1. Introduction
Among various metabolic pathways present in the organism, the metabolism of sulfur- (S) methionine and its seleno- (Se) analogous (Se-methionine) are particularly important because they not only contribute to protein mass but also produce (Se) homocysteine, a key metabolite connecting two fundamental metabolic functions, the one-carbon metabolism and the antioxidative system.
The transfer of one-carbon groups, represented by methyl (–CH3), methylene (–CH2–), formyl (–CHO), formimino (–CHNH), and methenyl (–CH=), is involved in the remethylation of (Se) homocysteine to (Se) methionine, as well as related pathways such as the folate cycle and the choline oxidation pathway [1]. Additionally, during the demethylation of (Se) methionine to (Se) homocysteine, the universal bioactive methyl donor S-adenosylmethionine (SAM) is synthesized and donates its methyl group to a large number of methyl acceptors catalyzed by methyltransferases [1,2] with profound impacts on DNA synthesis, protection and repair, cellular metabolism, and cell proliferation and, consequently, with direct effects on embryo/fetal growth [3,4].
During redox challenges, however, the high level of reactive oxygen species (ROS) induces a negative feedback on methionine synthase (MTR) [5] and betaine-homocysteine S-methyltransferase (BHMT) [6], which are enzymes that catalyze the regeneration of methionine from homocysteine [7]. Concomitantly, a positive feedback on cystathionine _-synthase (CBS), the first enzyme of the transsulfuration pathway, increases the flow of carbon towards transsulfuration reactions [8].
Although the transmethylation of (Se) methionine is not dependent on B6 [11], the direct impact of this vitamin on the transsulfuration of (Se) homocysteine to (Se) cysteine and the intrinsic relation between these two metabolic pathways make the B6 status of an individual important not only for the antioxidant system but for the one-carbon pool as well. Considering that the transsulfuration of homocysteine to the synthesis of GSH requires two B6-dependent reactions, whereas Se-homocysteine requires five B6-dependent reactions to synthesize SeGPX, the impact of B6 is expected to be more evident for Se metabolism. Additionally, the dissimilar metabolisms between organic and mineral Se, including different mechanisms of regulation for the synthesis of selenoprotein, indicate possible different fates for the one-carbon metabolism.
All the above concepts are even more fundamental in developing embryos because of the singular transfer of Se to pre-implantation embryos [12], the redox alterations brought by the placentation process [13], and the incomplete transsulfuration pathway [14].
The present review discusses the impact of B6 on the equilibrium between the synthesis and the consumption of one-carbon units under different oxidative stress conditions, focusing not only sulfur-related metabolism but also Se metabolism and the differences between Se sources both in adults and embryos at 5- and 30-days of gestation using a pig model.
2. Vitamin B6 Metabolism
Vitamers and Their Metabolism
Vitamin B6 is a general description of six interconvertible metabolites, pyridoxine (alcohol), pyridoxamine (amine), pyridoxal (aldehyde), and their respective phosphates. Its main metabolic form differs between plant (pyridoxine) and animal (pyridoxal and/or pyridoxamine) sources, but, independently of origin, they are mainly found in the phosphorylated state or bound to proteins [15].
The enteric absorption of the phosphorylated protein-bound B6 vitamers, mostly at jejunum and ileum, is dependent on their dephosphorylation by membrane-bound alkaline phosphatases present in the intestinal mucosa [16]. All three dephosphorylated vitamers are absorbed by passive diffusion, and, within the enterocytes, these metabolites are re-phosphorylated, generating a metabolic trapping of the vitamin with further oxidation to pyridoxal-5-phosphate (P-5-P), its metabolically active form [17]. This latter metabolite must be dephosphorylated at the intestinal serosal surface before its release into the portal circulation. Dephosphorylated forms of B6 are readily taken up by membranes, whereas the phosphorylated analogs are not; therefore, phosphorylation may be considered a mechanism for the intracellular retention of this vitamin [15].
Pyridoxal released into the portal circulation is absorbed in the liver by passive diffusion, followed by re-phosphorylation within the cells. In order to cross liver cell membranes, P-5-P is hydrolyzed to pyridoxal and released into the general circulation bound to albumin and/or hemoglobin [18]. Any P-5-P that is not bound to proteins is readily hydrolyzed and the free pyridoxal remaining in the liver is oxidized to 4-pyridoxic acid and excreted.
Considering that no specific tissular storage of B6 is present in the organism, both short- and long-term whole body pools of B6 (about 12 h and 1 month, respectively) are present as P-5-P bound to enzymes/proteins [15]. Blood plasma is the major source of extrahepatic B6, which occurs mainly as P-5-P bound to albumin [16]. Circulatory P-5-P must be hydrolyzed (by extracellular alkaline phosphatases) to pyridoxal that can cross cell membranes and then is trapped intracellularly by phosphorylation. Pyridoxal-5-P is over six times more concentrated in erythrocytes than in plasma possibly because the Schiff base with hemoglobin is stronger than the one with albumin, driving the uptake of the vitamin to erythrocytes; however, its storage in these cells is saturable [16,19].
3. Vitamin B6 Metabolic Functions
In addition to the role of B6 as a cofactor for the degradation of stored carbohydrates [20] and its function as a protective agent against ROS generated in vitro [21], P-5-P was shown to interact with steroid hormone-receptors, affecting their genes expression [22]. Pyridoxal-5-P also acts as a cofactor in the biosynthesis of many neurotransmitters such as dopamine, epinephrine, gamma-aminobutyric acid, histamine, norepinephrine, and serotonin, as well as the neuromodulator serine [17]. However, the most recognized role of P-5-P is the catalysis of many important steps in the metabolism of amino acids, such as transamination, racemization, decarboxylation, and α,β-elimination reactions [23,24]. The various reactions of P-5-P in the metabolism of amino acids depend on its ability to stabilize amino acid carbanions. In the absence of a substrate, P-5-P forms an internal aldimine (Schiff base) with the lysyl residue (ε-amino group) of the enzyme. Once a substrate amino acid displaces its lysyl residue (α-amino group), P-5-P transfers the aldimine linkage from the ε-amino group to the α-amino group.
4. Transmethylation vs. Transsulfuration
4.1. Transmethylation and the One-Carbon Metabolism
Methylation is an essential metabolic function that controls the addition of –CH3 groups to a variety of organic compounds in every cell of the body. Transmethylation represents metabolic reactions in which –CH3 groups are transferred from one compound to another, comprising both demethylation and remethylation reactions. The methionine cycle contains good examples of transmethylation reactions, in which methionine is demethylated to homocysteine, S-adenosylmethionine acts as a methyl donor, with a further possible remethylation to methionine with either 5-methyl tetrahydrofolate or betaine as the methyl donor.
During the methionine cycle, the –CH3 group of methionine, which contains a sulfur atom, is activated by adenosine triphosphate (ATP) through the addition of adenosine to its sulfur. This reaction, which is catalyzed by methionine adenosyltransferase (MAT), forms S-adenosylmethionine (SAM) [25], which is an important donor of –CH3 groups to nucleic acids, proteins, and neurotransmitters.
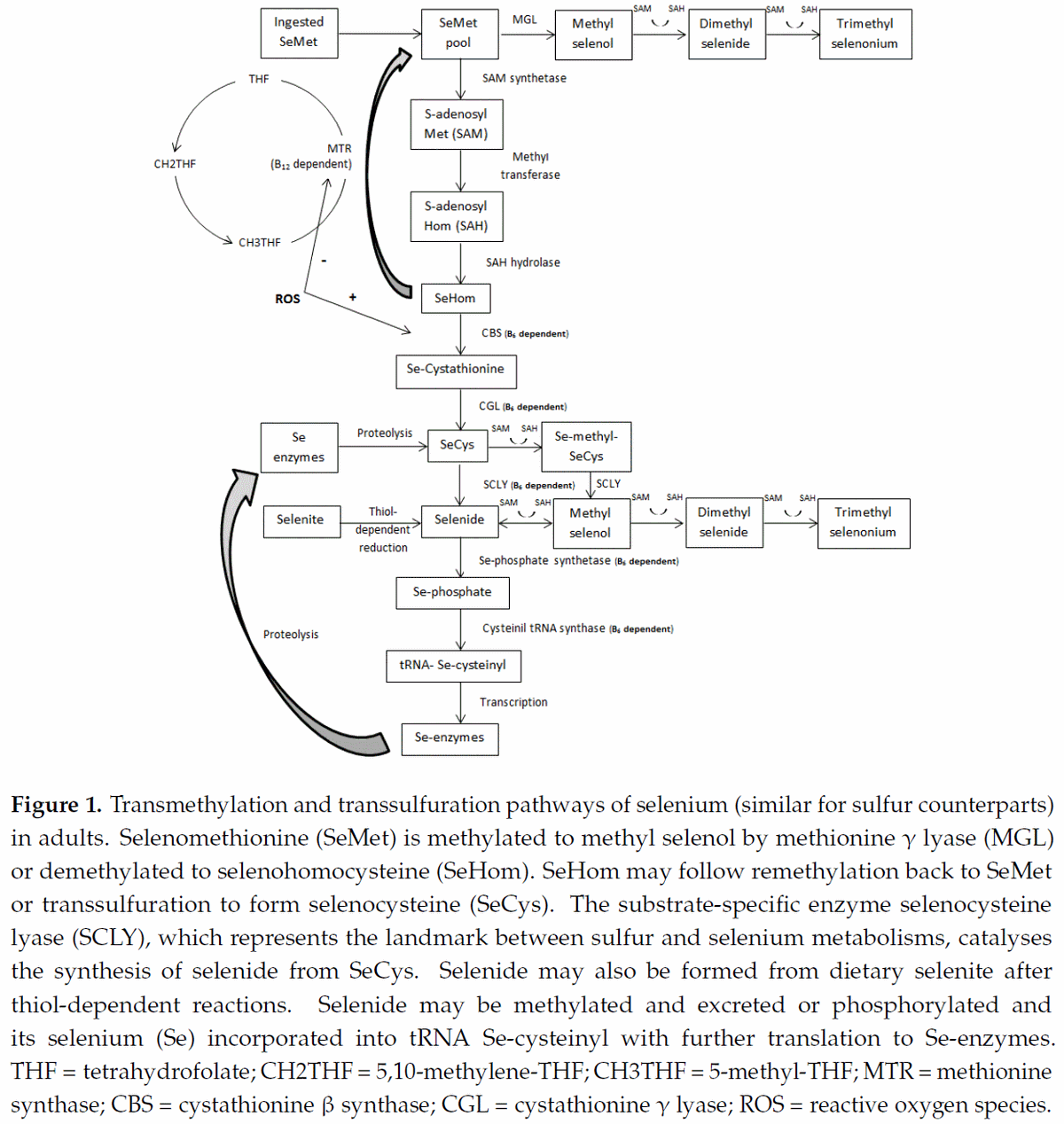
Upon transfer of its –CH3 group, SAM is rapidly converted to S-adenosylhomocysteine (SAH) that, by removal of the adenosine molecule catalyzed by SAH hydrolase (SAHH), is immediately hydrolyzed to homocysteine [26]. Homocysteine can then follow two major pathways; transsulfuration (see next sub-item) or remethylation (Figure 1). Two remethylation pathways regenerate methionine; one is independent of cobalamin (Cbl) but depends on betaine as the one-carbon donor, and the other is Cbl-dependent and requires folate (5-methyltetrahydrofolate) as the one-carbon donor [27].
Folate has two carbon-carbon double bonds that yield dihydrofolate (DHF) and tetrahydrofolate (THF) after saturation of its first and second carbon, respectively, by hydrogen. Folates serve as donors of single carbons in their reduced (5-methyl-THF; CH3THF), intermediate (5,10-methylene-THF; CH2THF), and oxidized (10-formyl-THF; CHOTHF) states [28]. Although not clearly shown in the literature [11,29,30], B6 status may influence the flux –CH3 groups through remethylation because 5,10-methylene-THF is formed by the methylene group of the side-chain of serine (after its conversion into glycine) along with THF in a reaction catalyzed by the B6-dependent enzyme serine hydroxylmethyltransferase. Although THF and serine, respectively, are the most important metabolic carrier and source of one-carbon groups, they do not promote the most energetically favorable reactions. The interaction of methionine with ATP produces SAM that easily donates its –CH3 group; SAM is considered the most important one-carbon donor [3]. Folate and SAM transfer of single carbons, which are volatile and bind easily to other molecules, generates the one-carbon metabolism. The one-carbon metabolism creates interplay between amino acid and nucleotide metabolisms, playing a fundamental role in DNA synthesis, repair, and replication [3]. The one-carbon donor 5-methyl-THF is used to convert homocysteine into methionine. The overall reaction transforms 5-methyl-THF into THF while transferring a –CH3 group to homocysteine to form methionine [4].
Methionine synthase (MTR), a vitamin B12 (Cbl)-dependent enzyme, catalyzes the final step in the regeneration of methionine from homocysteine [7]. The complex Cbl(I)MTR binds the –CH3 group of 5-methyl-THF to form methylCbl(III)MTR, activating the enzyme. The activated –CH3 group is transferred from methylCbl(III)MTR (regenerating Cbl(I)MTR) to homocysteine synthesizing methionine, which is released from the enzyme [31]. Under folate and/or vitamin B12 deficiency, MTR reactions are severely impaired.
4.2. Transsulfuration and the GPX System
As described above, after its synthesis from SAH, homocysteine can follow the remethylation or the transsulfuration pathway (Figure 1). Transsulfuration is a metabolic pathway involving the conversion of homocysteine into cysteine through the intermediate metabolite cystathionine. Briefly, in a reaction catalyzed by cystathionine β-synthase (CBS; B6-dependent enzyme), a β-replacement of the thiol group of homocysteine by the acetyl or succinyl group of homoserine forms cystathionine [32]. By acting at the homocysteine junction, CBS represents a critical step, regulating both the maintenance of the methionine pool and the synthesis of cysteine. Therefore, this enzyme would be expected to be strictly regulated. In fact, CBS is allosterically regulated by SAM, in which low SAM concentrations direct homocysteine into remethylation, whereas at high SAM concentrations, transsulfuration is favored [33]. Following cystathionine formation, the B6-dependent enzyme cystathionine γ-lyase (CGL) cleaves the molecule by γ-elimination of its homocysteine portion. This reaction leaves an unstable amino acid that binds to water molecules to form cysteine, _-ketobutyrate, and ammonia [34]. By the action of gamma-glutamylcysteine synthetase (GCL), cysteine and glutamate synthesize gamma-glutamylcysteine, a rate-limiting step in GSH synthesis [35]. Further, glycine binds to the C-terminal of gamma-glutamylcysteine via the enzyme GSH synthetase to form GSH.
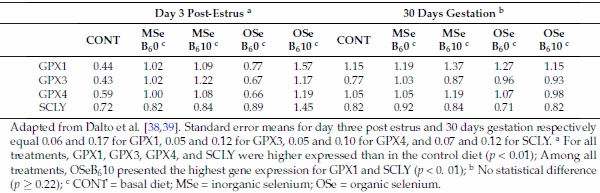
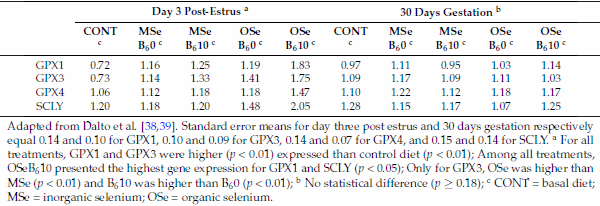
Therefore, the transsulfuration pathway is a straight connection between homocysteine and GSH, the major redox buffer in mammalian cells. Consequently, it is expected that enzymes related to this metabolism display sensitivity to redox changes. Indeed, studies on purified mammalian MTR and CBS have revealed the reciprocal sensitivity of these two major homocysteine-utilizing enzymes to oxidative conditions [5,36]. In mammals, CBS contains a heme cofactor that functions as a redox sensor, increasing CBS activity and consequently transsulfuration under oxidizing conditions [36]. In contrast, under these same conditions, remethylation is depressed because MTR activity is reduced, most likely due to the lability of the reactive cofactor intermediate Cbl(I) [5]. Additionally, the betaine-related enzyme BHMT was also shown to be inhibited by oxidizing agents [6]. Considering that mammals metabolize seleno-amino acids in the same way as their sulfur counterparts [37], recent studies by this laboratory on the effects of selenium (Se) sources and levels combined or not with B6 provided indirect support to this effect of ROS. Dalto et al. [38] reported that, under the oxidative stress of ovulation, the gene expression of GPX1, 3, and 4 in the livers (Table 1) and kidneys (Table 2) of Se-supplemented animals were higher than in the control Se-unsupplemented group, and animals supplemented with organic Se plus B6 had the highest gene expression of GPX1 and selenocysteine lyase (SCLY), indicating that the transsulfuration pathway was stimulated. In contrast, Dalto et al. [39] observed, under basal oxidative stress conditions, no differential expression for the same genes (Tables 1 and 2).
Table 1. Real-time mRNA abundance of liver glutathione peroxidase (GPX) and selenocysteine-lyase (SCLY) genes in gilts three days after the fourth estrus and at day 30 of gestation, according to selenium and vitamin B6 treatments.
Adapted from Dalto et al. [38,39]. Standard error means for day three post estrus and 30 days gestation respectively equal 0.06 and 0.17 for GPX1, 0.05 and 0.12 for GPX3, 0.05 and 0.10 for GPX4, and 0.07 and 0.12 for SCLY. a For all treatments, GPX1, GPX3, GPX4, and SCLY were higher expressed than in the control diet (p < 0.01); Among all treatments, OSeB610 presented the highest gene expression for GPX1 and SCLY (p < 0. 01); b No statistical difference (p ≥ 0.22); c CONT = basal diet; MSe = inorganic selenium; OSe = organic selenium.
Table 2. Real-time mRNA abundance of kidney glutathione peroxidase (GPX) and selenocysteine-lyase (SCLY) genes in gilts three days after the fourth estrus and at day 30 of gestation, according to selenium and vitamin B6 treatments.
Adapted from Dalto et al. [38,39]. Standard error means for day three post estrus and 30 days gestation respectively equal 0.14 and 0.10 for GPX1, 0.10 and 0.09 for GPX3, 0.14 and 0.07 for GPX4, and 0.15 and 0.14 for SCLY. a For all treatments, GPX1 and GPX3 were higher (p < 0.01) expressed than control diet (p < 0.01); Among all treatments, OSeB610 presented the highest gene expression for GPX1 and SCLY (p < 0.05); Only for GPX3, OSe was higher than MSe (p < 0.01) and B610 was higher than B60 (p < 0.01); b No statistical difference (p ≥ 0.18); c CONT = basal diet; MSe = inorganic selenium; OSe = organic selenium.
5. The Role of B6
Differential Effects on Organic and Mineral Se Metabolisms in Gilts
Selenium is an essential trace element derived from inorganic (MSe) or organic (OSe) sources. Both forms are involved in the activation of SeGPX. The metabolism of Se-methionine, the natural organic source present in food, is interchangeable with a sulfur-methionine metabolism [37], and, therefore, the influence of B6 on the one-carbon metabolism by the regulation between remethylation and transsulfuration is predictable. Although less evident, selenide, which is the metabolized form of selenite (commonly used dietary MSe) and a key intermediate for the utilization and/or excretion of both OSe and MSe [40,41], may represent another regulatory step influencing one-carbon metabolism. In this sense, two important B6-dependent reactions direct selenide to be incorporated into Se-enzymes in preference to its excretion through the use of one-carbon groups in methylation reactions.
For Se-methionine, it is converted to Se-homocysteine through the action of SAM synthetase and SAHH, supplying the one-carbon system with –CH3 groups (Figure 1). As discussed above for sulfur-homocysteine, Se-homocysteine may be remethylated to Se-methionine or transsulfurated to Se-cystathionine, depending on the influence of SAM levels and ROS feedback on CBS. In this context, the SAM levels may promote equilibrium between remethylation and transsulfuration depending on dietary OSe levels, whereas the positive feedback of ROS on CBS (and the negative feedback of MTR) favors the transsulfuration pathway independently of dietary Se levels. Se-methionine may also be transaminated to methylselenol and then transformed to selenide via methyltransferases [42] or methylated to excretory forms. However, considering the importance of this amino acid in protein synthesis and the vital consequences of transmethylation, the partition of its pool to transamination and methylation may have a secondary relevance.
After SeCys synthesis from Se-cystathionine via Se-cystathionine gamma lyase (CGL; B6-dependent enzyme), this amino acid can be incorporated into proteins or metabolized by the B6-dependent enzyme SeCys lyase (SCLY) to alanine and selenide [43]. Considering that SCLY is substrate specific, this step is the landmark between sulfur and Se metabolisms and settles their fates in relation to the metabolism of glutathione. It has to be stated that the greater volume of sulfur, rather than SeCys, molecules available in the organism makes the impact of B6 important not only for the enzyme SeGPX but also for its substrate (glutathione) as well.
Dietary selenite is non-enzymatically reduced via thiol-dependent reactions to selenide [44]. This direct reduction of selenite, short-cutting the transsulfuration pathway and with no regulatory mechanism, allows the accumulation of toxic levels of selenide. To avoid toxicity, selenide from both OSe (synthesized from SeCys) and MSe may follow two pathways: synthesis of Se-proteins or methylation (excretion). By the action of Se-phosphate syntethase 2 (B6-dependent enzyme), selenite is converted into Se-phosphate [45]. This last molecule acts as a Se donor in the exchange of the phosphate moiety of serine-tRNA for Se, generating the SeCys-specific tRNA(Ser)Sec, in a B6-dependent reaction [46]. This special tRNA, which contains the SeCys-insertion sequence (SECIS) along with a SeCys-tRNA-specific elongation factor (eEFSEC) and a specific SECIS binding protein (SECISBP2), is essential for SeCys incorporation into Se-proteins [47]. Remaining selenide must be successively methylated using –CH3 groups from SAM to generate the monomethylated intermediate methylselenol and multimethylated excretory metabolites (dimethylselenide and trimethylselenonium) [48].
Although the metabolism of OSe also generates selenide, the balance between SeCys incorporation into protein and its degradation by the saturable enzyme SCLY prevents the accumulation of excessive selenide. It is known that methylation and demethylation reactions between selenide and methylselenol promote equilibrium between the two molecules [41,42,49]; however, considering that low levels of dietary selenite supplementation provoke toxicity, whereas dietary Se-methionine is tolerated up to extreme high levels [50], one can assume that the utilisation of dietary selenite will direct the metabolism to consume greater amounts of one-carbon groups for the methylation of selenide than dietary Se-methionine. In contrast, OSe metabolism may not only preserve –CH3 molecules through the controlled synthesis of selenide, but also acts generating these one-carbon groups via demethylation/remethylation reactions between Se-methionine and Se-homocysteine. In this context, the regulatory steps (SAM levels, ROS feedback on CBS and MTR, and controlled synthesis of selenide from SCLY) in this metabolic pathway may promote a balance between the synthesis of one-carbon molecules and Se-proteins with possible beneficial effects to the organism.
Recent studies on Se and B6 metabolisms by this laboratory, using the peri-estrus period as a model for oxidative stress in gilts [38,39,51], provided peculiar information about the control of this metabolic pathway. Although those studies did not specifically address the homeostasis of –CH3 groups, based on the above information this review makes some indirect inferences about the one-carbon metabolism from responses reported in relation to the transsulfuration pathway.
According to Dalto et al. [38,39], B6 supplementation (10 mg/kg) does not affect the deposition of blood or tissue Se on a long-term basis (3 to 5 estrus), as well as blood Se levels during the peri-estrus period (oxidative stress condition) for both MSe and OSe. For MSe it is not surprising because, independently of the physiological state, absorbed selenite is quickly transformed into selenide and then converted into functional Se-proteins or methylated instead of being actively stored [9]. Consequently, the impact of MSe on the pool of one-carbon molecules for the methylation of selenide would be mainly related to the level of selenite supplementation rather than to B6 availability.
For OSe supplemented animals, however, the explanation is not so straightforward. Under basal oxidative stress conditions, the absence of positive feedback of ROS on CBS suggests that the remethylation of Se-homocysteine would be the preferential pathway, whereas the high SAM levels would stimulate CBS to transsulfurate Se-homocysteine. However, according to Bekaert et al. [52], in humans, high doses of daily supplementation with selenium yeast (a source of OSe) over six months did not affect the homocysteine concentrations in plasma. Also, considering that CBS is a B6-dependent enzyme, its activity would direct more Se to SeGPX synthesis under B6 supplementation. Davis [53] and Lima [54] reported that CBS is slightly affected by marginal vitamin B6 status, whereas CGL is more sensitive, but neither of them affected cysteine synthesis. Although under marginal B6 levels, these studies indicate that the B6 status may not be a determining factor of the transsulfuration pathway flux, which is possibly mainly controlled by ROS and SAM levels. In fact, Dalto et al. [39] reported no effect of B6 on either blood SeGPX activity or GPX and SCLY gene expression in gilts at 30-days of gestation (Tables 1 and 2), a condition shown to be of low oxidative stress in this species. These results suggest that under basal oxidative stress conditions, in animals supplemented with Se and B6, neither remethylation nor transsulfuration but Se deposition into proteins is the major route for this mineral.
In contrast, as also described above, Dalto et al. [38] reported that, under the oxidative stress of the peri-estrus period (3-days after estrus), GPX’s and SCLY genes were highly expressed in OSe + B6 supplemented animals in the liver and kidneys, in agreement with the positive feedback of ROS on CBS, increasing the ratio of transsulfuration. Interestingly, however, these same authors did not find any B6 effect on SeGPX activity in the liver and kidneys (day 3 after estus) or whole blood (day ?? 4 to day + 3 of the peri-estrus period). In this regard, Lubos et al. [55] reported that GPX can be regulated by transcriptional, post-transcriptional, translational, or post-translational factors, and changes in gene transcription may not be reflected on GPX protein or its related enzymatic activity.
6. Embryo Metabolism
6.1. 5-Days Porcine Embryos
Using microarray technology to evaluate the global gene expression in 5-days porcine embryos, Dalto et al. [56] provided unique and interesting information on Se and B6 metabolisms and, consequently, on the distinct interplay between remethylation and transsulfuration during early porcine embryo development.
The first aspect to be considered in 5-days embryos is the transfer of Se from dam to embryos. Dalto et al. [56] showed that Se was undetectable in the uterine flushing, indicating that this is a negligible source of this mineral for the pre-implantation embryo. Nevertheless, embryos from OSe supplemented dams are richer in Se than those from MSe supplemented dams. It was hypothesized that the most probable source is the pre-ovulatory oocyte that might be affected by the systemic maternal blood Se concentration through follicular fluid, considering that OSe is known to be more common deposited in tissues than is MSe [39,51,57,58]. In fact, Dalto et al. [56] observed that maternal supplementation with OSe plus B6 stimulated 28.8 times more genes than MSe plus B6 (six and 173 genes for MSe and OSe, respectively). These findings suggest that embryos coming from OSe supplemented dams have greater Se-methionine reserves and, therefore, are more suitable to go through demethylation steps to Se-homocysteine than are embryos from MSe supplemented dams.
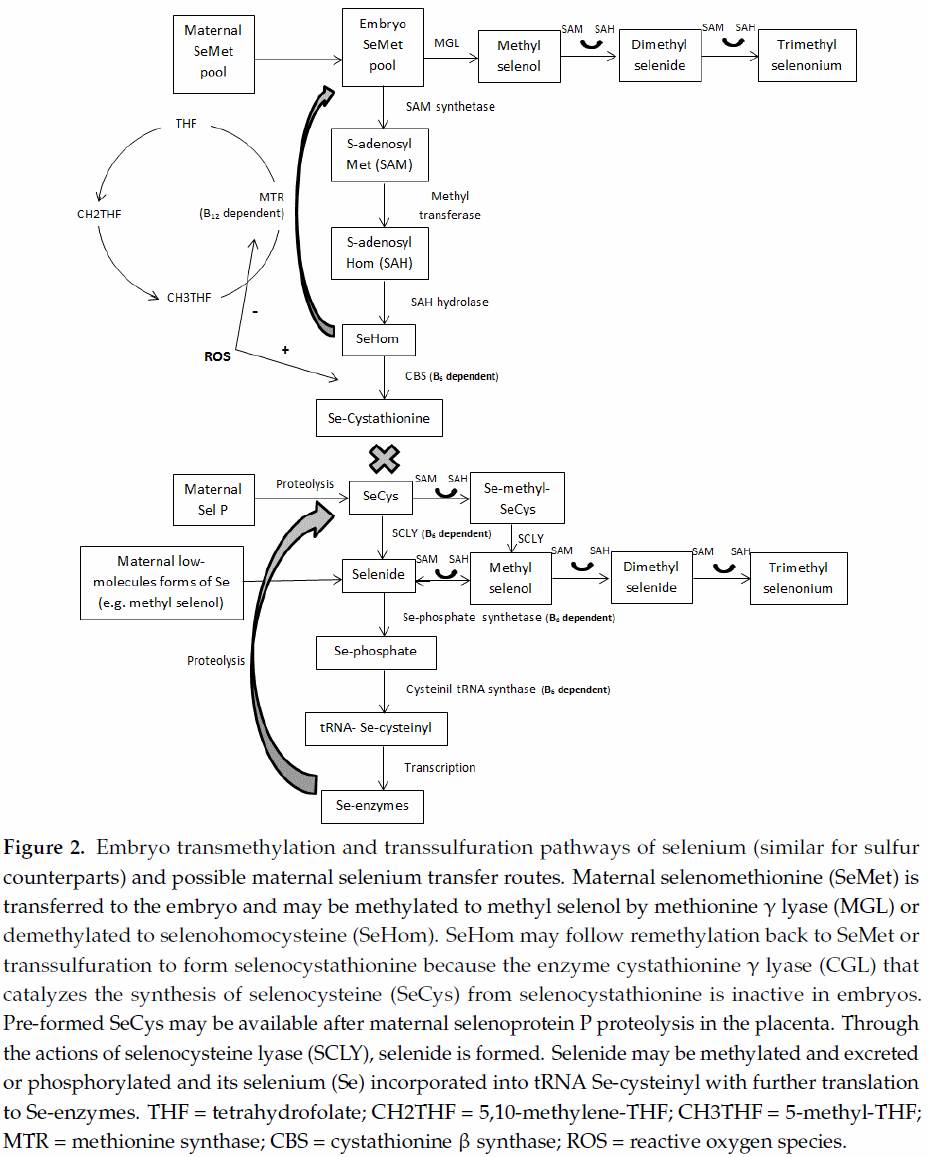
A crucial aspect when studying the remethylation/transsulfuration pathway in embryos, fetuses, and newborns is the absence of CGL activity, in spite of its mRNA expression [10,14] (Figure 2). This relevant finding implies that, from conception to neonatal age, individuals are not able to convert Se-methionine into SeCys via the transsulfuration pathway. Therefore, Se-methionine would be directly incorporated into protein, methylated to methylselenol (with further methylation to excretory forms or demethylation to selenide), or demethylated to Se-homocysteine (generating the –CH3 donor SAM) and thereafter remethylated to Se-methionine via folate-dependent reactions, supplying the one-carbon metabolism. Dalto et al. [56] evaluated the expression of genes exclusively related to each source of Se (common genes between sources were excluded from the analysis) in 5-days embryos from gilts supplemented with OSe or MSe combined with B6 and did not find differentially expressed genes related to the demethylation or remethylation of Se-methionine, whereas several genes related to general elongation factors and biological processes related to translation and mitotic cell cycle were stimulated by OSe. Those authors concluded that, in 5-days porcine embryos, Se-methionine preferentially follows protein deposition. However, recent results from this laboratory (data not yet published), using the database generated by Dalto et al. [56], evaluated global gene expression in 5-days embryos (common genes between sources of Se were not excluded from the analysis), showing that OSe, but not MSe, may affect DNA methylation and epigenetic events in 5-days embryos through the higher generation of –CH3 groups (stimulation of methyltransferases and SAM carrier genes). Therefore, although Se-methionine may be preferentially incorporated into protein, the magnitude of the demethylation and remethylation reactions that compose the one-carbon metabolism cannot be disregarded.
Although the transsulfuration pathway is not complete in embryos, many genes involved in Se-enzymes synthesis were found in Dalto et al. [56] suggesting that 5-days embryos are potentially capable of synthesizing these enzymes. If so, the most probable sources of Se to the embryo are SeCys, methylselenol, and/or Se-proteins (after proteolysis in the embryo) coming from the pre-ovulatory oocyte. Furthermore, SeGPX synthesis would possibly not be under the feedback control by ROS but limited by the B6-dependent enzyme SCLY, which controls the synthesis of selenide and, consequently, the consumption of one-carbon molecules for its methylation (Figure 2). Considering the importance of these one-carbon molecules for DNA processing and genetic stability, these routes of maternal Se transfer to early embryos may be considered a mechanism of defense to protect the pool of –CH3.
Similarly to 5-days embryos, the enzyme CGL is absent in 30-days embryos and, consequently, the above aspects related to remethylation/transsulfuration are expected to be analogous. However, at 30 days of gestation the presence of the placenta may change the dynamic of the interaction between these metabolic pathways.
One of the main features brought about by the presence of the placenta is the rise in the oxygen tension in the embryo, increasing the production of ROS. These levels are, in fact, higher in early gestation than thereafter [13], possibly because of the more immature embryonic antioxidant system. This suggests that, under oxidative stress conditions, the feedback of ROS stimulates CBS, directing Se-homocysteine to transsulfuration, whereas it would affect remethylation by reducing MTR activity. According to Kalhan [2], alterations to the metabolism of one-carbon may be the main cause of impaired fetal growth. Additionally, because the transsulfuration pathway is interrupted at Se-cystathionine, large amounts of this metabolite could possibly accumulate with negative effects in embryos from OSe supplemented dams. In this sense, under these conditions, B6 supplementation levels might be carefully considered due to its effect on CBS activity. Evidences of this negative effect of B6 on embryo development were observed in a study by Dalto et al. [39], in which maternal supplementation with 12.4 mg/kg of feed of B6, with either MSe or OSe at 0.6 mg/kg of feed, increased the within-litter Se content variation and the within-litter weight variation, compared to animals receiving 2.4 mg/kg of B6. Under similar experimental conditions and pig genetic lines, Fortier et al. [51] fed gilts with different sources of Se (MSe or OSe at 0.5 mg/kg of feed) but a fixed amount of B6 at 3 mg/kg and observed improved within-litter Se content variation, embryo weight and length, and protein and DNA content, with no detrimental effect on the within-litter weight variation, compared to the control diet at similar levels of B6 but 0.2 mg/kg of feed of Se.
As mentioned above, OSe increases body Se concentration more than MSe due to its deposition in proteins following the methionine metabolism, and B6 does not interfere with Se deposition in the tissues of adult individuals [38,39]. Also in 30-days porcine embryos, Fortier et al. [51] and Dalto et al. [39] showed that, independently of B6 status, gilts supplemented with OSe produced embryos with higher Se concentrations than gilts supplemented with MSe. Considering that the transsulfuration pathways is not complete at this stage of development, this higher load of Se-methionine in OSe embryos is not utilized in the synthesis of selenoenzymes but rather will be deposited in proteins and/or transmethylated, producing one-carbon groups. However, it has to be stated that even MSe supplemented gilts supply their embryos with OSe, not as Se-methionine but as SeCys, after selenoprotein P catabolism in the placenta [12]. In spite of the low activity of SCLY in vivo [59], one can assume that the majority of SeCys would be metabolized through SCLY because its concentration in tissues is enough to metabolize all SeCys available and the KM (Michaelis constant) value of the enzyme is greater than the tissue levels of the substrate. However, Fortier et al. [51] and Dalto et al. [39] reported no effect of maternal Se levels (control vs. Se-supplemented gilts) and sources (OSe vs. MSe) on SeGPX activity in 30-days porcine embryos, indicating that at this stage of development the enzyme flux is not primarily substrate driven. Therefore, it is reasonable to hypothesize that embryo SeCys, from the catabolism of selenoprotein P in the placenta, follows primarily protein deposition and secondarily catabolism by SCLY, which is not controlled by the feedback of ROS, for the synthesis of selenide and further selenoenzymes. This mechanism would protect the embryo from wasting one-carbon groups for the methylation and elimination of excess selenide.
7. Conclusions
Both sulfur and Se methionine are important metabolic suppliers of one-carbon molecules through transmethylation reactions. The equilibrium between transmethylation and transsulfuration pathways, majorly regulated by redox conditions on CBS and MTR, influences the flow of –CH3 molecules between the one-carbon and antioxidation metabolisms.
Vitamin B6 is important for the interplay between the synthesis (transmethylation) and the utilization (transsulfuration) of one-carbon groups by acting in most of their regulatory enzymes. However, its effects on the GPX system depend on other parameters such as the oxidative stress conditions and metabolic maturity.
Finally, whereas MSe may act by wasting one-carbon molecules to methylate the excess of selenide produced, OSe not only preserves one-carbon groups (due to the control of SCLY) but also promotes equilibrium between transsulfuration and transmethylation via the control of B6-dependent CBS by SAM and ROS.
Author Contributions: Both authors equally contributed to the writing of the manuscript, revision of its content and approval of the final version submitted for publication.
Conflicts of Interest: The authors declare no conflict of interest.
This article was originally published in Nutrients 2017, 9, 189; doi:10.3390/nu9030189. This is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/). References
1. Joseph, J.; Loscalzo, J. Methoxistasis: Integrating the roles of homocysteine and folic acid in cardiovascular pathobiology. Nutrients 2013, 5, 3235–3256.
2. Kalhan, S.C. One carbon metabolism in pregnancy: Impact on maternal, fetal and neonatal health. Mol. Cell. Endocrinol. 2016, 435, 48–60.
3. Selhub, J. Homocysteine metabolism. Annu. Rev. Nutr. 1999, 19, 217–246.
4. Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry, 5th ed.; W.H. Freeman and Company: New York, NY, USA, 2002; pp. 571–572, 674–678, 686, 694, 698–700, 704, 771–772.
5. Chen, Z.; Chakraborty, S.; Banerjee, R. Demonstration that mammalian methionine synthases are predominantly cobalamin-loaded. J. Biol. Chem. 1995, 270, 19246–19249.
6. Castro, C.; Millian, N.S.; Garrow, T.A. Liver betaine-homocysteine S-methyltransferase activity undergoes a redox switch at the active site zinc. Arch. Biochem. Biophys. 2008, 472, 26–33.
7. Li, Y.N.; Gulati, S.; Baker, P.J.; Brody, L.C.; Banerjee, R.; Kruger, W.D. Cloning, mapping and RNA analysis of the human methionine synthase gene. Hum. Mol. Genet. 1996, 5, 1851–1858.
8. Mosharov, E.; Cranford, M.R.; Banerjee, R. The quantitatively important relationship between homocysteine metabolism and glutathione synthesis by the transsulfuration pathway and its regulation by redox changes. Biochemistry 2000, 39, 13005–13011.
9. Windisch,W. Interaction of chemical species with biological regulation of the metabolism of essential trace elements. Anal. Bioanal. Chem. 2002, 372, 421–425.
10. Gardiner, C.S.; Reed, D.J. Glutathione redox cycle-driven recovery of reduced glutathione after oxidation by tertiary-butyl hydroperoxide in preimplantation mouse embryos. Arch. Biochem. Biophys. 1995, 321, 6–12.
11. Davis, S.; Scheer, J.; Quinlivan, E.; Coats, B.; Stacpoole, P.; Gregory, J. Dietary vitamin B6 restriction does not alter rates of homocysteine remethylation or synthesis in healthy young women and men. Am. J. Clin. Nutr. 2005, 81, 648–655.
12. Burk, R.F.; Olson, G.E.; Hill, K.E.;Winfrey, V.P.; Motley, A.K.; Kurokawa, S. Maternal-fetal transfer of selenium in the mouse. FASEB J. 2013, 27, 3249–3256.
13. Jauniaux, E.; Watson, A.L.; Hempstock, J.; Bao, Y.P.; Skepper, J.N.; Burton, G.J. Onset of maternal arterial blood flow and placental oxidative stress. A possible factor in human early pregnancy failure. Am. J. Pathol. 2000, 157, 2111–2122.
14. Levonen, A.L.; Lapatto, R.; Saksela, M.; Raivio, K.O. Human cystathionine gamma-lyase: Developmental and in vitro expression of two isoforms. Biochem. J. 2000, 347, 291–295.
15. Bender, D.A. Nutritional Biochemistry of the Vitamins, 2nd ed.; Cambridge University Press: Cambridge, UK, 2003.
16. Combs, G.F., Jr. The Vitamins, 4th ed.; Academic Press: San Diego, CA, USA, 2012.
17. Combs, G.F. The Vitamins: Fundamental Aspects in Nutrition and Health; Elsevier: San Diego, CA, USA, 2008.
18. Berdanier, C.D.Water-Soluble Vitamins. In Advanced Nutrition-Micronutrients; Berdanier, C.D., Ed.; CRC Press: New York, NY, USA, 1998.
19. Matte, J.J.; Girard, C.L.; Sève, B. Effects of long-term parenteral administration of vitamin B6 on B6 status and some aspects of the glucose and protein metabolism of early-weaned piglets. Br. J. Nutr. 2001, 85, 11–21.
20. Helmreich, E.J. How pyridoxal 50-phosphate could function in glycogen phosphorylase catalysis. Biofactors 1992, 3, 159–172.
21. Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000, 71, 129–134.
22. Oka, T. Modulation of gene expression by vitamin B6. Nutr. Res. Rev. 2001, 14, 257–265.
23. Drewke, C.; Leistner, E. Biosynthesis of vitamin B6 and structurally related derivatives. Vitam. Horm. 2001, 61, 121–155.
24. Mittenhuber, G. Phylogenetic analyses and comparative genomics of vitamin B6 (pyridoxine) and pyridoxal phosphate biosynthesis pathways. J. Mol. Microbiol. Biotechnol. 2001, 3, 1–20.
25. Mudd, S.H.; Cantoni, G.L. Activation of methionine for transmethylation. III. The methionine-activating enzyme of Bakers’ yeast. J. Biol. Chem. 1958, 231, 481–492.
26. Ulrey, C.L.; Liu, L.; Andrews, L.G.; Tollefsbol, T.O. The impact of metabolism on DNA methylation. Hum. Mol. Genet. 2005, 14.
27. Födinger, M.; Hörl,W.; Sunder-Plassmann, G. Molecular biology of 5,10-methylenetetrahydrofolate reductase. J. Nephrol. 2000, 13, 20–33.
28. Tibbetts, A.S.; Appling, D.R. Compartmentalization of mammalian folate-mediated one-carbon metabolism. Annu. Rev. Nutr. 2010, 30, 57–81.
29. Martinez, M.; Cuskelly, G.J.; Williamson, J.; Toth, J.P.; Gregory, J.F. Vitamin B6 deficiency in rats reduces hepatic serine hydroxymethyltransferase and cystathionine beta-synthase activities and rates of in vivo protein turnover, homocysteine remethylation and transsulfuration. J. Nutr. 2000, 130, 1115–1123.
30. Perry, C.; Yu, S.; Chen, J.; Matharu, K.; Stover, P. Effect of vitamin B6 availability on serine hydroxymethyltransferase in MCF-7 cells. Arch. Biochem. Biophys. 2007, 462, 21–27.
31. Matthews, R.G.; Smith, A.E.; Zhou, Z.S.; Taurog, R.E.; Bandarian, V.; Evans, J.C.; Ludwig, M. Cobalamin-dependent and cobalamin-independent methionine synthases: Are there two solutions to the same chemical problem? Helv. Chim. Acta 2003, 86, 3939–3954.
32. Aitken, S.M.; Lodha, P.H.; Morneau, D.J.K. The enzymes of the transsulfuration pathways: Active-site characterizations. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 1511–1517.
33. Ereno-Orbea, J.; Majtan, T.; Oyenarte, I.; Kraus, J.P.; Martinez-Cruz, L.A. Structural insight into the molecular mechanism of allosteric activation of human cystathionine beta-synthase by S-adenosylmethionine. Proc. Natl. Acad. Sci. USA 2014, 111, E3845–E3852.
34. Flavin, M.; Slaughter, C. Cystathionine cleavage enzymes of neurospora. J. Biol. Chem. 1964, 239, 2212–2219.
35. Franklin, C.C.; Backos, D.S.; Mohar, I.; White, C.C.; Forman, H.J.; Kavanagh, T.J. Structure, function, and post-translational regulation of the catalytic and modifier subunits of glutamate cysteine ligase. Mol. Aspects Med. 2009, 30, 86–98.
36. Taoka, S.; Ohja, S.; Shan, X.; Kruger, W.D.; Banerjee, R. Evidence for heme-mediated redox regulation of human cystathionine _-synthase activity. J. Biol. Chem. 1998, 273, 25179–25184.
37. Schrauzer, G.N. The nutritional significance, metabolism and toxicology of selenomethionine. Adv. Food Nutr. Res. 2003, 47, 73–112.
38. Dalto, D.B.; Roy, M.; Audet, I.; Palin, M.-F.; Guay, F.; Lapointe, J.; Matte, J.J. Interaction between vitamin B6 and source of selenium on the response of the selenium-dependent glutathione peroxidase system to oxidative stress induced by oestrus in pubertal pig. J. Trace Elem. Med. Biol. 2015, 32, 21–29.
39. Dalto, D.B.; Audet, I.; Lapointe, J.; Matte, J.J. The importance of pyridoxine for the impact of the dietary selenium sources on redox balance, embryo development, and reproductive performance in gilts. J. Trace Elem. Med. Biol. 2016, 34, 79–89.
40. Birringer, M.; Pilawa, S.; Flohe, L. Trends in selenium biochemistry. Nat. Prod. Rep. 2002, 19, 693–718.
41. Suzuki, K.T.; Doi, C.; Suzuki, N. Metabolism of 76Se-methylselenocysteine compared with that of 77Se-selenomethionine and 82Se-selenite. Toxicol. Appl. Pharmacol. 2006, 217, 185–195.
42. Suzuki, K.T.; Kurasaki, K.; Suzuki, N. Selenocysteine _-lyase and methylselenol demethylase in the metabolism of Se-methylated selenocompounds into selenide. Biochim. Biophys. Acta 2007, 1770, 1053–1061.
43. Esaki, N.; Nakamura, T.; Tanaka, H.; Soda, K. Selenocysteine lyase, a novel enzyme that specifically acts on selenocysteine. Mammalian distribution and purification and properties of pig liver enzyme. J. Biol. Chem. 1982, 257, 4386–4391.
44. Foster, L.H.; Sumar, S. Selenium in health and disease: A review. Crit. Rev. Food Sci. Nutr. 1997, 37, 211–228.
45. Xu, X.-M.; Carlson, B.A.; Mix, H.; Zhang, Y.; Saira, K.; Glass, R.S.; Berry, M.J.; Gladyshev, V.N.; Hatfield, D.L. Biosynthesis of selenocysteine on its tRNA in eukaryotes. PLoS Biol. 2007, 5, 96–105.
46. Sunde, R.A.; Thompson, B.M.; Palm, M.D.; Weiss, S.L.; Thompson, K.M.; Evenson, J.K. Selenium regulation of selenium-dependent glutathione peroxidases in animals and transfected CHO cells. Biomed. Environ. Sci. 1997, 10, 346–355.
47. Johansson, L.; Gafvelin, G.; Arnér, E.S.J. Selenocysteine in proteins—Properties and biotechnological use. Biochim. Biophys. Acta Gen. Subj. 2005, 1726, 1–13.
48. Hsieh, S.H.; Ganther, H.E. Biosynthesis of dimethyl selenide from sodium selenite in rat liver and kidney cell-free systems. Biochim. Biophys. Acta Gen. Subj. 1977, 497, 205–217.
49. Ganther, H.E. Enzymic synthesis of dimethyl selenide from sodium selenite in mouse liver extracts. Biochemistry 1966, 5, 1089–1098.
50. Kim, Y.Y.; Mahan, D.C. Comparative effects of high dietary levels of organic and inorganic selenium on selenium toxicity of growing-finishing pigs. J. Anim. Sci. 2001, 79, 942–948.
51. Fortier, M.-E.; Audet, I.; Giguère, A.; Laforest, J.-P.; Bilodeau, J.-F.; Quesnel, H.; Matte, J.J. Effect of dietary organic and inorganic selenium on antioxidant status, embryo development, and reproductive performance in hyperovulatory first-parity gilts. J. Anim. Sci. 2012, 90, 231–240.
52. Bekaert, B.; Cooper, M.L.; Green, F.R.; McNulty, H.; Pentieva, K.; Scott, J.M.; Molloy, A.M.; Rayman, M.P. Effect of selenium status and supplementation with high-selenium yeast on plasma homocysteine and B vitamin concentrations in the UK elderly. Mol. Nutr. Food Res. 2008, 52, 1324–1333.
53. Davis, S.R.; Quinlivan, E.P.; Stacpoole, P.W.; Gregory, J.F., 3rd. Plasma glutathione and cystathionine concentrations are elevated but cysteine flux is unchanged by dietary vitamin B-6 restriction in young men and women. J. Nutr. 2006, 136, 373–378.
54. Lima, C.P.; Davis, S.R.; Mackey, A.D.; Scheer, J.B.;Williamson, J.; Gregory, J.F., 3rd. Vitamin B-6 deficiency suppresses the hepatic transsulfuration pathway but increases glutathione concentration in rats fed AIN-76A or AIN-93G diets. J. Nutr. 2006, 136, 2141–2147.
55. Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 5, 1957–1997.
56. Dalto, D.B.; Tsoi, S.; Audet, I.; Dyck, M.K.; Foxcroft, G.R.; Matte, J.J. Gene expression of porcine blastocysts from gilts fed organic or inorganic selenium and pyridoxine. Reproduction 2015, 149, 31–42.
57. Svoboda, M.; Ficek, R.; Drabek, J. Efficacy of selenium from Se-enriched yeast on selenium transfer from sows to piglets. Acta Vet. Brno 2008, 77, 515–521.
58. Ma, Y.L.; Lindemann, M.D.; Pierce, J.L.; Unrine, J.M.; Cromwell, G.L. Effect of inorganic or organic selenium supplementation on reproductive performance and tissue trace mineral concentrations in gravid first-parity gilts, fetuses, and nursing piglets. J. Anim. Sci. 2014, 92, 5540–5550.
59. Soda, K.; Oikawa, T.; Esaki, N. Vitamin B6 enzymes participating in selenium amino acid metabolism. BioFactors 1999, 10, 257–262.