microbial phytase in pig production
Efficacy, use and application of microbial phytase in pig production: a review
In this paper the current status of knowledge on the impact of microbial phytases in pig production is reviewed. In most studies, exogenous phytases supplemented to diets for pigs are obtained from Aspergilli. They produce 3-phytase (EC 3.1.3.8), a non-specific phosphomonoesterase, catalysing dephosphorylation of myoinositol hexakisphosphate (phytate) in a stepwise manner, producing orthophosphate, which can be absorbed by the animal.
During the last decades various experiments have been conducted all over the world with commercially available microbial phytases. Supplementation of these enzymes to diets for pigs (and poultry) results in enhancement of phosphorus (P) digestibility/availability, as assessed by different criteria.
In studies at our institute, mostly with Natuphos®, doses of microbial phytase in the range of 500 to 2000 phytase units per kg of feed resulted in generation of 0.8 to 1.0 g digestible phosphorus per kg of feed. The dose-response relationship seems to be dependent on the type of feed and the dose of microbial phytase. In addition, digestible calcium (Ca) content is increased in amounts averaging from 50 to 80% of the increase in digestible phosphorus content. Apart from the well-defined positive effects of this enzyme on calcium and phosphorus digestibility, ileal digestibility of amino acids and total tract digestibility of magnesium and trace elements (mainly zinc) are enhanced by microbial phytase. The amount and origin of phytate appear to have a substantial effect on the efficacy of microbial phytase.
Feeding frequency and amount of feed exert minor effects on the efficacy of microbial phytase. Performance (daily gain, feed conversion ratio) of pigs fed microbial phytase appears to be better as compared with a nonsupplemented diet or even with a positive control diet. Microbial phytase is now used in most pig and poultry diets in The Netherlands and can be used as a feed ingredient in linear programming of the diets. Microbial phytase supplementation to diets for pigs can substantially reduce phosphorus excretion and also to a certain extent nitrogen excretion, and can therefore reduce environmental pollution from pig production.
Introduction
Phytate is the main phosphorus-containing constituent (P content 28.2%) of many seeds and tubers; and its primary physiological role is P storage for germination (Cosgrove, 1980). In general, phytates constitute about 1-2% by weight of many cereals and oilseeds. Approximately 60-90% of total P in these seeds is present in phytate-bound form (Cheryan, 1980).
Phytates are of very limited digestibility/availability for pigs (Cromwell, 1980; Jongbloed, 1987). It therefore contributes to the P pollution in regions where land and water resources are limited and animal production is intensive such as in The Netherlands.
A possible structure of the phytate molecule (myoinositol 1,2,3,4,5,6- hexakisphosphate) when complexed with minerals and protein and starch in acidic medium is presented in Figure 1. 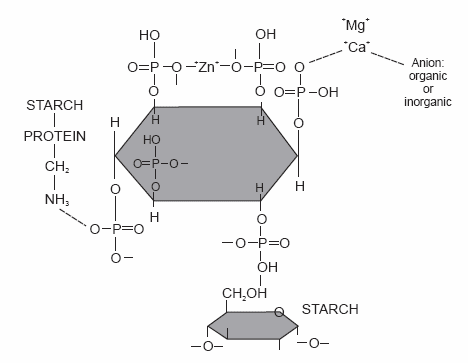
Figure 1. Phytate-protein-starch complex molecule: a potential structure (modified after Thompson, 1986; and Kies, 1998).
Phytic acid is strongly negatively charged (six reactive phosphate groups) over a wide pH range, indicating tremendous potential for complexing or binding positively charged molecules such as cations or proteins below the isoelectric point. The binding is possible within a phosphate group or between two phosphate groups on either the same or different phytic acid molecules; and the resulting structure is said to be a chelate compound (Cheryan, 1980).
Usually, phytate-bound nutrients are poorly available in the digestive tract of monogastric animals due to the lack of phytase enzyme required to cleave the phytate molecule (Cosgrove, 1980; Reddy et al., 1982; Jongbloed et al., 1993). Numerous animal experiments have documented the nutritional implications of phytates in binding dietary macro- and microelements (Ca, Mg, Fe, Zn, Cu, Mn, Mo and Co), thus reducing their solubility and bioavailability (Erdman, 1979; Torre et al., 1991). The effect of phytic acid on mineral bioavailability is influenced by pH, amount of phytic acid, mineral concentration, association/configuration of phytic acid with dietary protein/ fiber/starch, heat treatment, pre-feeding processing and the presence of other metal ions in a diet (Cheryan, 1980; Torre et al., 1991). Phytates are generally soluble at low pH, but almost completely insoluble at intestinal pH (Cheryan, 1980). The interaction between phytates and minerals leads to formation of complexes that precipitate in the duodenum (Reddy et al., 1982). Since the small intestine is the principal site of divalent cation absorption (Jongbloed et al., 1993), this implies that bioavailability of minerals can be affected by the presence of phytate. The availability of dietary minerals bound to phytate is low, especially when two or more cations are present so that a synergistic binding effect can occur (Maga, 1982).
To enable dephosphorylation of dietary phytates, there is increasing interest all over the world in supplementing such diets with purified preparations of exogenous (microbial) phytases (often obtained from genetically modified microorganisms) instead of additional phosphorus from feed phosphates.
In this paper, the effects of exogenous phytases in pig rations on animal performance and availability of P, N and some other minerals will be discussed. In addition, dietary and management factors that affect the efficacy of microbial phytase, its use in practice and the environmental impact will be considered.
Some characteristics of microbial phytases
There are several commercial microbial phytases now on the market either in dry powder or liquid forms. The phytases may differ slightly in their physical properties and enzyme activity. This may imply different efficacies of the phytase preparations. Natuphos® was the first commercially available phytase from a genetically modified Aspergillus niger strain. Since 1991 the Dutch feed industry has been using this exogenous microbial phytase, an enzymatic product obtained by fermentation of Aspergillus niger, into which the gene encoding phytase from Aspergillus niger var. Van Tieghem has been cloned using recombinant DNA technology. Therefore, it is obvious that most knowledge concerning microbial phytase has been obtained with this enzyme, although subsequently several other phytase preparations have been used. Phytase activity of this product (FTU) is assayed by measuring the amount of orthophosphate released from phytic acid within a period of linear increase with time. One unit of phytase activity is equal to 1 μmol of orthophosphate liberated from 1.5 mM Na-phytate in 1 min. at 37°C and pH 5.5 (Engelen et al., 1994). The activity of other phytase sources may be assayed under slightly different conditions.
Phytase is a phosphomonoesterase capable of hydrolyzing phytic acid (myoinositol hexakisphosphate) to yield inorganic orthophosphate that can be absorbed through the gastrointestinal wall of the pig along with a series of lower phosphoric esters of myoinositol. The dephosphorylation is necessary not only for digestibility of phytate P, but also for the utilization of minerals and proteins bound to phytates.
Intrinsic phytase present in the seeds of higher plants (e.g. wheat) is recognized by the International Union of Pure and Applied Chemistry and the International Union of Biochemistry as 6-phytase (EC 3.1.3.26). Exogenous phytase produced by microorganisms inhabiting the gastrointestinal tract such as fungi, bacteria, yeasts or intestinal endogenous phytase secreted into the lumen from the intestinal mucosa of some species is recognized as 3-phytase (EC 3.1.3.8; Wise, 1980). However, the endogenous phytase activity in the intestinal mucosa of pigs is negligible (Pointillart et al., 1984). Jongbloed et al. (1992) detected no phytase activity in the ileal digesta, which indicates that phytases from the bacterial flora up to the terminal ileum and intestinal mucosal phytases are negligible. For that reason, the ability of the pig to utilize nutrients bound to phytate complexes is primarily dependent on intrinsic phytase activity of the feed ingredients or enzymatic preparations of microbial phytase added to diets. The mode of action of phytase is illustrated in Figure 2. 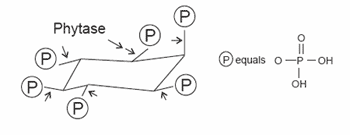
Figure 2. Schematic mode of action of microbial phytase on dietary phytates.
The enzyme cannot stand high temperatures. For instance, by pelleting a pig diet at 70°C, the initial activity is reduced by 15-25% (Schwarz and Hoppe, 1992). Microbial phytases are active over a wide range of pH, with optima at pH 2.5 and 5.5 (Simons et al., 1990). Jongbloed et al. (1992) reported from studies on cannulated pigs (ileal and duodenal cannulae) that hydrolysis of phytate in diets by microbial phytase takes place mainly in the stomach (43%), and decisively less in the small intestine (7%). This indicates that only half of the phytate was hydrolyzed by microbial phytase, which can be explained by the limited retention time in the stomach.
Effect of microbial phytase on digestibility/availability of phosphorus
Since 1990 various experiments with exogenous microbial phytases have been reported to quantify their effect on the apparent digestibility/availability of phosphorus. A survey of a large part of these studies was presented by Jongbloed et al. (1993) and Düngelhoef and Rodehutscord (1995). One of the first and most interesting experiments was the dose-response effect of microbial phytase (Natuphos®) on apparent digestibility of P in growing pigs from 20 to 55 kg (Beers and Jongbloed, 1992). Six doses of phytase (from 0 to 1800 FTU/kg) were used in two types of grower diets (based either on corn-soybean meal or phytate-rich by-products). The efficacy of microbial phytase appeared to be related to its dose and the type of diet (Figure 3).
For the corn-soybean meal diet the relation could be described by an exponential curve with the following formula:
Digestible P (g/kg) = 1.86 - 0.9963dose,
R2 = 96.7%; r.s.d. = 0.067 g digestible P/kg.
From 0 to 400 FTU/kg there was a rapid increase in microbial phytase efficacy, which flattened afterwards. For the phytate-rich by-product diet, the response could be illustrated by a logistic curve using the following
formula:
Digestible P (g/kg) = 0.95 + 1.31/(1 + e(-5.51 x 10 - 3 (dose-377.8))),
R2 = 95.5%; r.s.d. = 0.092 g digestible P/kg. 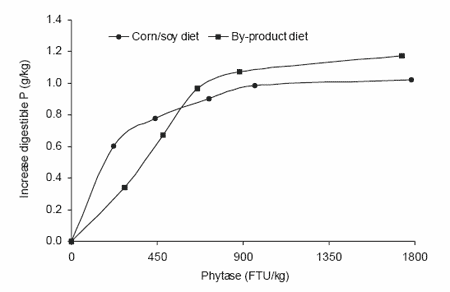
Figure 3. Improvement in digestible P by microbial phytase (Natuphos®) in two diets for growing pigs (Beers and Jongbloed, 1992).
In most cases the dose-response relationship can best be described by an exponential curve. This can also be concluded from studies of Cromwell et al. (1995) with the phytase preparation Allzyme Phytase® (Figure 4). The conclusion of the experiment of Beers and Jongbloed (1992) was that the efficacy of microbial phytase per FTU appeared to be the largest up to 500 FTU/kg diet. This dose was estimated to be equivalent to 0.8 g digestible P/ kg diet. 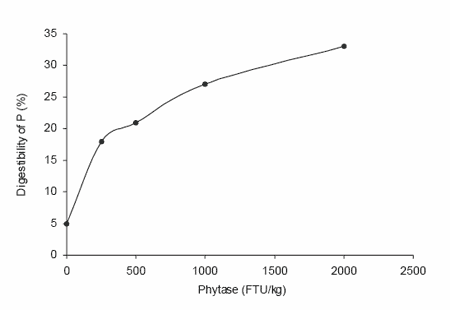
Figure 4. Improvement of availability of P (%) by microbial phytase (Allzyme Phytase®) in a corn-soybean meal diet (Cromwell et al., 1995).
Based on the curve for the corn-soybean meal based diet (Figure 3), the responses of a large number of our own experiments have also been plotted (figures not shown). This showed that responses in other experiments in our laboratory fit quite well with the curve for corn-soybean meal. In all experiments it is shown that microbial phytase is considerably effective in enhancing P digestibility and so increases the amount of digestible/available P in the feed for pigs.
The efficacy of microbial phytase also depends on animal related factors such as physiological status and housing conditions (Kemme et al., 1997a). Kemme et al. (1997b) showed that the efficacy of phytase in generating digestible P decreased in the order of lactating sows, growing-finishing pigs, sows at the end of pregnancy, piglets and sows at midpregnancy.
Dietary factors affecting efficacy of microbial phytase
A prerequisite for a good evaluation of microbial phytase efficacy is that the animals be fed below their P requirement. This is due to intestinal regulation of P absorption when animals are fed above their P requirement.
There is little information available on dietary factors affecting efficacy of microbial phytase. It is commonly known that higher dietary Ca levels decrease apparent absorption of P (Jongbloed, 1987). However in most experiments dietary treatments with different Ca levels were implemented only in diets with microbial phytase and not simultaneously in diets without microbial phytase (Lei et al., 1994; Lantzsch et al., 1995). Mostly a linear decrease of P absorption is observed. In two experiments we studied whether a possible interaction could be demonstrated between dietary Ca level and microbial phytase supplementation on P absorption (Mroz et al., 1994b; Jongbloed et al., 1995). We observed a linear decrease in apparent digestibility of P at higher Ca levels, but no interaction with microbial phytase could be demonstrated.
Another dietary factor is the amount and source of phytate. In two experiments we studied the effect of phytate amount and source on the efficacy of microbial phytase (Dekker et al., 1992). For this purpose we used diets based on either corn or sunflower seed meal at two inclusion levels (and so two levels of phytate P). The concentrations of phytate P were 1.2 and 1.8 g/kg; and phytase activities in the corn and sunflower-based diets were 450 and 340 FTU/kg, respectively. We observed that in the cornbased diet the higher level of phytate generated substantially more digestible P, while in the sunflower-based diet slightly more digestible P was generated at the higher phytate level. It was concluded firstly that in both diets the lower level of phytate was too low to get maximal effect of the enzyme; and secondly that phytate in corn is more readily available than phytate in sunflower seed meal. Lack of substrate (phytate) may occur in piglet diets formulated with large proportions of animal products as a protein or P source or with diets that already contain a high oncentration of intrinsic phytase like wheat, wheat bran, barley, rye or triticale (Düngelhoef et al., 1994; Eeckhout and De Paepe, 1992).
Jongbloed et al. (2000) performed two experiments in which diets were supplemented with or without microbial phytase and/or lactic acid and formic acid. Supplementary lactic acid and formic acid exerted a synergistic effect on apparent digestibility of P. This means that efficacy of microbial phytase could be further enhanced by some organic acids.
Effect of phytase on calcium digestibility
As stated earlier, phytate complexes may be formed with various di- and trivalent cations (Wise, 1980). Microbial phytase supplementation also increases Ca digestibility. To quantify this effect the ratio was calculated between the increase in amount of digestible Ca and the amount of digestible P. All available data from our own experiments and those in the literature were used (Figure 5). The ratios in our own experiments (n=12) were on average 0.55±0.19, while those from data in the literature (n=20) were significantly higher 0.84±0.33. We have no clear explanation for this difference, but the levels of dietary Ca and digestible P certainly play a part (Mroz et al., 1994b). In practical diet formulation the higher Ca digestibility means that supplementation of Ca in diets with microbial phytase can be slightly reduced, which may be beneficial as this lowers buffering capacity of the diet (less digestive disorders). Using 500 FTU/kg of feed generates 0.8 g/kg of digestible P and 0.4 to 0.7 g/kg of digestible Ca. Assuming a Ca digestibility of supplemental limestone of 60%, then etween 1.7 and 3.0 g less limestone per kg of feed need be supplied. 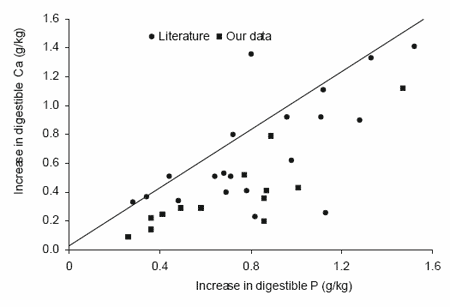
Figure 5. Effect of microbial phytase on the improvement of both digestible Ca and digestible P in the same diet.
Apart from its effect on P and Ca digestibility, phytase affects the availability of other minerals and nutrients. Microbial phytase can improve the apparent absorption of Mg, Na, K, Zn, Cu and Fe (Jongbloed et al., 1999).
Effect of phytase on digestion and/or utilization of dietary protein, amino acids and energy
Some in vitro and in vivo studies on rats have documented that dietary protein and amino acid digestibilities may be reduced by phytates due to ability to form insoluble complexes under gastrointestinal conditions and to inhibit some endogenous proteolytic enzymes (Atwal et al., 1980; Knuckles, 1985; Ritter et al., 1987). Based on these studies, it is assumed that dietary phytases (intrinsic and extrinsic) may prevent the negative effects of phytates on protein and starch digestibilities by hydrolysing the phytate-protein-starch complexes and returning dietary protein, amino acids and starch to soluble and absorbable form in the small intestine of the pig.
APPARENT TOTAL TRACT DIGESTIBILITY AND UTILIZATION OF PROTEIN BY PIGS FED EXOGENOUS PHYTASE
From a nutritional point of view, total tract digestibility of dietary protein and amino acids is not an accurate measure of nutritive value. However, in practice it may be important to determine the environmental burden of itrogen in response to various doses of phytase. Data on total tract digestibility of dry matter, organic matter and crude protein in relation to microbial phytase addition as extracted from the publications of Mroz et al. (1994a) and Näsi et al. (1995) are presented in Table 1. Data from studies of Mroz et al. (1994a) indicate that microbial phytase (Natuphos®) significantly increased apparent total tract digestibility of crude protein by 2.3 ercentage units. In addition, nitrogen retention tended to be higher though the difference was not statistically significant. In contrast, Näsi et al. (1995) showed only a marginal improvement in crude protein total tract digestibility whereas N retention was minimally negative.
Table 1. Apparent total tract digestibility of dry matter (%), organic matter and crude protein and N retention (as % of intake) in pigs fed diets without or with microbial phytase. 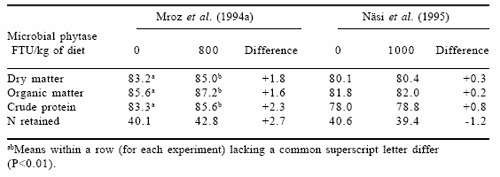
In another study, Khan and Cole (1993) fed a barley-based diet to six cannulated gilts without or with Aspergillus niger phytase (1000 FTU/kg). The authors found that total tract crude protein digestibility for the control and microbial phytase groups was 81.4 and 85.5% (SED=1.7), respectively. Total tract digestibility of gross energy increased also by 1.2 percentage units in the presence of microbial phytase, but this difference was not statistically significant.
Ketaren et al. (1993) fed 60 gilts (initial weight 20 kg) a soybean mealsucrose- based diet without and with microbial phytase (1000 FTU/kg). After 35 days the gilts were slaughtered to assess the deposition of protein, fat and energy (Table 2). The authors observed that microbial phytase (1000 FTU/kg) significantly increased protein deposition (+15 g/d), protein retained:protein intake ratio (+0.031 kg/kg), energy retained:DE intake (+0.02 MJ/MJ) and protein retained:DE intake (+0.4 g/MJ).
Table 2. Retention of protein, fat and energy in growing pigs given a soybeansucrose- based diet with and without microbial phytase (1000 FTU/kg)1. 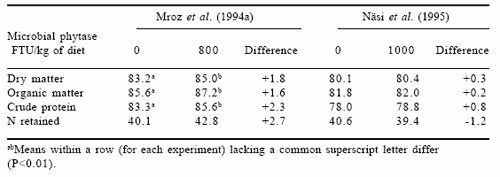
Han et al. (1997) used 48 pigs over 10 to 90 kg to study the effects of supplemental microbial phytase (1000-1200 FTU/kg), cereal phytase (230- 460 FTU/kg) and inorganic P (0.20-0.24%) on apparent total tract digestibility and retention of N. Microbial phytase in this study resulted in a higher crude protein digestibility (+3.0 percentage units), but this difference was not statistically significant from the control group. Biehl and Baker (1996) concluded from studies with 15 young pigs (9.1 kg initial weight) that phytase supplementation (1200 FTU/kg) improved amino acid utilization in amino acid-deficient corn-soybean meal diets. Kemme and Jongbloed (1994) reported that both Aspergillus niger phytase and intrinsic phytase from wheat middlings exerted no effect on crude protein digestibility in diets for piglets, sows or growing-finishing pigs. In line with these findings were the conclusions of Näsi (1990), Näsi and Helander (1994), Pallauf et al. (1994), Bruce and Sundstøl (1995), Kornegay et al. (1996) and Valaja et al. (1998).
We calculated the effect of microbial phytase on apparent fecal digestibility of some nutrients. In our studies microbial phytase enhanced average dry matter digestibility by 0.54 ±0.81 percentage units (n=18), organic matter by 0.33 ±0.72 (n=9) and N by 1.21±1.73 (n=7). From these data it can be concluded that there is a tendency toward increased fecal digestibilities of dry matter, organic matter and N in response to supplementary microbial phytase. Data from the literature showed a slightly different picture. Fecal digestibilities of dry matter and organic matter were slightly reduced by 0.11±0.69 (n=9) and 0.37±0.57 (n=3), respectively, while digestibility of N was increased by 0.60±1.66 (n=10) by supplementary phytase.
APPARENT ILEAL DIGESTIBILITY
There is a limited number of reports on the apparent ileal digestibility of crude protein in pigs in relation to phytase supplementation. Ileal digestibility of N and amino acids is usually considered a proper criterion of dietary protein value (Low, 1989). Officer and Batterham (1992) reported that ileal digestibility of crude protein and essential amino acids increased by 7-12 percentage units. Also, Khan and Cole (1993) observed an increase in ileal nitrogen digestibility of 12.8 percentage units (P<0.077) when feeding a high-phytate barley-based diet with Aspergillus niger phytase (1000 FTU/ kg) to six cannulated gilts. Mroz et al. (1994a) observed a lower ffect (+3.5 percentage units); and in a second experiment Kemme et al. (1999) showed a significantly higher ileal digestibility for several amino acids (Table 3). However, there are a few reports showing that microbial phytase exerted no effect on apparent ileal digestibility of crude protein or amino acids (Lantzsch et al., 1995; Valaja et al., 1998).
Table 3. Effect of microbial phytase on apparent ileal digestibility of N and some amino acids. 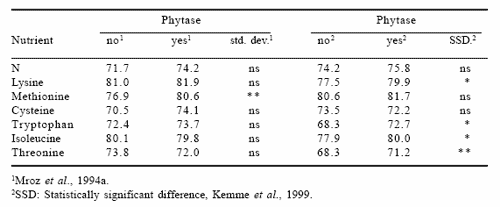
Effect of management factors on efficacy of microbial phytase
In practice, pigs are raised under a wide variety of housing and feeding conditions. Therefore, it is important to know the effects of different feeding regimens and housing conditions on the efficacy of microbial phytase. In one study we could not show any effect of feeding level (2.3 and 2.8 times maintenance requirement for metabolizable energy) on the efficacy of microbial phytase (Mroz et al., 1994a). In the same experiment no effect on phytase efficacy was noted between feeding two and seven times a day, while efficacy was slightly decreased when pigs were fed once daily.
The efficacy of microbial phytase (500 FTU/kg) is increased by soaking a phytate-rich diet (maize, tapioca, beans, phytase-inactivated wheat bran and extracted sunflower meal) for 8-15 hrs prior to feeding (Kemme and Jongbloed, 1993). Some results of this treatment in a trial with growing pigs from 30 to 70 kg body weight are presented in Table 4. Soaking a barleysoybean meal diet supplemented with microbial phytase for 3 hrs, however, did not result in a higher P digestibility (Näsi and Helander, 1994). This may have been due to the short soaking time and the rather high dietary Ca content. In a second experiment of Näsi et al. (1995), soaking a barleyrapeseed meal diet for 3 hrs with dried whey at 40°C increased P digestibility by 4%.
Table 4. Effects of soaking a phytate-rich diet without or with microbial phytase on the apparent digestibility of Ca and total P1. 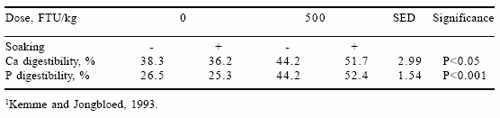
Effect of microbial phytase on pig performance
Jongbloed et al. (1999) presented results on the effect of microbial phytase on performance of pigs compared, where possible, with both negative control and positive control treatments. However, the choice of a positive control treatment was not always evident. Therefore, they chose for the positive control a treatment with about 1 g of supplementary P from a feed phosphate. However, in half of the experiments the difference with the negative control treatment was often between 2 and 3 g of P. For their review, they chose 11 experiments where a reasonable comparison was possible. Relative values were calculated based on a value for the negative control diet of 100 (Table 5). Data in Table 5 show that the performance of both the positive control and the phytase groups were superior to the negative control group. The positive control group and the phytase supplemented group were almost identical. From the eleven experiments there were six in which the positive control group was also supplemented with microbial phytase. Performance of the pigs receiving the positive control group with supplementary phytase was slightly better than those without phytase. The relative ratios compared with the positive control diet for growth rate, feed intake and feed conversion ratio were 106.0±5.5, 103.0±3.2 and 95.7±4.9, respectively. This may imply that either the P requirement was not yet met, which is unlikely, or there is another positive effect of microbial phytase on performance.
Further calculations were performed on the effect of Natuphos® on the performance of piglets fed above their P requirement (Kies, 1997). Feed conversion ratio (corrected for differences in weight gain using 25 g equal to 0.01 unit FCR) was taken as the response variable. In total, 11 experiments comprising 17 trials met this criteria. The experiments were analysed using the model:
Table 5. Relative performance of pigs using the negative control diet as 100 (n=11) and effect of phytase when added to the positive control diet (n=6). 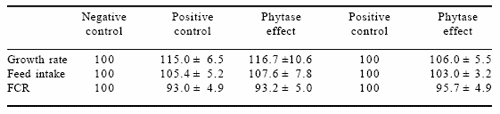
Corrected FCR = Experimenti + b*FTU/kg added
Experimenti (i=1.17) is a constant per experiment and b is the regression coefficient. The mean corrected FCR was 1.483. The value for b was -0.000094, with a 95% confidence interval of -0.000051 to -0.000136; R2 =0.94.
It can be derived from the equation that dietary supplementation of 500 FTU Natuphos®/kg improved the corrected FCR by 0.047 units, which is close to the value reported by Jongbloed et al. (1999). Due to lack of sufficient data he could only use a linear model, while it may be speculated that an exponential curve might be more physiologically appropriate. From the experiments of Campbell et al. (1995) and Selle et al. (1996) it an also be concluded that dietary level of phytate influences the improvement in FCR. The improvement in FCR can be attributed to a higher digestibility of protein/amino acids and a slightly higher energy digestibility. Also, other unknown factors may play a part.
Environmental impact of microbial phytase
In the absence of microbial phytase only ~16% of P in corn and ~38% of P in soybean meal is digested by pigs. Because of the large amount of undigested dietary P, a substantial amount of P is excreted via feces. Based on the estimates of Cromwell et al. (1993), a dose of microbial phytase equal to 1000 FTU/g converted approximately one-third of the unavailable P to an available form. About 500 FTU/kg of diet generates approximately 0.8 g digestible P/kg, which is equivalent to 1.0 g P from monocalcium phosphate or 1.23 g P from dicalcium phosphate, which is often used in the United States. This is illustrated in Table 6. This table shows that with supplementation of 500 FTU/kg of feed, total P content is 1.3 g/kg lower.
With the same performance as the control diet between 20 and 50 kg live weight and a feed conversion ratio of 2.5, it can be calculated that P excretion per kg growth is 4.75 instead of 8.0 g, which is 40% lower.
Table 6. Effect of microbial phytase (500 FTU/kg feed) on diet formulation and mineral content in a grower diet (g/kg). 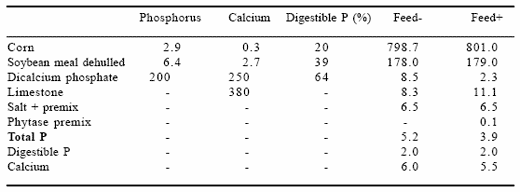
In The Netherlands, legislation has been imposed to allow a maximum amount of P to be spread per hectare of land. When this amount has been exceeded, the farmer must pay a levy and has to transport the manure elsewhere (Jongbloed and Lenis, 1998). This has stimulated the replacement of inorganic feed phosphates by microbial phytase in pig and poultry diets.
Microbial phytase is considered as a raw material containing a high phosphorus value and can be used in linear programming for feed formulation. Therefore, in most cases the feed compounder chooses a certain amount of phytase units, which is linked to the amount of digestible P generated. In general, between 300 and 500 FTU/kg is added. Beyond that, inorganic feed phosphate can be added to meet the P requirement as necessary. Microbial phytase can be used as a dry or liquid raw material. The latter is increasingly used, because the phytase is not completely resistant to high steam-pelleting temperatures (exceeding 80°C). Much effort has gone into developing a more heat-resistant phytase and to choosing formulations in which phytase can better resist high temperatures. Microbial phytase is most effective in layer diets, and is therefore highly competitive to inorganic feed phosphates for this category. In general, microbial phytase is incorporated in most pig and poultry diets in The Netherlands, but is becoming increasingly popular in other countries for reasons beyond its positive effect on the reduction of P excretion. This is predominantly due to associated enhanced effects on the digestibility of protein and amino acids, energy, calcium and zinc, and last but not least on animal performance.
In The Netherlands, the concentrations of several minerals in animal diets are occasionally monitored. Therefore, it is possible to estimate the excretion of P by several categories of pigs. In Table 7 a survey is presented for growing-finishing pigs from 1973 to 1998. In that period performance increased considerably. Growth rate increased from 625 to 755 g/day, while feed conversion ratio improved from 3.37 to 2.69. Despite the better performance, P content in pig diets decreased by more than 2.0 g/kg. From 1988 onwards it has been common to feed a starter diet and a grower-finisher diet. Data in Table 7 show that the excretion of P in growingfinishing pigs has reduced by more than half in 20 years. This lower P excretion has, apart from increased nutritional knowledge, undoubtedly been stimulated by legislation based on P. As microbial phytase has been commercially available in The Netherlands since late 1991, the decrease in P excretion since then is predominantly due to microbial phytase. Incorporation levels of microbial phytase will undoubtedly increase in the coming years resulting in a further decrease of P excretion by pigs.
Table 7. Mean excretion of P of a growing-finishing pig from 25 to 110 kg in The Netherlands. 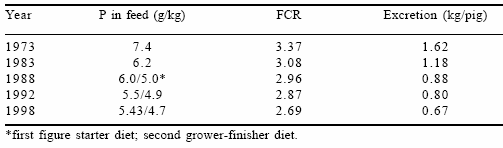
References
Atwal, A.S., N.A.M. Eskin, B.E. McDonald and M. Vaisey-Genser. 1980. The effects of phytate on nitrogen utilization and zinc metabolism in young rats. Nutr. Rep. Int. 21(2)257-267.
Beers, S. and A.W. Jongbloed. 1992. Apparent overall (total tract) digestibility of P in relation to doses of Aspergillus niger phytase in diets for pigs. J. Anim. Sci. 70(Suppl. 1):242.
Biehl, R.R. and D.H. Baker. 1996. Efficacy of supplemental 1"- hydroxycholecalciferol and microbial phytase for young pigs fed phosphorus- or amino acid-deficient corn-soybean meal diets. J. Anim. Sci. 74:2960-2966.
Bruce, J.A.M. and F. Sundstøl. 1995. The effect of microbial phytase in diets for pigs on apparent ileal and faecal digestibility, pH and flow of digesta measurements in growing pigs fed a high-fibre diet. Can. J. Anim. Sci. 75:121-127.
Campbell, R.G., D.T. Harrison, K.J. Butler and P.H. Selle. 1995. Effects of dietary available phosphorus and phytase (Natuphos) on the performance of pigs from 19 to 40 days post-weaning. In: Manipulating pig production V (D.P. Hennessy and P.D. Cranwell, eds), Proc. of the Fifth Biennial Conf. of the Australasian Pig Science Association, Werribee, Victoria, Australia, p. 193.
Cheryan, M. 1980. Phytic acid interactions in food systems. CRC Crit. Rev. Food Sci. 13: 297-335.
Cromwell, G.L. 1980. Biological availability of phosphorus in feedstuffs for swine. Feedstuffs 52:(9)14-16.
Cromwell, G.L., T.S. Stahly, R.D. Coffey, H.J. Monegue and J.H. Randolph. 1995. Efficacy of phytase in improving the bioavailability of phosphorus in soybean meal and corn-soybean meal diets for pigs. J. Anim. Sci. 71:1831-1840.
Cromwell, G.L., R.D. Coffey, H.J. Monegue and J.H. Randolph. 1993. Efficacy of low-activity phytase in improving the bioavailability of phosphorus in corn-soybean meal diets for pigs. J. Anim. Sci. 73:449-456.
Dekker, R.A., P.A. Kemme and A.W. Jongbloed. 1992. Methodological comparison of the assessment of P digestibility of tapioca and maize, and the influence of amount and origin of phytic acid on the efficacy of microbial phytase from Aspergillus niger. Report IVVO-DLO, Lelystad, nr. 244.
Düngelhoef, M., M. Rodehutscord, H. Spiekers and E. Pfeffer. 1994. Effects of supplemental microbial phytase on availability of phosphorus contained in maize, wheat and triticale to pigs. Anim. Feed Sci. Technol. 49:1-10.
Düngelhoef, M. and M. Rodehutscord. 1995. Effects of phytases on the digestibility of phosphorus in pigs. Übers. Tierernährg. 23:133-157.
Eeckhout, W. and M. De Paepe. 1992. Betere benutting van de voeders. 1.1. Synthetische fytase. 1.1.2. Tarwefytase en een microbieel fytasepreparaat en de schijnbare fosforverteerbaarheid van een eenvoudig biggenvoeder. Landbouwtijdschrift-Revue de l’Agriculture 45:193-204. Engelen, A.J., F.C. van der Heeft, H.G. Randsdorp and E.L.C. Smit. 1994. Simple and rapid determination of phytase activity. J. AOAC Intern. 77:760-764.
Erdman, J.W. 1979. Oilseed phytates: nutritional implications. J. Am. Oil Chem. Soc. 56:736-741.
Han, Y.M., F. Yang, A.G. Zhou, E.R. Miller, P.K. Ku, M.G. Hogberg and X.G. Lei. 1997. Supplemental phytases of microbial and cereal sources improve dietary phytate phosphorus utilization by pigs from weaning through finishing. J. Anim. Sci. 75:1017-1025.
Jongbloed, A.W. 1987. Phosphorus in the feeding of pigs. Effect of diet on absorption and retention of phosphorus by growing pigs. PhD thesis, Report IVVO-DLO nr. 179, Lelystad, The Netherlands, 343 pp.
Jongbloed, A.W. and N.P. Lenis. 1998. Environmental concerns about animal manure. J. Anim. Sci. 76:2641-2648.
Jongbloed, A.W. and Z. Mroz. 1999. Influence of phytase on availability of phosphorus, amino acids and energy in swine. In: Proc. BASF Technical
Symposium. Use of Natuphos® Phytase in Swine and waste Management Midwest Series 1999, 1-20.
Jongbloed, A.W., Z. Mroz and P.A. Kemme. 1992. The effect of supplementary Aspergillus niger phytase in diets for pigs on concentration and
Cosgrove, D.J. 1980. Inositol phosphates. Their chemistry, biochemistry and physiology. Elsevier Science Publishers, Amsterdam, p.175.
Cromwell, G.L. 1980. Biological availability of phosphorus in feedstuffs for swine. Feedstuffs 52:(9)14-16.
Cromwell, G.L., T.S. Stahly, R.D. Coffey, H.J. Monegue and J.H. Randolph. 1995. Efficacy of phytase in improving the bioavailability of phosphorus in soybean meal and corn-soybean meal diets for pigs. J. Anim. Sci. 71:1831-1840.
Cromwell, G.L., R.D. Coffey, H.J. Monegue and J.H. Randolph. 1993. Efficacy of low-activity phytase in improving the bioavailability of phosphorus in corn-soybean meal diets for pigs. J. Anim. Sci. 73:449-456.
Dekker, R.A., P.A. Kemme and A.W. Jongbloed. 1992. Methodological comparison of the assessment of P digestibility of tapioca and maize, and the influence of amount and origin of phytic acid on the efficacy of
microbial phytase from Aspergillus niger. Report IVVO-DLO, Lelystad, nr. 244.
Düngelhoef, M., M. Rodehutscord, H. Spiekers and E. Pfeffer. 1994. Effects of supplemental microbial phytase on availability of phosphorus contained in maize, wheat and triticale to pigs. Anim. Feed Sci. Technol. 49:1-10.
Düngelhoef, M. and M. Rodehutscord. 1995. Effects of phytases on the digestibility of phosphorus in pigs. Übers. Tierernährg. 23:133-157. Eeckhout, W. and M. De Paepe. 1992. Betere benutting van de voeders. 1.1. Synthetische fytase. 1.1.2. Tarwefytase en een microbieel fytasepreparaat en de schijnbare fosforverteerbaarheid van een eenvoudig biggenvoeder. Landbouwtijdschrift-Revue de l’Agriculture 45:193-204.
Engelen, A.J., F.C. van der Heeft, H.G. Randsdorp and E.L.C. Smit. 1994. Simple and rapid determination of phytase activity. J. AOAC Intern. 77:760-764.
Erdman, J.W. 1979. Oilseed phytates: nutritional implications. J. Am. Oil Chem. Soc. 56:736-741.
Han, Y.M., F. Yang, A.G. Zhou, E.R. Miller, P.K. Ku, M.G. Hogberg and X.G. Lei. 1997. Supplemental phytases of microbial and cereal sources improve dietary phytate phosphorus utilization by pigs from weaning through finishing. J. Anim. Sci. 75:1017-1025.
Jongbloed, A.W. 1987. Phosphorus in the feeding of pigs. Effect of diet on absorption and retention of phosphorus by growing pigs. PhD thesis, Report IVVO-DLO nr. 179, Lelystad, The Netherlands, 343 pp.
Jongbloed, A.W. and N.P. Lenis. 1998. Environmental concerns about animal manure. J. Anim. Sci. 76:2641-2648.
Jongbloed, A.W. and Z. Mroz. 1999. Influence of phytase on availability of phosphorus, amino acids and energy in swine. In: Proc. BASF Technical Symposium. Use of Natuphos® Phytase in Swine and waste Management Midwest Series 1999, 1-20.
Jongbloed, A.W., Z. Mroz and P.A. Kemme. 1992. The effect of supplementary Aspergillus niger phytase in diets for pigs on concentration and apparent digestibility of dry matter, total phosphorus, and phytic acid in
different sections of the alimentary tract. J. Anim. Sci. 70:1159-1168.
Jongbloed, A.W., P.A. Kemme and Z. Mroz. 1993. The role of microbial phytases in pig production, In: Enzymes in Animal Production (C. Wenk and M. Boessinger, eds), Proceedings of the 1st Symposium, Kartause Ittingen, Switzerland, p. 173-180.
Jongbloed, A.W., P.A. Kemme, Z. Mroz and R. ten Bruggencate. 1995. Apparent total tract digestibility of organic matter, N, Ca, Mg and P in growing pigs as affected by levels of Ca, microbial phytase and phytate.
In: W. van Hartingsveldt, M. Hessing, J.P. van der Lugt and W.A.C. Somers (Eds). Proceedings of Second Symposium on Feed Enzymes (ESFE2), Noordwijkerhout, Netherlands, TNO Nutrition and Food Research Institute, Zeist, p. 198-204.
Jongbloed, A.W., R. van der Weij-Jongbloed, P.A. Kemme and Z. Mroz. 2000. The effects of microbial phytase, organic acids and their interaction in diets for growing pigs. Livestock. Prod. Sci. (in press).
Kemme, P.A. and A.W. Jongbloed. 1993. Effect of Aspergillus niger phytase and soaking on the digestibility of P in diets for pigs. J. Anim. Sci. 71: (Suppl. 1):181.
Kemme, P.A. and A.W. Jongbloed. 1994. Effekt van plantaardig en Aspergillus niger fytase, leeftijd en voerniveau op de verteerbaarheid van Weende analyse komponenten, Ca en P bij biggen [Effect of intrinsic and Aspergillus niger phytase, age and feeding level on digestibilities of proximate components, Ca en P in diets for piglets]. Report IVVO-DLO no. 257.
Kemme, P.A., J.S. Radcliffe, A.W. Jongbloed and Z. Mroz. 1997a. The effects of body weight, housing, and calculation method on mineral digestibility and the efficacy of microbial phytase in diets for growingfinishing pigs. J. Anim. Sci. 75:2139-2146.
Kemme, P.A., A.W. Jongbloed, Z. Mroz and A.C. Beynen. 1997b. The efficacy of Aspergillus niger phytase in rendering phytate phosphorus available for absorption in pigs is influenced by their physiological status.
J. Anim. Sci. 75:2129-2138.
Kemme, P.A., A.W. Jongbloed, Z. Mroz, J. Kogut and A.C. Beynen. 1999. Digestibility of nutrients in growing-finishing pigs is affected by Aspergillus niger phytase, phytate and lactic acid levels. 1. Apparent ileal digestibility of amino acids. Livest. Prod. Sci. 58:107-117.
Ketaren, P.P., E.S. Batterham, E.B. Dettmann and D.J. Farrel. 1993. Phosphorus studies in pigs. 3. Effect of phytase supplementation on the digestibility and availability of phosphorus in soyabean meal for grower pigs. Br. J. Nutr. 70:289-311.
Khan, N. and D.J.A. Cole. 1993. The effect of dietary inclusions of phytase and yeast on apparent phosphorus digestibility in pigs. Proc. of the Winter Meeting of the British Society of Animal Production. Scarborough, England. p. 2.
Kies, A. 1997. Niet-fosfor effecten van Natuphos®. In: BASF Symposium“Enzymen en organische zuren in de Nederlandse mengvoederindustrie”, Arnhem, The Netherlands. p. 11.
Kies, A. 1998. The influence of Natuphos® phytase on the bioavailability of protein in swine. In: BASF Technical Symposium, World Pork Expo, De Moines, IA. p. 1-12.
Knuckles, B.E., D.D. Kuzmicky and A.A. Betschart. 1985. Effect of phytate and partially hydrolyzed phytate on in vitro protein digestibility. J. Food Sci. 50:1080-1082.
Kornegay, E.T., D.M. Denbow, Z. Yi and V. Ravindran. 1996. Response of broilers to graded levels of Natuphos® phytase added to corn-soybean meal based diets containing three levels of nonphytate phosphorus. Br. J. Nutr. 75:839-852.
Lantzsch, H.J., S. Wjst, and W. Drochner. 1995. The effect of dietary calcium on the efficacy of microbial phytase in rations for growing pigs. J. Anim. Physiol. and Anim. Nutr. 73:19-26.
Lei, X.G., P.K. Ku, E.R. Miller, M.T. Yokoyama and D.E. Ullrey. 1994. Calcium level affects the efficacy of supplemental microbial phytase in corn-soybean meal diets of weanling pigs. J. Anim. Sci. 72:139-143.
Low, A.G. 1989. Research into the digestive physiology of pigs. In: Nutrition and digestive physiology in monogastric farm animals (E.J. van Weerden and J. Huisman, eds.). Wageningen, The Netherlands. p. 1-12.
Maga, J.A. 1982. Phytate: Its chemistry, occurrence, food interactions, nutritional significance, and methods of analysis. J. Agric. Food Chem. 30:1-8.
Mroz, Z., A.W. Jongbloed and P.A. Kemme. 1994a. Apparent digestibility and retention of dietary nutrients bound to phytase complexes as influenced by microbial phytase and feeding regimen in pigs. J. Anim. Sci.
72:126-132.
Mroz, Z., A.W. Jongbloed and P.A. Kemme. 1994b. The influence of graded calcium supply on microbial phytase efficacy in starter diets for pigs. In: 45th Annual Meeting of the EAAP, Edinburgh; paper P4.3.
Näsi, M. 1990. Microbial phytase supplementation for improving availability of plant phosphorus in the diet of the growing pigs. J. Agric. Sci. in Finland 62:435-443.
Näsi, M. and E.H. Helander. 1994. Effects of microbial phytase supplementation and soaking of barley-soybean meal on availability of plant phosphorus for growing pigs. Acta Agric. Scand., Sect. A, Animal Sci. 44:79-86.
Näsi, M., E.H. Helander and K.H. Partanen. 1995. Availability for growing pigs of minerals and protein of a high phytate barley-rapeseed meal diet treated with Aspergillus niger phytase or soaked with whey. Animal Feed Sci. Technol. 56:83-98.
Officer, D.I. and E.S. Batterham. 1992. Enzyme supplementation of LinolaTM meal. In Proc. Wollongbar Pig Industry Seminar on Feed Enzymes, p. 56- 57.
Pallauf, J., G. Rimbach, S. Pippig, B. Schindler and E. Most. 1994. Effect of phytase supplementation to a phytate-rich diet based on wheat, barley and soya on the bioavailability of dietary phosphorus, calcium, magnesium, zinc and protein in piglets. Agribiol. Res. 47:39-48.
Pointillart, A., N. Fontaine and M. Thomasset. 1984. Phytate phosphorus utilization, intestinal phosphatases in pigs fed low phosphorus: wheat or corn diets. Nutr. Report Int. 29: 473-483.
Reddy, N.R., S.K. Sathe and D.K. Salunkhe. 1982. Phytates in legumes and cereals. Adv. Food Res. 28:1-92.
Ritter, M.A., C.V. Morr and R.L. Thomas. 1987. In vitro digestibility of phytate-reduced and phenolic-reduced soy protein isolates. J. Food Sci. 52:325-327.
Schwarz, G. and P.P. Hoppe. 1992. Phytase enzyme to curb pollution from pigs and poultry. Feed Magazine 1:22-26.
Selle, P.H., V. Ravindran, D.J. Cadogan, A.R. Walker and W.L. Bryden. 1996. The role of microbial phytases in poultry and pig production. Proc. Tenth Australian Poultry and Feed Convention, Melbourne, Australia, p. 219-224.
Simons, P.C., H.A.J. Versteegh, A.W. Jongbloed, P.A. Kemme, P. Slump, K.D. Bos, M.G.E. Wolters, R.F. Beudeker and G.J. Verschoor. 1990. Improvement of phosphorus availability by microbial phytase in broilers and pigs. Brit. J. Nutr. 64:525-540.
Thompson, L.U. 1986. In: Phytic acid chemistry and applications (E. Graf, ed.), Pilatus Press, Minneapolis. p 173-194.
Torre, M., A.R. Rodriguez and F. Saura-Calixto. 1991. Effects of dietary fiber and phytic acid on mineral availability. Crit. Rev. Food Sci. Nutr. 1(1):1-22.
Valaja, J., S. Plaami and H. Siljander-Rasi. 1998. Effect of microbial phytase on digestibility and utilisation of phosphorus and protein in pigs fed wet barley protein with fibre. Animal Feed Sci. Technol. 72:221-233.
Wise, A. 1980. Dietary factors determining the biological activities of phytate. Nutr. Abstr. Rev. 53:791-806.






.jpg&w=3840&q=75)