Assessing Dietary Branched-Chain Amino Acids to Achieve Linear Programming Goals through Model Extrapolation and Empirical Research
Published: August 4, 2022
By: Craig W. Maynard 1, Ed E. Gbur 2, Vinh-Loi Ly 3, Minh-Duc Le 3, Thi H. Ngan Le Jr. 3, Justina Caldas 4, and Michael T. Kidd 1 / 1 Center of Excellence for Poultry Science, Division of Agriculture, University of Arkansas, Fayetteville, AR, USA; 2 Agricultural Statistics Laboratory, Division of Agriculture, University of Arkansas, Fayetteville,
AR, USA; 3 Computer Science and Computer Engineering, University of Arkansas, Fayetteville, AR, USA; 4 Aviagen, Inc., Huntsville, AL, USA.
Overview
Renewed interest, especially in the United States, has sparked in assessing branched-chain amino acid interactions in practical diets for broilers. Indeed, as L-valine enters formulation bird nitrogen excesses are reduced as diet protein falls to the new first limiting amino acid (e.g., isoleucine, arginine, or tryptophan). For a United States based example, the result is less oilseeds and more gains, which typically result in increased inclusions in corn or corn by-products, coupled with a concomitant increase in dietary leucine.
The proceedings outline the foundations of the branched-chain amino acid early research, antagonism studies, and a meta-analysis conducted on publications with Cobb and Ross birds from 2000 to present. Results indicate that branched-chain amino acid interactions can occur in broilers fed on practical diets, and that responses vary by strain.
Foundations
The history of branched-chain amino acid research in poultry started in the 1940’s with the determination that these amino acids were required in poultry to facilitate growth (Almquist and Grau, 1944; Hegsted, 1944). This classification lead to the work of Grau and Peterson (1946) who published the first set of branched-chain amino acid requirements, establishing requirements as 0.5, 1.5, and 0.7 percent of the diet for isoleucine, leucine, and valine, respectively. Two years later, Almquist (1948) wrote a review of amino acid requirements in preparation for the second revision of the National Research Council’s Nutrient Requirement of Poultry (1950) in which he indicated that the requirements were likely 0.6, 1.4, and 0.8 percent of the diet for isoleucine, leucine, and valine, respectively. Investigations into branched-chain amino acid requirements continued from these initial recommendations through the development of crystalline amino acid diets in the late 1950’s and 60’s (Klain et al., 1958; 1960; Dean and Scott, 1965) but these investigations were not of large concern for diet formulation beyond the laboratory as supplemental methionine and lysine were largely a new concept and nutritionists were hesitant in their use of supplemental amino acids (Kidd et al., 2013).
The race to characterize branched-chain amino acid antagonism
In the 1960’s, antagonism among the branched-chain amino acids were inadvertently discovered while researchers were testing the protein quality of various protein meals. Following an observation by Fisher et al. (1960) that additions of leucine to a sesame meal based diet resulted in depressed feed intake and body weight gain of chicks, Mathieu and Scott published an abstract in 1968 outlining what has been thought to be the first example of branched-chain amino acid antagonism in poultry. In this abstract, Mathieu and Scott (1968) reported that feeding diets containing excess leucine resulted in depressed body weight when isoleucine and valine were limiting. However, Laksevela had published a paper in 1960 outlining similar responses. The Leksevela (1960) paper implemented both an addition and deletion assay to test the quality of herring solubles, in which “combinative protein value” of herring solubles was reduced 27 percent when isoleucine was deleted from the diet, whereas an addition of leucine reduced the “combinative protein value” by 16 percent. While this discovery was not new to the world of nutrition, as branched-chain amino acid antagonism was known from rodent research by Harper et al. (1954), it kickstarted a series of investigations into the importance of this antagonism in poultry nutrition.
In the early 1970’s, D’Mello and Lewis published a series of four papers on amino acid interactions in chick nutrition, of these three contained studies focused on the branched-chain amino acids (D’Mello and Lewis 1970a;b; 1971). The first branched-chain amino acid paper, second in the amino acid series, sought to confirm the existence of this antagonism and determine if both isoleucine and valine were both independently affected by excess leucine.
D’Mello and Lewis (1970a) repeatedly showed over the course of five experiments that excess leucine depressed the growth of diets containing limiting or adequate levels of valine or isoleucine, but supplementation of the limiting branched-chain amino acid above requirement levels would restore growth. D’Mello and Lewis (1970a) indicated that these results confirmed the existence of branched-chain amino acid antagonism and that the antagonism between valine and leucine would likely be the most significance. In their second study, D’Mello and Lewis (1970b) investigated the effect of branched-chain amino acid antagonism on requirements for the branched-chain amino acids. Through the course of this set of experiments, D’Mello and Lewis (1970b) found that dietary isoleucine and valine requirements increased by approximately 0.121 and 0.218 percent for every percent increase in dietary leucine levels. On a percent of total diet basis, these changes corresponded to a 0.047 and 0.120 percentage unit increase for isoleucine and valine, respectively, when leucine was increased by 1.000 percentage units (Figure 1). These conclusions agreed with their previous paper in that the interaction between valine and leucine is the most significant due to the heightened effect that leucine has on the valine requirement. In the final paper of the series, D’Mello and Lewis (1971) sought to determine whether the depression of feed intake caused by excess leucine was responsible for the reduced body weight gain associated with the antagonism as feed intake was one of the key findings that lead to the identification of this antagonism. Chicks were pair fed a diet containing adequate and excessive levels of leucine, which resulted in a continued disparity of body weight between the two groups. Furthermore, as feed intake was identical between groups, feed conversion was decreased by 11 percent when chicks fed a basal diet compared with chicks fed a diet containing excess leucine, indicating a metabolic mechanism behind the antagonism.
Further emphasizing the idea that a mechanism existed behind the branched-chain amino acid antagonism, Allen and Baker (1972) conducted a series of six experiments to test the efficacy of isoleucine and valine to promote chick body weight gains when leucine was fed in excess. Dose response curves were constructed through the course of these experiments and compared to a control in order to perform a slope-ratio comparison. These techniques resulted in the finding that isoleucine’s efficacy reduced linearly from 100 to 80 percent as leucine was increased from 0 to 3 percent, whereas the reduction in valine efficacy followed a quadratic curve, decreasing from 100 to 74 percent as leucine was increased from 0 to 6 percent, with an inclusion rate of 5.57 percent at the valine efficacy minima. Evaluating the raw data at a common leucine inclusion level, 3 percent, isoleucine and valine efficacy were, on average, 81 and 79 percent respectively. This datapoint again shows that valine is more affected by excess leucine, although to a lesser extent than previous researchers had found.
As the transamination step of the branched-chain amino acids was found to utilize a common enzyme (Ichihara and Koyama, 1966) and Featherston and Horn (1973) indicated that a common decarboxylation enzyme was used on branched-chain amino acid α-keto acids, researchers investigated the theory that excess leucine resulted in increased catabolism of valine and isoleucine. A pair of studies conducted at the University of Nottingham investigating this hypothesis and found minimal differences in the catabolism of valine when excess leucine was fed (Boorman and Buttery, 1972; Boldizsár et al., 1973). Contrary to these findings, Smith and Austic (1978) observed a significant increase in valine and isoleucine catabolism, as measured by CO2 production from C14 labeled isoleucine and valine, when 2.25 percent leucine was fed. The findings also indicated that the increase in catabolism was the same for both valine and isoleucine as CO2 production increased by 16 percent for both amino acids. Four years later, Calvert et al. (1982) published a paper in which a significant increase in valine and isoleucine catabolism was observed when excess leucine was fed. A larger increase in leucine was employed, 5%, which resulted in a 50 and 43 percent increase in catabolism for isoleucine and valine, respectively. While these relative increases in catabolism pose a strong indicator as to the mechanism behind branched-chain amino acid antagonism, Calvert et al. (1982) also force-fed chicks control or diets containing excess leucine to determine the influence that reduced feed intake had on performance variables, similar to the work done by D’Mello and Lewis (1971) 11 years prior. In agreement with the previous work, Calvert et al. (1982) found that differences in growth persisted even with equal feed intake, but attributed approximately 70 percent of the depressions in growth to reduced feed intake.
In the late 1980’s the topic of branched-chain amino acids shifted back to understanding its impacts on performance and ultimately its practicality. While attempting to reduce crude protein in broiler diets, Mendonca and Jensen (1989) found that if valine and isoleucine were supplemented incorrectly (i.e., if valine was marginally adequate and isoleucine was added) performance could be negatively affected due to antagonism between the two without drastically increased levels of leucine. Farran and Thomas (1990) then attempted to determine the requirements of the branched-chain amino acids as a whole, the first time since D’Mello had determined requirements in 1974 using plasma levels. Due to the interconnected nature of the branched-chain amino acids, Farran and Thomas (1990) implemented a central-composite, rotatable design in which levels of all three were manipulated. At the conclusion of their study, Farran and Thomas (1990) reported that leucine did not influence the requirements of valine and isoleucine, therefore excluding leucine from their contour plots when determining the valine and isoleucine requirements.
Along the same vein, two papers were published attempting to determine the isoleucine requirement in practical type diets (Barbour and Latshaw, 1992; Burnham et al., 1992). Both papers determined that branched-chain amino acid antagonism was of little concern in practical diets, citing that classic studies modeling this antagonism implemented large amounts of feedgrade L-leucine creating large shifts in dietary leucine levels. These large shifts were found to be hard pressed to occur using conventional protein sources. The final paper evaluating branched-chain antagonism was published by Waldroup et al. in 2002. Waldroup et al. (2002) used corn gluten meal to drive leucine levels in practical type diets in order to observe how branched-chain amino acid antagonism could present in commercial settings. It was determined that this antagonism was of little concern in practical formulation as long as branched-chain amino acid requirement minimums were maintained according to the ideal protein concept. At the time the work of Waldroup et al. (2002) was published, diets contained feed grade lysine and methionine with growing acceptance of L-threonine (Kidd et al., 2013). These dietary conditions resulted in excesses of both valine and isoleucine as dietary crude protein level was held up by the threonine minimum. Waldroup et al. (2002) acknowledged this in the concluding remarks of the paper, indicating that branched-chain amino acid antagonism may play a larger role when dietary crude protein is reduced, thereby reducing inherent excesses of valine and isoleucine.
A renewed endeavor
With the turn of the 21st century, branched-chain amino acid antagonism was shifted aside in favor of the determination of requirements. From 2002-2020, 41 titration studies were conducted and published elucidating requirements of the branched-chain amino acids (Maynard et al., 2021a). While the bulk of these studies focused on valine requirements, isoleucine work continued at a similar rate. The similar progression of valine and isoleucine research likely stems from the close limitation of the two amino acids in common broiler diets (Kidd and Hackenhaar, 2005). Likewise, leucine research largely fell to the wayside in practical poultry research due to its common excess in poultry diets (Maynard et al., 2021b).
Leucine made a resurgence in amino acid requirement research in 2017 when Ospina Rojas et al. published the first of three papers on the interactive effects of leucine and valine on their respective requirements. For a 21 to 42 d feeding period, Ospina-Rojas et al. (2017) found an interaction between valine and leucine for feed intake and body weight gain. While the response for body weight gain resulted in a true maximum allowing for determination of requirement estimates (valine to lysine, 83; leucine to lysine, 111), the feed intake response had two maxima values along the edges of the response surface. The observations of two maxima for feed intake indicated that as leucine was increased, it would require a subsequent increase in dietary valine in order to attain maximum feed intake. In the third paper by Ospina-Rojas et al. (2019) an earlier feeding period, 1 to 21 d, was implemented in which responses were observed for both feed intake and feed conversion. Unlike in the older feed phase tested in their previous paper, both responses resulted in single maximal responses (feed intake: valine to lysine, 77; leucine to lysine 104; feed conversion: valine to lysine, 73; leucine to lysine, 102). The differences in responses between the two papers are intriguing but an interesting observation can be made when looking at the response surface for feed conversion in Ospina-Rojas et al. (2019). Contrary to the valine-leucine relationship observed in 21 to 42 d feed intake (Ospina-Rojas et al., 2017), the contour plot for 1 to 21 d feed conversion shows that better feed conversion was observed when valine was decreased as leucine increased. The second paper by Ospina Rojas et al. (2018) discussed tibiotarsus bone characteristics but did not discuss broiler performance.
In 2019, Zeitz et al. published a pair of papers evaluating the effect of increasing leucine alone (Zietz et al., 2019a) or the branched-chain amino acids as a group (Zeitz et al., 2019b). In these studies, Zietz et al. (2019a) found no influence of increasing leucine from approximately 129 to 183 leucine to lysine on live performance and carcass traits when isoleucine and valine levels were fixed. In the second study where all three branched-chain amino acids were increased, but maintained at a constant ratio, differences were observed for d 1 to 10 feed conversion, d 11 to 21 body weight gain, and d 21 and 35 breast meat yield. While the differences in these two responses pose another interesting quandary, two differences occurred between the trials. The strain of the birds used differed with Ross broilers used in the excess leucine study (Zeitz et al., 2019a) and Cobb broilers used in the maintained ratio study (Zeitz et al., 2019b). The second difference, and likely the more important, was that all base tested levels of leucine in the first study were above requirement values for leucine, ranging from 123 to 135 leucine to lysine (Zeitz et al., 2019a). In stark contrast to this, the second study contained values of 110, 95, and 101 leucine to lysine for the 1 to 10, 11 to 21 and 22 to 35 d feeding periods, respectively (Zeitz et al., 2019b). Although few studies on leucine requirements exist, the leucine requirement for growth and carcass traits is approximately 108 leucine to lysine (Maynard et al., 2021a). Therefore, the differences generated in Zeitz et al. (2019b) are likely driven by the deficient leucine levels fed in the low leucine diets as performance of these birds were the lowest among the three treatments. Interestingly, despite the potential leucine deficiency, birds fed the low leucine diets had higher breast meat yields at both 21 and 35 d than those fed the highest levels of leucine.
The work of Ospina-Rojas et al. continued with the publishing of a paper in 2020 that evaluated all three branched chain amino acids. This study combined the factorial research they had previously employed with modeling techniques through the implantation of central composite, rotatable design. Three central-composite, rotatable design studies were conducted in order to determine branched-chain amino acid requirements for 1 to 14, 14 to 28, and 28 to 42 d feeding periods. Significant interactions were observed for both body weight gain and feed conversion. Generated requirements from Ospina-Rojas et al. (2020) are presented in Table 1. For both parameters leucine was determined to have the most influence on the response based on eigenvector values. The second most influential branched-chain amino acid varied, but for feed conversion valine was found to be the second most influential branched-chain amino acid in the starter period but was replaced by isoleucine in subsequent periods. Interestingly, requirement estimates for both followed the same period where they decreased from the starter to grower and slightly increased in the finisher.
Modeling work continued with the branched-chain amino acids with the publication of two studies implementing Box-Behnken design (Kidd et al., 2021b; Maynard et al., 2021b). In the first paper, Maynard et al. (2021b) conducted two studies using Cobb 500 male broilers in which the three test factors were valine, isoleucine, and leucine and valine, isoleucine, and glycine + serine. The valine, isoleucine, and leucine study yielded significant interactions between isoleucine and valine for body weight gain, feed conversion, and breast meat yield and between leucine and valine for breast meat yield. All of the response surface graphs generated from these interactions resulted in saddle points in which no requirement could be generated due to more than one “optimal” combination. Due to the minimal apparent involvement of leucine in the observed responses, leucine was replaced with glycine + serine in the second Box-Behnken design experiment. This replacement resulted in a lack of similar responses observed in the previous experiment. Due the lack of continuity, Maynard et al. (2021b) indicated that the effect of leucine may be a “shadow effect” in that it instigates the antagonism but cannot be directly pinned unless levels reach overly excessive levels. These findings align with those of Farran and Thomas (1990) 30 years prior, who similarly ruled out the influence of leucine in practical diets. The second study, conducted by Kidd et al. (2021b) evaluated the effects of valine, isoleucine, and leucine using the same methodology and design as Maynard et al. (2021b) but used male and female Lohman Indian River broilers, which represents a commercial Aviagen line sold in parts of Southeast Asia and mimics the Ross 308. Responses from this study indicated that, for the Lohman Indian River broilers, females were more responsive to manipulation of the branched-chain amino acids with interactions identified for body weight gain, feed conversion, carcass yield, and breast yield, whereas carcass yield was the only response for males. As with Maynard et al. (2021b), responses generated saddle points and prevented the determination of requirements for the majority of interactions, with the only exception being for female carcass yield, producing a requirement around 75 and 130 for valine and leucine to lysine, respectively. Although different sex broilers were used to generate responses, comparison across the Maynard et al. (2021b) and Kidd et al. (2021b) papers elucidate the difference in response to branched-chain amino acid supplementation between the Cobb 500 and Ross 308 type “Lohman Indian River” broilers (Figure 2).
Comparing strain responses via meta-analysis
In an effort to assess recent data (2000 to present) for Cobb and Ross birds used in published studies aimed at evaluating the birds’ response to dietary branched-chain amino acids, a meta-analysis was conducted. The meta-analysis built on previous data that focused on Cobb broilers (Kidd et al., 2021a). Publications measuring Ross birds were included, and analysis of covariance was used to analyze the data. Each of the four responses (i.e., average daily body weight gain, average daily feed intake, feed conversion ratio, and breast meat yield) were assumed to follow a gamma distribution. The initial model contained the three amino acids (i.e., valine, isoleucine, and leucine) as the covariates and strain as the factor. For each of the four responses, the initial model allowed all of the model coefficients to depend on strain. The amino acid term with the largest p-value was removed from the model and the reduced model was refitted. The process was repeated until all remaining terms were significant at the five percent level. The least square means for selected levels of the amino acids in the final model were compared using a protected least significant difference (LSD) procedure. All analyses were carried using the SAS procedure GLIMMIX (version 9.4).
The four responses in the meta-analysis consisted of average daily body weight gain, average daily feed intake, feed conversion ratio, and breast meat yield. Least square means of valine, isoleucine, and leucine levels selected for final model assessment were: 0.70, 0.74, 0.78, 0.82, and 0.86; 0.62, 0.65, 0.68, 0.71, and 0.74; and 1.05, 1.35, 1.45, and 1.60, respectively. Analysis of PROC GLIMMIX produced several strain intercept and coefficient significant (P ≤ 0.05) responses. Specifically, Figures 3 and 4 represent strain differences to dietary valine, isoleucine, and leucine for feed intake and breast meat yield, respectively.
In addition to evaluation of the responses in SAS, a meta-regression using weighted least squares, as opposed to ordinary least squares, was used to create best fit non-linear functions. To address overfitting, both Akaike and Bayesian information criterion were used minimize predictors.
In these proceedings, feed intake data has been discussed indicating that branched-chain amino acid variations produce similar feed intake responses; however, differences in efficiency occur metabolically as feed conversion ratios vary. Moreover, the meta-analysis indicates Cobb and Ross strains responded similarly to feed intake from increases in dietary leucine to lysine ratios from 1.10 to 1.25, but opposite feed intake responses as leucine to lysine ratios increased from 1.45 to 1.60 (Figure 3).
As the leucine to lysine ratio increased from 1.05 to 1.35 in Cobb broilers, breast meat yield increased, and this response was further improved with increasing isoleucine relative to lysine (Figure 4A). However, Ross broilers exhibited an opposite response with lowered breast meat yield to both increasing leucine and isoleucine relative to lysine (Figure 4A) when fed dietary leucine to lysine ratios at 1.10 or 1.25. However, as leucine to lysine was increased to 1.45 or 1.60, both strains responded linearly to isoleucine increases, as well as valine decreases, relative to lysine (Figure 4B).
Average daily gain of Ross broilers from the meta-regression is presented in Figure 5. Valine to lysine and isoleucine to lysine are presented on the y and z axes, respectively, with the left graph representing low leucine/lysine (< 1.40) and the right graph representing high leucine/lysine (> 1.40) experiments. Ross birds responded to increasing valine when fed lower levels of leucine. However, as leucine was increased, the highest weight gain was achieved when both valine and isoleucine were above typical ratio requirements (e.g., 0.77 and 0.68, respectively). Moreover, isoleucine had the greatest sensitivity to improving weight gain in the high leucine graph (Figure 5).
Conclusions
Overall, the current branched-chain amino acid research is reflective of the understanding and discoveries reported 30 years ago. While these current findings have not produced a definitive methodology or strategy as to how to deal with this antagonism, they have presented a new perspective on the conundrum. The repeated observation of saddle points gives a potential explanation for some of the conflicting information found in the literature. As the branched-chain amino acid “requirements” appear to be on a sliding scale dependent upon one another, the antagonism may be redefined as a synergism in the future when mitigation strategies may allow nutritionist to turn modern negative effects into positives through adequate supplementation of valine and isoleucine in order to handle inherent leucine excesses in the diet. The understanding of the branched-chain amino acids will likely require flexibility in the mind of researchers as the complexity of this interaction defies current research practices, in that it has been repeatedly displayed with p-values above the traditional 0.05 significance level but with a relative consistency below 0.10 across experiments and researchers. Moreover, further research on strain differences warrants attention.
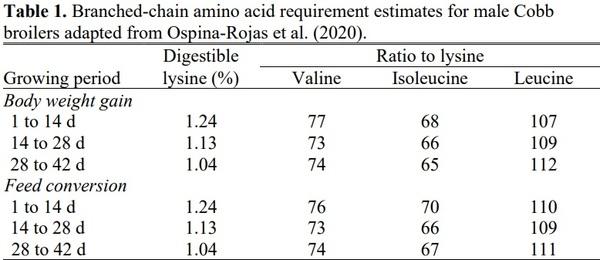
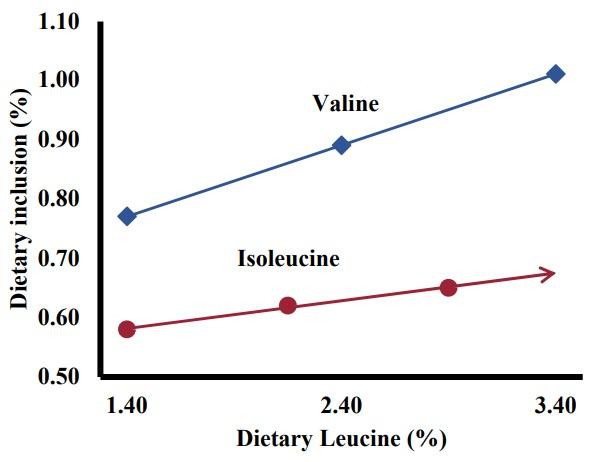
Figure 1. Increase in dietary valine and isoleucine requirement estimates when dietary leucine is increased. Adapted from D’Mello and Lewis (1970b).
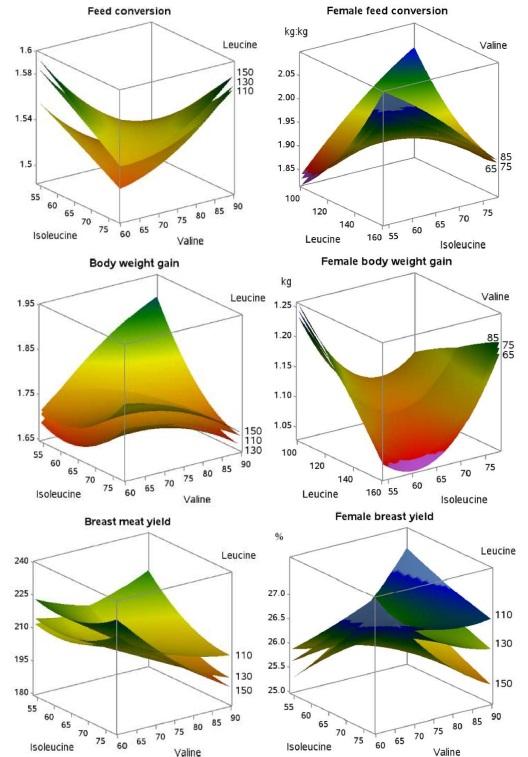
Figure 2. Influence of branched-chain amino acid supplementation on feed conversion, body weight gain, and breast meat yield of Cobb 500 male (left) and Lhoman Indian River female broilers (right).
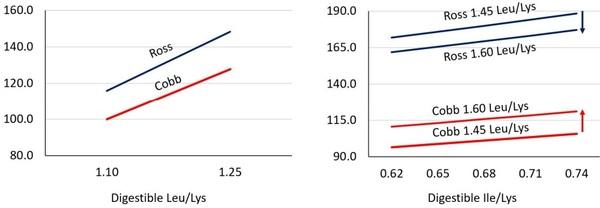
Figure 3. Feed intake of Cobb and Ross broilers from peer review manuscripts published from 2000 to present. Predicted least square means were used in meta-analysis data to assess strain trends at 1.10, 1.25 (Left), 1.45, and 1.60 (Right) digestible leucine to lysine ratio. Projections are to assess strain response differences to the branched-chain amino acids, and are not intended to suggest strain differences in feed intake. Estimates for low Leu levels (Left) are: Cobb intercept, Ross intercept, and Leu coefficient at P< 0.0001. Estimates for high Leu levels (Right) are: Cobb intercept, Ross intercept, Common Ile, Cobb Leu, and Ross Leu at P< 0.01.
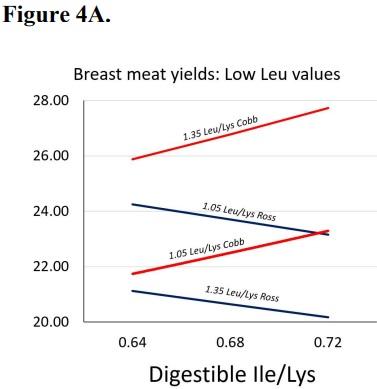
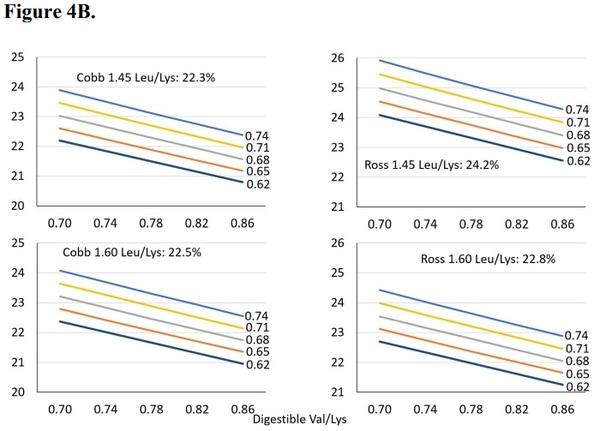
Figure 4. Breast meat yields of Cobb and Ross broilers from peer review manuscripts published from 2000 to present. Predicted least square means were used in meta-analysis data to assess strain trends at 1.10, 1.25 (Figure 4A), 1.45, and 1.60 (Figure 4B) digestible leucine to lysine ratio. Projections are to assess strain response differences to the branched-chain amino acids, and are not intended to suggest strain differences in yield. Estimates for low Leu levels (4A) are: Cobb intercept, Ross intercept, Ross Val coefficient, Cobb Ile coefficient, Ross Ile coefficient, Cobb Leu coefficient, and Ross Leu coefficient at P< 0.05. Estimates for high Leu levels (4B) are: Cobb intercept, Ross intercept, Common Val coefficient, Common Ile coefficient, and Ross Leu coefficient at P< 0.05.
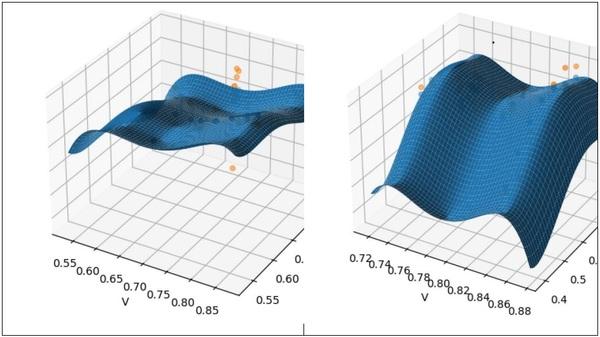
Figure 5. Average daily body weight gain (ADG) in 35-day old Ross broilers as affected by valine/lysine (I) and isoleucine/lysine (I) when leucine/lysine is below 1.40 (left) or above 1.40 (right).
Presented at the Arkansas Nutrition Conference, August 31st to September 2nd, 2021, Rogers, AR.
Content from the event:
Related topics:
Authors:

")

Aviagen

Influencers who recommended :
Ali AfsarRecommend
Comment
Share

Would you like to discuss another topic? Create a new post to engage with experts in the community.
Featured users in Animal Feed




")
