.
Radi, M.H. and D.C. Warren. 1938. Studies on the physiology and inheritance of feathering in growing chicks. J. Agricult. Res. 56:679-705.
Rigobello, M.P., M.T. Callegaro, E. Barzon, M. Benetti and A. Bindoli. 1998. Purification of mitochondrial thioredoxin reductase and its involvement in redox regulation of membrane permeability. Free Rad. Biol. Med. 24:370-376.
Ringdal, O., E.O. Bjornestad and K. Julshamn. 1985. Comparative utilization of fish selenium and inorganic selenite in rats of normal selenium status. Ann. Nutr. Metab. 29:297-305.
Rotruck, J.T., A.L. Pope, H.E. Ganther, A.B. Swanson, D.G. Hafeman and W.G. Hoekstra. 1973. Selenium: Biochemical role as a component of glutathione peroxidase. Science 179:588-590.
Rozell, B., H.A. Hansson, M. Luthman and A. Holmgren. 1985. Immunohistochemical localization of thioredoxin and thioredoxin reductase in adult rats. Eur. J. Cell Biol. 38:79-86.
Rozell, B., A. Holmgren and H.A. Hansson. 1988. Ultrastructural demonstration of thioredoxin and thioredoxin reductase in rat hepatocytes. Eur. J. Cell. Biol. 46:470-477.
Salavtore, D., T. Bartha, J.W. Harney and P.R. Larsen. 1996. Molecular, biological and biochemical characterization of the human type-2 selenodeiodinase. Endocrinol. 17:3308-3315.
Schen, L., K. van Dyke, J. Luten and H. Deelstra. 1997. Diffusibility of selenate, selenite, selenomethionine and seleno-cysteine during simulated gastrointestinal digestion. Biol. Trace Elem. Res. 58:55-63.
Schrauzer, G.N. 2001. Nutritionasl selenium supplements: Product types, quality and safety. J. Am. Coll. Nutr. 20:1-4.
Schrauzer, G.N. 2003. The nutritional significance, metabolism and toxicology of selenomethionine. Adv. Food Nutr. Res. 47:73-112.
Schwarz, K. 1951. Production of dietary necrotic liver degeneration using American Torula yeast. Proc. Soc. Exp. Biol. Med. 77:818-823.
Schwarz, K. 1954. Influence of food supply and fat intake on dietary necrotic liver degeneration. Fed. Proc., Fed. Amer. Soc. Exp. Biol. 13:477 (abstr).
Schwarz, K. and C.M. Foltz. 1957. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 79:3292- 3293.
Schwarz, K., J.G. Bieri, G.M. Briggs and M.L. Scott. 1957. Prevention of exudative diathesis in chicks by Factor 3 and selenium. Proc. Soc. Exp. Biol. Med. 95:621-625.
Scott, M.L. and J.N. Thompson. 1971. Selenium content in foodstuffs and effects of dietary selenium levels upon tissue selenium in chicks and poults. Poult. Sci. 50:1742-1748.
Smith, A.D. and O.A. Levander. 2001. Rapid determination of glutathione peroxidase and thioredoxin reductase activities using a 96-well microplate format: comparison to standard cuvettebased assays. Int. J. Vitam. Nutr. Res. 71:87-92.
Smith, A.D. and O.A. Levander. 2002. High throughput 96-well microplate assays for determining specific activities of glutathione peroxidase and thioredoxin reductase. Meth. Enzymol. 347:113-321.
Snityns’kyi, V.V. and H.L. Antoniak. 1994. Biochemical role of selenium. Ukr. Biokhim Zh. 66:3-16 (in Ukrainian).
Spallholz, J.E. 1997. Free radical generation by selenium compounds and their pro-oxidant toxicity. Biomed. Environ. Sci. 10:260-270.
Spencer, R.P. and M. Blau. 1962. Intestinal transport of selenium 75selenomethionine. Science 163:155- 156.
Stadtman, T.C. 1980. Selenium-dependent enzymes. Ann. Rev. Biochem. 49:93-110.
Stadtman, E.R., J. Moskovitz, B.S. Berlett and R.L. Levine. 2002. Cyclic oxidation and reduction of protein methionine residues is an important antioxidant mechanism. Mol. Cell. Biochem. 234:3-9.
Sun, Q-A., Y. Wu, F. Zappacosta, K-T. Jeang, B.J. Lee, D.L. Hatfield and V.N. Gladyshev. 1999. Redox regulation of cell signaling by selenocysteine in mammalian thioredoxin reductase. J. Biol. Chem. 274:24522-24530.
Sunde, R.A. 1990. Molecular biology of selenoproteins. Ann. Rev. Nutri. 10:451-474.
Surai, P.F., E. Blesbois, I. Grasseau, T. Chalah, J.P. Brillard, G.J. Wishart, S. Cerolini and N.H. Sparks. 1998a. Fatty acid composition, glutathione peroxidase and superoxide dismutase activity and total antioxidant activity of avian semen. Comp. Biochem. Physiol. B 120:527-533.
Surai, P.F., I. Kostjuk, G. Wishart, A. MacPherson, B. Speake, R. Noble, I. Ionov and E. Kutz. 1998b. Effect of vitamin E and selenium supplementation of cockerel diets on glutathione peroxidase activity and lipid peroxide susceptibility in sperm, testes and liver. Biol. Trace. Elem. Res. 64:119-132.
Swanson, C.A., B.H. Patterson, O.A. Levander, C. Veillon, P.R. Taylor, K. Helzlsouer, P.A. McAdam and L.A. Zech. 1991. Human [74Se]selenomethionine metabolism: kinetic model. Am. J. Clin. Nutr. 54:917-926.
Takahashi, K., N. Avissar, J. Whitin and H. Cohen. 1987. Purification and characterization of human plasma glutathione peroxidase: a selenoglycoprotein distinct from the known cellular enzyme. Arch. Biochem. Biophys. 256:677-686.
Tamura, T. and T.C. Stadtman. 1996. A new selenoprotein from human lung adenocarcinoma cells: purification, properties and thioredoxin activity. Proc. Natl. Acad. Sci. USA 93:1006-1011.
Terada, A., M. Yoshida, Y. Seko, T. Kobayashi, K. Yoshida, M. Nakada, K. Nakada, H. Echizen, H. Ogata and T. Rikihisa. 1999. Active oxygen species generation and cellular damage by additives of parenteral preparations: Selenium and sulfhydryl compounds. Nutrition 15:651-655.
Ursini, F, M. Maiorino and C. Gregolin. 1985. The selenoenzyme phospholipid hydroperoxides glutathione peroxidase. Biochim. Biophys. Acta 839:62-70.
van der Geyten, S, J.P. Sanders, E. Kaptein, V.M. Darras, E.R. Kuhn, J.L. Leonard and T.J. Tisser. 1997. Expression of chicken hepatic type I and type III iodothyronine deiodinases during embryonic development. Endocrinol. 138:5144-5152.
Vendland, S.C., M.A. Beilstein, J.Y. Yeh, C.L. Chen, O.N. Jensen, E Barofsky and PD. Whanger. 1993. Purification and properties of selenoprotein-W from rat muscle. J. Biol. Chem. 268:17103-17107.
Vendland, S.C., M.A. Beilstein, J.Y. Yeh, W. Ream and P.D. Whanger. 1995. Rat skeletal muscle selenoprotein W: cDNA clone and mRNA modulation by dietary selenium. Proc. Natl. Acad. Sci. USA 92:8749-8753.
Vinson, J.A. and P. Bose. 1987. Relative bioavailability of inorganic and natural selenium. In: Selenium in Biology and Medicine (G.F. Combs, Jr, O.A. Levander, J.E. Spallholz and J.E. Oldfield, eds). Van Nostrand, Inc., New York, New York, pp. 445-451.
Watabe, S., Y. Makino, K. Ogawa, T. Hiroi, Y. Yamamoto and S.Y. Takahashi. 1999. Mitochondrial thioredoxin reductase in bovine adrenal cortex. Its purification, nucleotide/amino acid sequence and identification of selenocysteine. Eur. J. Biochem. 264:74-84.
Wen, H.Y., R.L. Davis, B. Shi, J-J., Chen, L. Chen, M. Boylan and J.E. Spallholz. 1997. Bioavailability of selenium from veal, chicken, beef, pork , lamb, flounder, tuna, selenomethionine and sodium selenite assessed in selenium deficient rats. Biol. Trace Elem. Res. 58:43-53.
Whitacre, M. and J.D. Latshaw. 1982. Selenium utilization from menhaden fish meal as affected by processing. Poult. Sci. 61:2520-2522.
Williams, C. H., Jr. 1992. Lipoamide dehydrogenase, glutathione peroxidase, thioredoxin reductase and mercuric ion reductase - A family of flavoenzyme transhydrogenases. In: Chemistry and Biochemistry of Flavoenzymes. Vol. 3 (F. Muller, ed). CRC Press, Boca Raton, FL, pp. 121-211.
Wu, Q., K. Huang and H. Xu. 2003. Effects of longterm selenium deficiency on glutathione peroxidase and thioredoxin reductase expressions in rat aorta. J. Inorgan. Biochem. 94:301-306.
Xia, Y., L. Zhao, L. Zhu and P.D. Whanger. 1992. Metabolism of selenate and selenomethionine by a selenium-deficient population of men in China. J. Nutr. Biochem. 3:202-210.
Authors: FRANK W. EDENS and KYMBERLY M. GOWDY
Department of Poultry Science, North Carolina State University, Raleigh, North Carolina, USA


United States





