vitamin E for swine
Importance of form and source of vitamin E for swine
Vitamin E (α-tocopherol) is a critically important nutrient for all confinement-reared swine. It is a primary dietary antioxidant that maintains cell membrane integrity and also enhances humoral- and cell-mediated immunity. Other metabolic roles have been reviewed by Brigelius-Flohe and Traber (1999). Vitamin E is not synthesized in the body, thus all swine require dietary sources to meet metabolic needs. Serum alpha-tocopherol level is the method utilized to measure vitamin E adequacy in swine.
Production changes have dramatically increased the need for supplementing swine diets with vitamin E during all segments of swine production. The original NRC Nutrient Requirements of Swine was published in 1944 and subsequent revisions have been made since this first publication. The most recent revision was published in 1998 (NRC, 1998). The 1973 revision was the first to publish a requirement for vitamin E in swine (NRC, 1973). Prior to the 1973 revision, vitamin E supplementation was not recommended (see insert). 

Confinement feeding deprives swine from an excellent source of vitamin E from fresh forages. They become more dependent upon vitamin E supplementation compared to swine allowed to graze. During gestation and lactation, sows on pasture had higher vitamin E status compared to sows fed in confinement (Mutetikka and Mahan, 1993). The current vitamin E requirements for both gestation and lactation diets were increased from 11 IU per kg to 44 IU per kg diet (40,000 I.U. per ton) (NRC, 1998). Since gestating sows are limit-fed and lactating sows are not, the published requirement of 44 I.U./kg in gestating diets may be too low for the gestating sow. Vitamin A requirements in gestating diets are twice the level for lactation diets, thus it would appear that the same should be appropriate for vitamin E, especially for the gestating sow with limited feed intake.
Piglets are born with very low tissue levels of vitamin E due to low placental transfer (Mahan, et al., 2000, Lauridsen et al, 2002a). Therefore, the neonatal pig is immediately deficient in vitamin E at birth and totally dependent upon colostrum and milk to provide α-tocopherol. Colostrum and milk levels are dependent upon quantities of vitamin E consumed by the dam during gestation and subsequent lactation (Loundenslager et al, 1986; Mahan, 1991; Mahan et al. 2000). After weaning, vitamin E status typically decreases due to poor utilization of dietary supplemented vitamin E (Bonnette et al. 1990; Chung et al. 1992). The piglet’s most critically important times of need for vitamin E appear to be at birth and immediately after weaning. It is essential that the newborn piglet receive adequate vitamin E via the colostrum and milk to enhance immunity and boost the piglet’s resistance to infectious diseases. It is also important that the supplemental vitamin E be in a form and source that the newly-weaned piglet can utilize (Specht et al. 2003).
Research has demonstrated that supplemental vitamin E during gestation and lactation improves sow litter size, increases sow’s colostrum and milk α-tocopherol and enhances health status (Malm et al, 1976; Mahan, 1991; Mahan, 1994; Wuryastuti et al, 1993). Recent research has demonstrated that supplementation of sows during late gestation, especially just prior to farrowing, increased the vitamin E content of colostrum and subsequently a very significant increase in the plasma vitamin E concentration of newborn piglets after colostrum consumption (Lauridsen et al, 2002a). After initial suckling, piglet plasma and tissues showed dramatic increases in vitamin E concentrations demonstrating the importance of colostrum and milk for enhancing the vitamin E status of newborn piglets.
Intestinal Absorption of Vitamin E
Vitamin E (alpha-tocopherol) is absorbed in the intestine and transported to the liver in chylomicrons via the lymph system. The two sources of vitamin E activity at the intestinal level are naturally occurring vitamin E from the diet (RRR-alpha-tocopherol) and supplemental vitamin E (typically vitamin E esters). Due to oxidation of natural vitamin E in feeds during processing and storage and interactions with other nutrients, rations are supplemented with vitamin E esters to meet the total dietary requirements since esters are more stable than un-esterified natural vitamin E. Prior to absorption, supplemental vitamin E esters must be de-esterified and both natural and supplemental sources have to be micellized prior to absorption and transport in the lymph system (Figure 1). 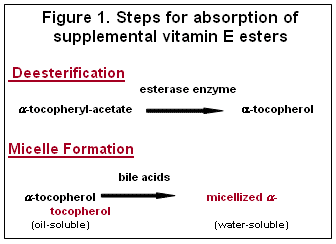
The efficiency of absorption of supplemental vitamin E esters can be very low and depends upon two main factors: adequate intestinal esterase to cleave the acetate moiety in the intestinal tract, and adequate bile salts to micellize alpha-tocopherol (fat-soluble) prior to absorption making it water-soluble which enhances absorption.
It has been shown that feed-supplemented α-tocopheryl-acetate is absorbed very poorly during early post-weaning, making it very difficult to increase and/or maintain vitamin E status of weanling pigs (Chung et al. 1992). Hedemann and Jensen 2001 proposed that the reason for poor absorption is a deficiency of a non-specific lipolytic pancreatic enzyme (carboxylester hydrolase, CEH) that cleaves ester linkages and is responsible for hydrolyzing vitamin E esters. A number of pancreatic enzymes, including CEH, decline upon weaning (Jensen et al, 1997; Hedemann and Jensen 2001). Low plasma vitamin E during the post-weaning period may be explained by a limited ability to hydrolyze vitamin E-esters and subsequently reduce efficiency of absorption. When newly-weaned piglets were fed synthetic vitamin E-acetate at levels up to 200 I.U./kg (181,000 I.U./ton) for four weeks post-weaning, none of the supplemental levels were able to maintain serum alpha-tocopherol levels when piglets were weaned (Moreira and Mahan, 2002). Dietary supplemental levels above 100 I.U./kg did not improve vitamin E status (Figure 2).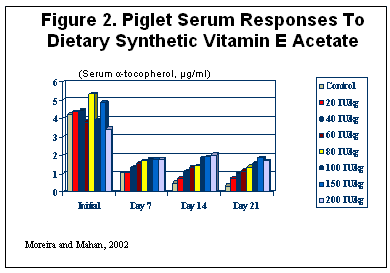
In order to enhance vitamin E absorption in sows and newly-weaned piglets, it is important to fortify the sow’s diet during late gestation and lactation, and supplement newly-weaned pigs with a source and form of vitamin E that the young piglet can utilize.
Role of Vitamin E in Immunocompetency
Vitamin E supplementation improves immune response to antigens by stimulating production of antibody-producing lymphocytes (Nockels, 1979; Tengerdy, et al. 1984).
In piglets, Rafaiet al. 1989 found thatvitamin E supplementation exerted a beneficial influence on the indices of cell-mediated immunity. The proportion of rosette-forming cells and blastogenic transformation induced by specific (horse globulin) and nonspecific (phytohaemagglutinin, PHA) mitogens showed the most significant increase in pigs supplemented with 5 mg selenium, 100 I.U. vitamin E and 5 mg riboflavin per kg diet. Jensen et al., 1988 reported that piglets with serum tocopherol levels above 3 μg/ml had optimum immune system function.
Babinszky et al. 1991 showed that immune response against ovalbumin was increased at 1 wk of age after immunization for weaned pigs from sows fed 136 I.U. vitamin E/kg diet. They also observed increased IgG concentration in pigs when dams were fed 48 or 136 I.U./kg vitamin E compared to NRC recommended levels. Mavromatis et al 1999 found that piglets born from sows supplemented with 30 mg alpha-tocopherol /kg diet plus 30 mg injectible Se (sodium selenite) on days 30, 60, and 90 of pregnancy, had the highest immunoglobulin concentration level up to weaning day compared to sows fed a basic feed containing 20 mg alpha-tocopherol (30 I.U.) per kg diet.
Since vitamin E is critically important for optimum function of the immune system, it is necessary to provide the vitamin at higher than typical levels fed to prevent frank deficiency symptoms (NRC, 1998). To achieve maximum immune response in pigs, serum α-tocopherol levels should be above 4 μg/ml while nursing and through the first two weeks post-weaning.
Sources of Vitamin E
Usually synthetic vitamin sources are, for the most part, equal in efficacy and structure to the natural source of that vitamin. That is not the case for vitamin E. Major differences in structure of natural-source and synthetic-source vitamin E exist. Natural vitamin E is a single entity, one isomer- RRR-α-tocopherol (or d-alpha-tocopherol). Synthetic vitamin E (all-rac-alpha-tocopherol or dl-alpha-tocopherol) consists of eight stereoisomers, with only one isomer identical to the natural RRR-isomer and the other seven isomers having different structures and lower biological activities. Furthermore, the RRR-isomer is preferentially transported and incorporated in tissues and milk, while some of the synthetic isomers have no biological activity (Lauridsen et al, 2001).
In nature, most plants and oils naturally contain a mixture of tocopherols (alpha-, beta-, gamma- and delta-) that show antioxidant activities. Of the four tocopherols, alpha-tocopherol has the highest biological potency and is the only tocopherol to be recognized to have vitamin E activity (DRI for Humans, 2000; Traber, 1999).
The two most common feed sources of supplemental vitamin E are natural-source (RRR-alpha-tocopherol) and synthetic-source (all-rac-alpha-tocopherol) and their corresponding acetate-esters. The International Unit (I.U.) converts quantities of vitamin E products to biological activity (United States Pharmacopeia, USP, 1999). The I.U. values for the various vitamin E products are given in Table I. The Dietary Reference Intake (DRI, 2000) has published a conversion for converting I.U. vitamin E activity to mg vitamin E activity. As can be seen in Table 1, the synthetic vitamin E sources are 67% less potent than an equal I.U. value of natural source vitamin E.
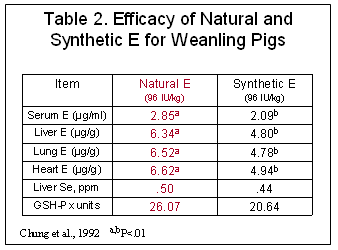
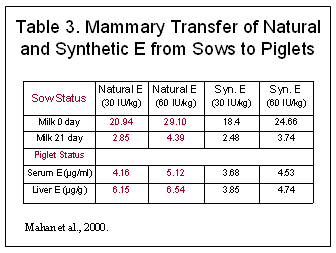
The biological assay utilized to arrive at the I.U. values was the rat fetal resorption assay; however recent research in humans and animals has shown that natural sources have at least twice the biological activity of an equal amount of synthetic (Traber et al, 1992), not 36% as derived using the rat fetal resorption assay. The rat fetal resorption appears to overestimate values of the synthetic sources compared to the natural sources. Research has shown preferential uptake and tissue deposition of the natural RRR-isomer compared to synthetic isomers in humans (Ingold, et al, 1987; Stone et al. 2003), pigs (Chung et al. 1992; Lauridsen et al. 2002b), and sows (Mahan et al. 2000, Lauridsen et al. 2002a; Lauridsen and Jensen, 2003) (Tables 2 and 3). These results demonstrate that the rat fetal resorption underestimates the value of natural vitamin E, especially in milk transfer which is critically important. 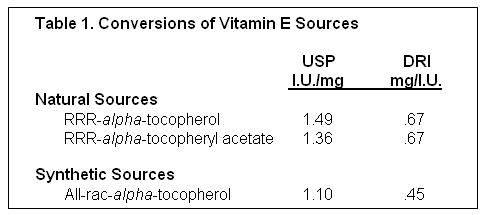
Importance of Form of Vitamin E
Since natural and synthetic vitamins E are antioxidants, esters are added to insure stability. However, the ester has to be removed and the alpha-tocopherol made water-soluble by the action of bile salts (Gallo-Torres, 1980) prior to absorption. This appears to be of greatest importance in post-weaned piglets immediately after weaning.
Research has shown that water-soluble, micellized, non-esterifed RRR-alpha-tocopherol is better utilized by newly-weaned piglets than other sources (Specht et al. 2003). However since the free tocopherol is unstable in complete feeds, the only viable means to provide micellized alpha-tocopherol is by water fortification. The only product commercially available that provides non-esterified RRR-alpha-tocopherol is EMCELLE® TOCOPHEROL (Stuart Products, Inc).
EMCELLE® TOCOPHEROL is a water-dispersible micellized source of d-alpha-tocopherol. Each ml contains 500 I.U. vitamin E activity. The product readily mixes into drinking water and does not affect water consumption. It should not be mixed into complete feeds and stored for extended time due to its unstable form. The micellization process converts the vitamin E oil into tiny microscopic water-soluble droplets (optimally 0.1 to 0.4 microns). This dramatically enhances the ability of pigs to absorb vitamin E (Specht et al, 2003, Figure 3). Being water soluble greatly enhances absorption from the small intestine and rapidly increases blood and tissue tocopherol levels. The micellized, non-esterified form of vitamin E, does not require the acetate-ester to be removed prior to absorption and there is no need for micellization by bile salts.
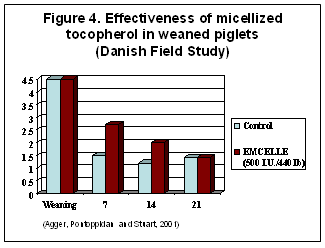
Micellized natural vitamin E has been shown to boost the vitamin E status and enhance survivability of piglets after weaning. Micellized natural vitamin E alcohol supplemented to post-weaned piglets (500 IU/200 kg BW in drinking water) for 10 days, increased plasma alpha-tocopherol at 7 and 14 days post-weaning in a Danish study (Agger et al. 2001, Figure 4). 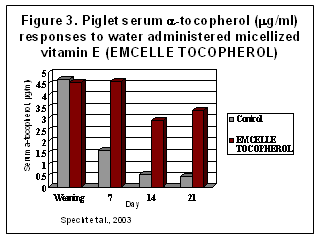
Summary
Confinement rearing increases the need for vitamin E supplementation. Generally, higher levels of supplementation enhance vitamin E status. Sows maintained in confinement and fed processed feedstuffs during gestation and lactation require at least 44 I.U. vitamin E per kg diet, and probably higher during gestation, due to limited feed intake.
Due to poor placental transfer and low body lipid stores, pigs are born with very low vitamin E status. Optimal supplementation of the sow’s diet during gestation and lactation is essential to promote increased levels of the vitamin in colostrum and milk in order to ensure adequate intake by nursing piglets. The benefit to piglets appears primarily to be enhanced immunity and possible reduction in cardiomyopathy.
Natural vitamin E is better utilized than synthetic by the sow, resulting in improved vitamin E status in the nursing pig through milk transfer of the natural RRR-isomer. Vitamin E status in piglets declines dramatically after weaning unless a biologically available source of vitamin E is provided. Research has shown that water supplementation of micellized natural vitamin E is the most effective method to enhance and maintain vitamin E status in post-weaned piglets.
Table 4 offers the vitamin E recommendations for all classes of swine. When using natural-source vitamin E in complete feeds, use approximately 70% of the synthetic vitamin E levels. For newly-weaned piglets, water supplementation of micellized tocopherol (EMCELLE TOCOPHEROL) is recommended over and above levels included in starter feed.
Literature Cited
Agger, N., L. Pontoppidan and R.L. Stuart. 2001. Increased immunity in weaned piglets by daily dosing of d-alpha-tocopherol in the drinking water. Acta Vet. Scand. Suppl. 98:286
Babinszky L., D.J. Langhout, M.W. Verstegen, L.A. den Hartog, P. Joling, and M. Nieuwland. 1991. Effect of vitamin E and fat source in sows' diets on immune response of suckling and weaned piglets.J Anim Sci. 69:1833.
Bonnette, E.D., E. T. Kornegay, M.D. Lindemann and C. Hammerberg. 1990. Humor and cell-mediated immune response and performance of weaned pigs fed four supplemental vitamin E levels and housed in two nursery temperatures. J. Anim. Sci. 68:1337-1345.
Brigelius-Flohe, R. and M.G. Traber. 1999. Vitamin E: function and metabolism. FASEB J. 13:1145.
Chung, Y.K, D.C. Mahan and A.J. Lepine. 1992. Efficacy of dietary d-α-tocopherol and dl-α-tocopheryl acetate for weanling pigs. J. Anim. Sci. 70: 2485.
DRI. 2000. Dietary reference intakes for vitamin C, vitamin E, selenium, and carotenoids. National Academy Press. Washington DC.
Ingold, K.U., A.C. Webb, D. Witter, G.W. Burton, T.A. Metcalfe and D.P. Muller. 1987. Vitamin E remains the major lipid-soluble, chain-breaking antioxidant in human plasma even in individuals suffering severe vitamin E deficiency. Arch. Biochem. Biophys. 259:224.
Jensen, S.K., C. Fossum, M. Ederoth and R.V.J. Hakkarainen. 1988. The effect of vitamin E on the cell-mediated immune response in pigs. J. Vet. Med. B. 35:549-555.
Hedemann, M.S. and S.K. Jensen. 2001. The activity of lipolytic enzymes is low around weaning - Measurements in pancreatic tissue and small intestinal contents. In: J.E. Lindberg and B. Ogle (eds) Proceedings of the 8th Symposium of Digestive Physiology of Pigs, Uppsala, Sweden. CABI Publishing, Wallingford Oxon, UK, 28-30.
Jensen, M.S., S.K. Jensen and K. Jakobsen. 1997. Development of digestive enzymes in pigs with emphasis on lipolytic activity in the stomach and pancreas. J. Anim. Sci. 75: 437.
Lauridsen, C. and S. K. Jensen. 2003. Dietary inclusion of vitamin E and C for sows and their progeny. Final Report, Dept of Animal Nutrition and Physiology. Danish Institute of Agr. Sci.
Lauridsen, C, H. Engel, A.M. Craig and M.G. Traber. 2002a. Relative bioactivity of dietary RRR- and all-rac-alpha-tocopheryl acetates in swine assessed with deuterium-labeled vitamin E. J. Anim. Sci. 80:702.
Lauridsen, C., H. Engel, S.K. Jensen, A.M. Craig and M.G. Traber. 2002b. Lactating sows and suckling piglets preferentially incorporate RRR- over all-rac-alpha-tocopherol into milk, plasma, and tissues. J. Nutr. 132:1258.
Loudenslager, M.J., P.K. Ku, P.A. Whetter, D.E. Ullrey, C.K. Whitechair, H.D. Stowe and E.R. Miller. 1986. Importance of diet of dam and colostrum to the biological antioxidant status and parenteral iron tolerance of the pig. J. Anim. Sci. 63:1905.
Mahan, D.C. 1991. Assessment of the influence of dietary vitamin E on sows and offspring in three parities: Reproductive performance, tissue tocopherol, and effects on progeny. J. Anim. Sci. 69:2904.
Mahan, D.C. 1993. Efficacy of dietary vitamin E on sow reproductive performance, tissue tocopherol and subsequent effects on their progeny over a three-parity period. Ohio State Univ. Ohio Swine Research and Industry Report 1992-1993.
Mahan, D. C. 1994. Effects of dietary vitamin E on sow reproductive performance over a five-parity period. J. Anim. Sci. 72:2870.
Mahan, D.C., Y.Y. Kim and R.L. Stuart. 2000. Effect of vitamin E sources (RRR- or all-rac-a-tocopheryl acetate) and levels on sow reproductive performance, serum, tissue, and milk a-tocopherol contents over a five-parity period, and the effects on the progeny. J. Anim. Sci. 78:110.
Malm, A.W., G. Pond, E.F. Walker Jr., M. Homan, A. Aydin and D. Kirtland. 1976. Effect of polyunsaturated fatty acids and vitamin E level of the sow gestation diet on reproductive performance and on level of alpha-tocopherol in colostrum, milk, and dam and progeny blood serum. J. Anim. Sci. 42:393.
Mavromatis J, G. Koptopoulos, S.C. Kyriakis, A. Papasteriadis, and K. Saoulidis. 1999. Effects of alpha-tocopherol and selenium on pregnant sows and their piglets' immunity and performance. Zentralbl Veterinarmed A. 46:545.
Moreira, I. and D.C. Mahan. 2002. Effect of dietary levels of vitamin E (all-rac-alpha-tocopheryl acetate) with or without added fat on weanling pig performance and tissue alpha-tocopherol concentration. J. Anim. Sci. 80:663.
Mutetikka, D.B and D.C. Mahan. 1993. Effect of pasture, confinement, and diet fortification with vitamin E and selenium on reproducing gilts and their progeny. J. Anim. Sci. 71:3211.
Nockels, C.F. 1979. Protective effects of supplemental vitamin E against infection. Fed. Proc. 38: 2134.
NRC. 1959. The Nutrient Requirements for Swine. National Academy of Sciences. Washington DC.
NRC. 1973. The Nutrient Requirements for Swine. National Academy of Sciences. Washington DC.
NRC. 1998. The Nutrient Requirements for Swine. National Academy of Sciences. Washington DC.
Rafai P, S. Tuboly, H. Biro, L. Jakab, Z. Papp and L. Molnar. 1989. Effect of selenium, vitamin E and riboflavin supplementation of the feed on the humoral- and cell-mediated immune responses of growing pigs. Acta Vet Hung. 37:201.
Specht, T.A., D.C. Mahan, N.D. Fastinger and R.L. Stuart. 2003. Effect of d-α-tocopherol alcohol or acetate in water soluble or emulsified form to the drinking water of weanling pig. J. Anim. Sci. 81(Suppl. 2):88 (Abst. 356).
Stone, W.L., I. LeClair, T. Ponder, G. Baggs and B.B. Reis. 2003. Infants discriminate between natural and synthetic vitamin E. Am. J. Clin. Nutr. 77:899.
Tengerdy, R.P. et al, 1984. Effect of vitamin E on immunity and disease resistance. In: Vitamins, Nutrition, and Cancer. K. Prasad, ed. Karger. Basel. p. 123-133.
Traber, M.G. 1999. Vitamin E. In: Shils, M.E., Olson J.A., Shike, M., Ross, A.C. eds. Modern Nutrition in Health and Disease, 9th Ed. Baltimore MD. Williams & Wilkins, p. 347.
Traber, M.G., G.W. Burton, L. Hughes, K.U. Ingold, H. Hiddaka, M. Malloy, J. Kane, J. Hyams and H.J. Kayden. 1992. Discrimination between forms of vitamin E by humans with and without genetic abnormalities of lipoprotein metabolism. J. Lipid Res. 33:1171.
United States Pharmacopeia. 1999. Rockville, MD.
Wuryastuti, H., H.D. Stowe, R.W. Bull and E.R. Miller. 1993. Effects of vitamin E and selenium on immune responses of peripheral blood, colostrum and milk leucocytes of sows. J. Anim. Sci. 71: 2464.











