Effect of Dietary Antimicrobials on Immune Status in Broiler Chickens
Author details:
Animal Parasitic Diseases Laboratory, Animal and Natural Resources Institute, Agricultural Research Service, USDA, Beltsville, MD 20705, USA; 1 Lasher Poultry Diagnostic Laboratory, University of Delaware, Georgetown, DE 16483, USA; 2 Mountaire Farms Inc., Millsboro, DE 19966, USA; 3 Department of Animal Science and Technology, Chung-Ang University, Anseong 456-756, Korea; 4 Danisco, W227 N752 Westmound Drive, Waukesha, WI 53186, USA; 5 Department of Pediatrics, University of Maryland School of Medicine, Baltimore, MD 21201, USA.
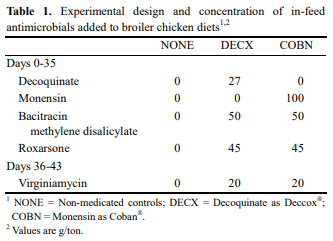
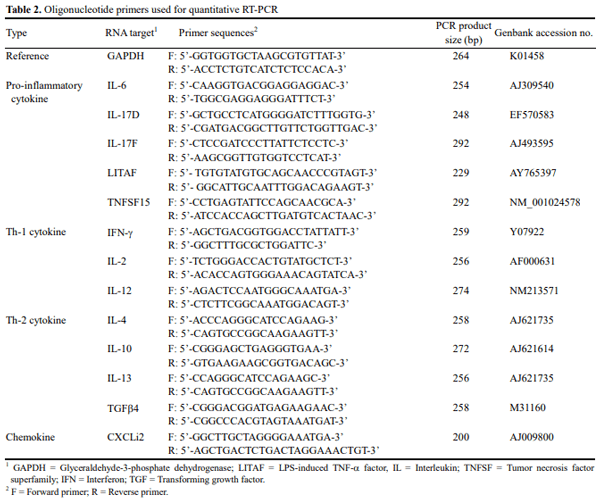
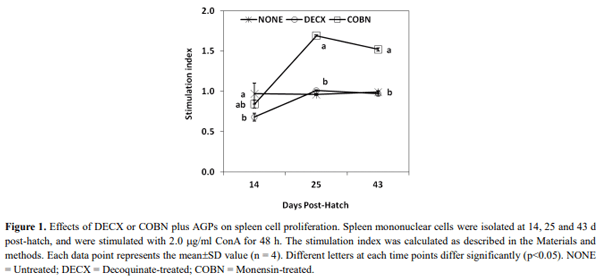
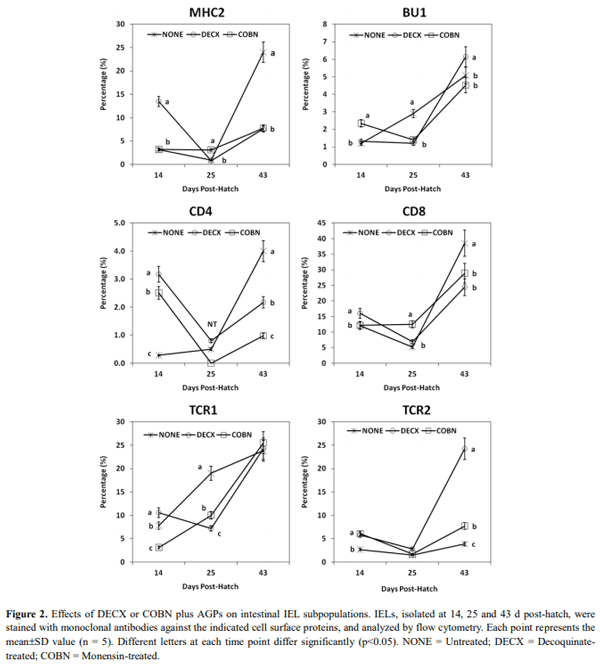
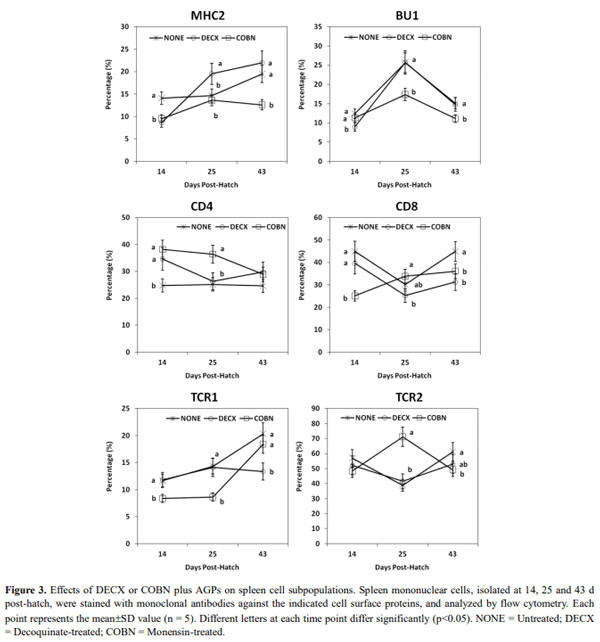
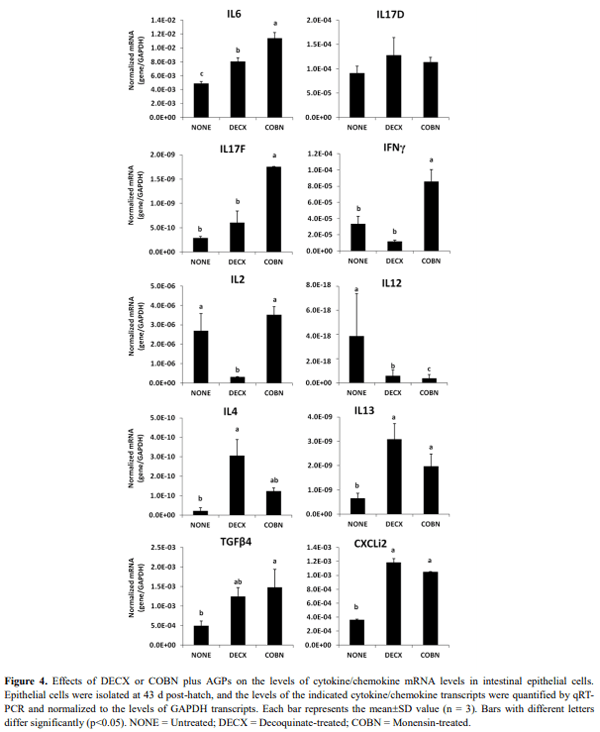
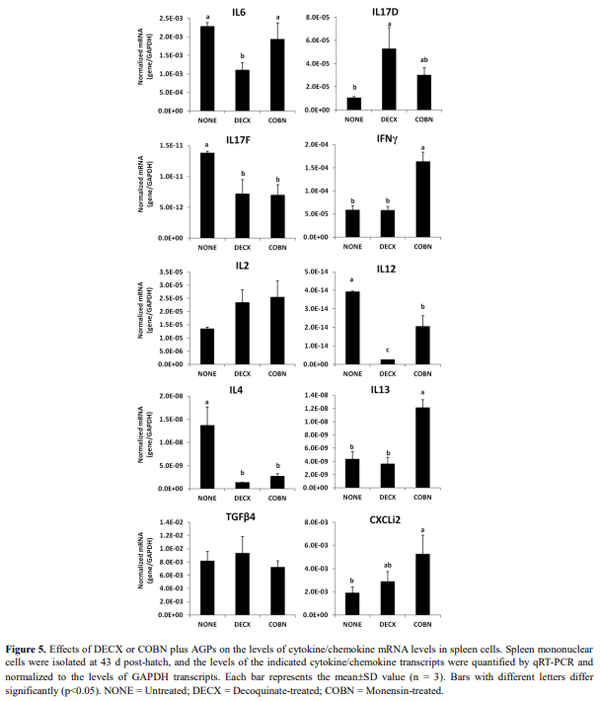
This study evaluated the effects of dietary anticoccidial drugs plus antibiotic growth promoters (AGPs) on parameters of immunity in commercial broiler chickens. Day-old chicks were raised on used litter from a farm with endemic gangrenous dermatitis to simulate natural pathogen exposure and provided with diets containing decoquinate (DECX) or monensin (COBN) as anticoccidials plus bacitracin methylene disalicylate and roxarsone as AGPs. As a negative control, the chickens were fed with a non-supplemented diet. Immune parameters examined were concanavalin A (ConA)-stimulated spleen cell proliferation, intestine intraepithelial lymphocyte (IEL) and spleen cell subpopulations, and cytokine/chemokine mRNA levels in IELs and spleen cells. ConA-induced proliferation was decreased at 14 d post-hatch in DECX-treated chickens, and increased at 25 and 43 d in COBN-treated animals, compared with untreated controls. In DECX-treated birds, increased percentages of MHC2+ and CD4+ IELS were detected at 14 d, but decreased percentages of these cells were seen at 43 d, compared with untreated controls, while increased TCR2+ IELs were evident at the latter time. Dietary COBN was associated with decreased fractions of MHC2+ and CD4+ IELs and reduced percentages of MHC2+, BU1+, and TCR1+ spleen cells compared with controls. The levels of transcripts for interleukin-4 (IL-4), IL-6, IL-17F, IL-13, CXCLi2, interferon-γ (IFN-γ), and transforming growth factorβ4 were elevated in IELs, and those for IL-13, IL-17D, CXCLi2, and IFN-γ were increased in spleen cells, of DECX- and/or COBN-treated chickens compared with untreated controls. By contrast, IL-2 and IL-12 mRNAs in IELs, and IL-4, IL-12, and IL-17F transcripts in spleen cells, were decreased in DECX- and/or COBN-treated chickens compared with controls. These results suggest that DECX or COBN, in combination with bacitracin and roxarsone, modulate the development of the chicken post-hatch immune system.
(Key Words : Decoquinate, Monensin, Bacitracin, Roxarsone, Immunity, Broiler)
Baba, T., N. Yamashita, H. Kodama, A. M. Mucamoto, M. Asada, K. Nakamoto, Y. Nose and E. D. Mcgruder. 1998. Effect of tylosin tartrate (Tylan soluble? ) on cellular immune responses in chickens. Poult. Sci. 77:1306-1311.
Bach Knudsen, K. E. 2001. Development of antibiotic resistance and options to replace antimicrobials in animal diets. Proc. Nutr. Soc. 60:291-299.
Chapman, H. D., T. K. Jeffers and R. B. Williams. 2010. Forty years of monensin for the control of coccidiosis in poultry. Poult. Sci. 89:1788-1801.
Chen, C. H., L. L. Ager, G. L. Gartland and M. D. Cooper. 1986. Identification of a T3/T-cell receptor complex in chickens. J. Exp. Med. 164:375-380.
Cheung, K. S., H. S. Lillehoj and M. C. Jenkins. 1991. Avian leucocyte common antigens: molecular weight determination and flow cytometric analysis using new monoclonal antibodies. Vet. Immunol. Immunopathol. 28:259-273.
Chichlowsk, M., J. Croom, B. W. McBride, L. Daniel, G. Davis and M. D. Koci. 2007. Direct-fed microbial PrimaLac and salinomycin modulate whole-body and intestinal oxygen consumption and intestinal mucosal cytokine production in the broiler chicks. Poult. Sci. 86:1100-1106.
Cressman, M. D., Z. Yu, M. C. Nelson, S. J. Moeller, M. S. Lilburn and H. N. Zerby. 2010. Interrelations between the microbiotas in the litter and in the intestines of commercial broiler chickens. Appl. Environ. Microbiol. 76:6572-6582.
Dibner, J. J., J. D. Richards and C. D. Knight. 2008. Microbial imprinting in gut development and health. J. Appl. Poult. Res. 17:174-188.
EFSA. 2003. Opinion of the scientific panel on additives and products or substances used in animal feed on a request from the commission on the coccidiostat DECCOX in accordance with article 9G of council directive 70/524/EEC. EFSA J. 17:1-40.
Erf, G. F., W. G. Bottje and T. K. Bersi. 1998. CD4, CD8 and TCR defined T-cell subsets in thymus and spleen of 2- and 7-week old commercial broiler chickens. Vet. Immunol. Immunopathol. 62:339-348.
Hong, Y. H., H. S. Lillehoj, E. P. Lillehoj and S. H. Lee. 2006. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following Eimeria maxima infection of chickens. Vet. Immunol. Immunopathol. 114:259- 272.
Larson, H. E. and S. P. Borriello. 1990. Quantitative study of antibiotic-induced susceptibility to Clostridium difficile enterocecitis in hamsters. Antimicrob. Agents Chemother. 34:1348-1353.
Lee, K. W., S. H. Lee, H. S. Lillehoj, G. X. Li, S. I. Jang, U. S. Babu, M. S. Park, D. K. Kim, E. P. Lillehoj, A. P. Neumann, T. G. Rehberger and G. R. Siragusa. 2010. Effects of direct-fed microbials on growth performance, gut morphometry, and immune characteristics in broiler chickens. Poult. Sci. 89:203- 216.
Lee, K.W., H. S. Lillehoj, S. H. Lee, S. I. Jang, D. Ritter, D. A. Bautista and E. P. Lillehoj. 2011a. Impact of fresh or used litter on the post-hatch immune system of commercial broilers. Avian Dis. (in press).
Lee, K. W. and H. S. Lillehoj. 2011. Antimicrobials, gut microbiota and immunity in chickens. Korean J. Poult. Sci. 38:155-164.
Lee, S. H., H. S. Lillehoj, S. I. Jang, K. W. Lee, D. Bravo and E. P. Lillehoj. 2011b. Effects of dietary supplementation with phytonutrients on vaccine-stimulated immunity against infection with Eimeria tenella. Vet. Parasitol. 181:97-105.
Lee, Y. K. and S. K. Mazmanian. 2010. Has the microbiota played a critical role in the evolution of the adaptive immune system? Science 330:1768-1773.
Lillehoj, H. S. and K. S. Chung. 1992. Postnatal development of Tlymphocyte subpopulations in the intestinal intraepithelium and lamina propria in chickens. Vet. Immunol. Immunopathol. 31:347-360.
Lillehoj, H. S., E. P. Lillehoj, D. Weinstock and K. A. Schat. 1988a. Functional and biochemical characterizations of avian T lymphocyte antigens identified by monoclonal antibodies. Eur. J. Immunol. 18:2059-2065.
Lillehoj, H. S. 1998. Role of T lymphocytes and cytokines in coccidiosis. Int. J. Parasitol. 28:1071-1081. Lillehoj, H. S. and E. P. Lillehoj. 2000. Avian coccidiosis. A review of acquired intestinal immunity and vaccination strategies. Avian Dis. 44:408-425.
Lillehoj, H. S., S. Kim, E. P. Lillehoj and L. D. Bacon. 1988b. Quantitative differences in Ia antigen expression in the spleens of 15I5-B congenic and inbred chickens as defined by a new monoclonal antibody. Poult. Sci. 67:1525-1535.
Lu, J., C. Hofacre, F. Smith and M. D. Lee. 2008. Effects of feed additives on the development on the ileal bacterial community of the broiler chicken. Animal 2:669-676.
Martel, A., L. A. Devriese, K. Cauwerts, K. De Gussem, A. Decostere and F. Haesebrouck. 2004. Susceptibility of Clostridium perfringens strains from broiler chickens to antibiotics and anticoccidials. Avian Pathol. 33:3-7.
Morikawa, K., H. Watabe, M. Araake and S. Morikawa. 1996. Modulatory effect of antibiotics on cytokine production by human monocytes in vitro. Antimicrob. Agents Chemother. 40:1366-1370.
Muller, P. Y., H. Janovjak, A. R. Miserez and Z. Dobbie. 2002. Processing of gene expression data generated by quantitative real-time RT-PCR. Biotechniques 32:1372-1379.
Munir, K., M. A. Muneer, A. Tiwari, E. Masaoud and R. M. Chaudhry. 2009. Effects of salinomycin on cell-mediated immunity of broiler chickens against hydropericardium syndrome and Newcastle disease viruses. Poult. Sci. 88:86-91.
Neumann, A. P., J. A. Benson, K. W. Lee, G. D. Ritter, D. A. Bautista, H. S. Lillehoj and G. R. Siragusa. 2011. Molecular analysis of broiler chicken gastrointestinal microbial communities associated with common anticoccidial feeding programs. In: Proceedings of International Poultry Scientific Forum, Atlanta, GA. p. 20.
Park, S. S., H. S. Lillehoj, P. C. Allen, D. W. Park, S. Fitzcoy, D. A. Bautista and E. P. Lillehoj. 2008. Immunopathology and cytokine responses in broiler chickens coinfected with Eimeria maxima and Clostridium perfringens with the use of an animal model of necrotic enteritis. Avian Dis. 52:14-22.
Parry, S. H., W. D. Allen and P. Porter. 1977. Intestinal immune response to E. coli antigens in the germ-free chickens. Immunology 32:731-741.
Reato, G., A. M. Cuffini, V. Tullio, N. Mandras, J. Roana, G. Banche, R. Foa and N. A. Carlone. 2004. Immunomodulating effect of antimicrobial agents on cytokine production by human polymorphonuclear neutrophils. Int. J. Antimicrob. Agents 23:150-154.
Ritter, G., G. Siragusa, S. Dunham and A. Neumann. 2010. Role of antibiotics on gut microbiota and incidence of gangrenous dermatitis in commercial broilers. Poult. Sci. 89(Suppl. 1): 480(Abstr.).
Rothwell, L., J. R. Young, R. Zoorob, C. A. Whittaker, P. Hesketh, A. Archer, A. L. Smith and P. Kaiser. 2004. Cloning and characterization of chicken IL-10 and its role in the immune response to Eimeria maxima. J. Immunol. 173:2675-2682.
Round, J. L. and S. K. Mazmanian. 2009. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 9:313-323.
Sekirov, I., N. M. Tam, M. Jogova, M. L. Robertson, Y. Li, C. Lupp and B. B. Finlay. 2008. Antibiotic-induced perturbations of the intestinal microbiota alter host susceptibility to enteric infection. Infect. Immun. 76:4726-4736.
Takahashi, K., Y. Miura and T. Mizumo. 2011. Antibiotics feeding accelerate functional maturation of intestinal immune-related cells of male broiler chicks after hatch. J. Poult. Sci. 48:187- 193.
Yin, Y., F. Lei, L. Zhu, S. Li, Z. Wu, R. Zhang, G. F. Gao, B. Zhu and X. Wang. 2010. Exposure of different bacterial inocula to newborn chicken affects gut microbiota development and ileum gene expression. ISME J. 4:367-376.
Yin, Y.-L., Z. R. Tang, Z. H. Sun, Z. Q. Liu, T. J. Li, R. L. Huang, Z. Ruan, Z. Y. Deng, B. Gao, L. X. Chen, G. Y. Wu and S. W. Kim. 2008. Effect of galacto-mannan-oligosaccharides or chitosan supplementation on cytoimmunity and humoral immunity in early-weaned piglets. Asian-Aust. J. Anim. Sci. 21:723-731.











