Grape Seed Waste Counteracts Aflatoxin B1 Toxicity in Piglet Mesenteric Lymph Nodes
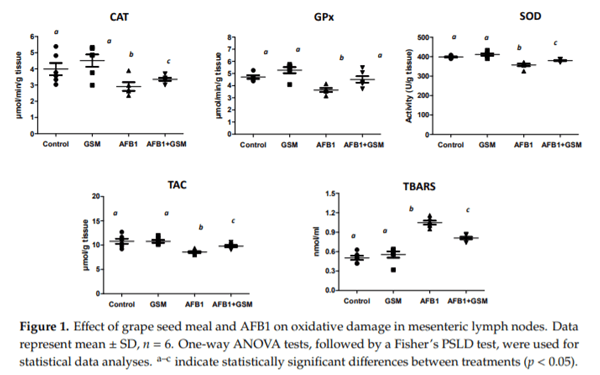
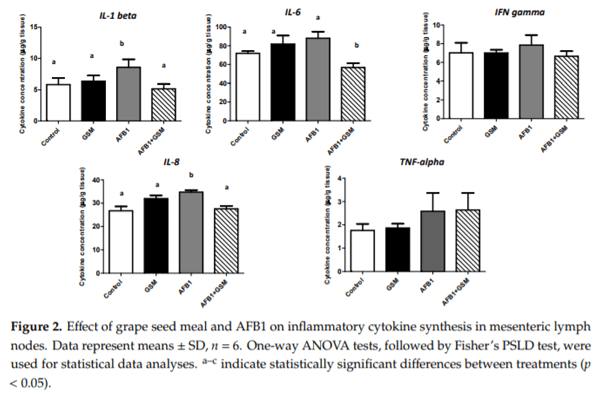
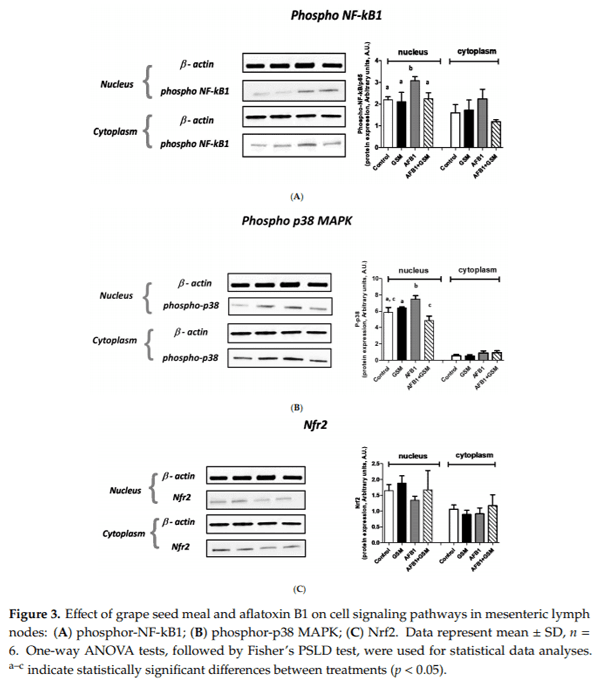
Aflatoxin B1 (AFB1) is a mycotoxin that frequently contaminates cereals and cereal byproducts. This study investigates the effect of AFB1 on the mesenteric lymph nodes (MLNs) of piglets and evaluates if a diet containing grape seed meal (GSM) can counteract the negative effect of AFB1 on inflammation and oxidative stress. Twenty-four weaned piglets were fed the following diets: Control, AFB1 group (320 μg AFB1/kg feed), GSM group (8% GSM), and AFB1 + GSM group (8% GSM + 320 μg AFB1/kg feed) for 30 days. AFB1 has an important antioxidative effect by decreasing the activity of catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPx) and total antioxidant status. As a result of the exposure to AFB1, an increase of MAP kinases, metalloproteinases, and cytokines, as effectors of an inflammatory response, were observed in the MLNs of intoxicated piglets. GSM induced a reduction of AFB1-induced oxidative stress by increasing the activity of GPx and SOD and by decreasing lipid peroxidation. GSM decreased the inflammatory markers increased by AFB1. These results represent an important and promising way to valorize this waste, which is rich in bioactive compounds, for decreasing AFB1 toxic effects in mesenteric lymph nodes.
Keywords: aflatoxin B1; grape seed meal; mesenteric lymph nodes; piglets
Key Contribution: Aflatoxin B1 induces inflammation and oxidative stress in mesenteric lymph nodes. In the mesenteric lymph nodes of AFB1-intoxicated piglets, grape seed meal can improve some markers of inflammation and oxidative stress.
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [CrossRef]
- Sarma, U.P.; Bhetaria, P.J.; Devi, P.; Varma, A. Aflatoxins: Implications on health. Indian J. Clin. Biochem. 2017, 32, 124–133. [CrossRef] [PubMed]
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin b1 and m1: Biological properties and their involvement in cancer development. Toxins 2018, 10, 214. [CrossRef] [PubMed]
- Taniwaki, M.H.; Pitt, J.I.; Magan, N. Aspergillus species and mycotoxins: Occurrence and importance in major food commodities. Curr. Opin. Food Sci. 2018, 23, 38–43. [CrossRef]
- Liew, W.-P.-P.; Mohd-Redzwan, S. Mycotoxin: Its impact on gut health and microbiota. Front. Cell Infect. Microbiol. 2018, 8, 60. [CrossRef]
- Yin, H.; Jiang, M.; Peng, X.; Cui, H.; Zhou, Y.; He, M.; Zuo, Z.; Ouyang, P.; Fan, J.; Fang, J. The molecular mechanism of g2m cell cycle arrest induced by afb1 in the jejunum. Oncotarget 2016, 7, 35592–35606. [CrossRef]
- Peng, X.; Chen, K.; Chen, J.; Fang, J.; Cui, H.; Zuo, Z.; Deng, J.; Chen, Z.; Geng, Y.; Lai, W. Aflatoxin b1 a ects apoptosis and expression of bax, bcl-2, and caspase-3 in thymus and bursa of fabricius in broiler chickens. Environ. Toxicol. 2016, 31, 1113–1120. [CrossRef]
- Yang, X.; Liu, L.; Chen, J.; Xiao, A. Response of intestinal bacterial flora to the long-term feeding of aflatoxin b1 (afb1) in mice. Toxins 2017, 9, 317. [CrossRef]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [CrossRef]
- Romero, A.; Ares, I.; Ramos, E.; Castellano, V.; Martínez, M.; Martínez-Larrañaga, M.R.; Anadón, A.; Martínez, M.A. Mycotoxins modify the barrier function of caco-2 cells through di erential gene expression of specific claudin isoforms: Protective e ect of illite mineral clay. Toxicology 2016, 353–354, 21–33. [CrossRef]
- Kohan, A.B.; Yoder, S.M.; Tso, P. Using the lymphatics to study nutrient absorption and the secretion of gastrointestinal hormones. Physiol. Behav. 2011, 105, 82–88. [CrossRef] [PubMed]
- Alexander, J.S.; Ganta, V.C.; Jordan, P.A.; Witte, M.H. Gastrointestinal lymphatics in health and disease. Pathophysiology 2010, 17, 315–335. [CrossRef]
- Worbs, T.; Bode, U.; Yan, S.; Ho mann, M.W.; Hintzen, G.; Bernhardt, G.; Förster, R.; Pabst, O. Oral tolerance originates in the intestinal immune system and relies on antigen carriage by dendritic cells. J. Exp. Med. 2006, 203, 519–527. [CrossRef] [PubMed]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [CrossRef] [PubMed]
- Meissonnier, G.; Pinton, P.; La tte, J.; Cossalter, A.-M.; Gong, Y.Y.; Wild, C.; Bertin, G.; Galtier, P.; Oswald, I. Immunotoxicity of aflatoxin b1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmacol. 2008, 231, 142–149. [CrossRef] [PubMed]
- Turcu, R.; Margareta, O.; Criste, R.D.; Mariana, R.; Tatiana, P.; S,oica, C.; Dragotoiu, D. The e ect of using grape seeds meal as natural antioxidant in broiler diets enriched in fatty acids, on meat quality. J. Hyg. Eng. Des. 2019, 35, 14–20.
- Chedea, V.S.; Palade, L.M.; Marin, D.E.; Pelmus, R.S.; Habeanu, M.; Rotar, M.C.; Gras, M.A.; Pistol, G.C.; Taranu, I. Intestinal absorption and antioxidant activity of grape pomace polyphenols. Nutrients 2018, 10, 588. [CrossRef]
- Veskoukis, A.S.; Kyparos, A.; Nikolaidis, M.G.; Stagos, D.; Aligiannis, N.; Halabalaki, M.; Chronis, K.; Goutzourelas, N.; Skaltsounis, L.; Kouretas, D. The antioxidant e ects of a polyphenol-rich grape pomace extract in vitro do not correspond in vivo using exercise as an oxidant stimulus. Oxid. Med. Cell Longev. 2012, 2012, 185867. [CrossRef]
- Taranu, I.; Habeanu, M.; Gras, M.A.; Pistol, G.C.; Lefter, N.; Palade, M.; Ropota, M.; Sanda Chedea, V.; Marin, D.E. Assessment of the e ect of grape seed cake inclusion in the diet of healthy fattening-finishing pigs. J. Anim. Physiol. Anim. Nutr. 2018, 102, e30–e42. [CrossRef]
- Abdel-Wahhab, M.A.; El-Nekeety, A.A.; Hathout, A.S.; Salman, A.S.; Abdel-Aziem, S.H.; Sabry, B.A.; Hassan, N.S.; Abdel-Aziz, M.S.; Aly, S.E.; Jaswir, I. Bioactive compounds from aspergillus niger extract enhance the antioxidant activity and prevent the genotoxicity in aflatoxin b1-treated rats. Toxicon 2020, 181, 57–68. [CrossRef]
- Oskoueian, E.; Abdullah, N.; Zulkifli, I.; Ebrahimi, M.; Karimi, E.; Goh, Y.M.; Oskoueian, A.; Shakeri, M. Cytoprotective e ect of palm kernel cake phenolics against aflatoxin b1-induced cell damage and its underlying mechanism of action. BMC Complement. Altern. Med. 2015, 15, 392. [CrossRef] [PubMed]
- Lu, H.; Liu, F.; Zhu, Q.; Zhang, M.; Li, T.; Chen, J.; Huang, Y.; Wang, X.; Sheng, J. Aflatoxin b1 can be complexed with oxidised tea polyphenols and the absorption of the complexed aflatoxin b1 is inhibited in rats. J. Sci. Food Agric. 2017, 97, 1910–1915. [CrossRef] [PubMed]
- Ali Rajput, S.; Sun, L.; Zhang, N.; Mohamed Khalil, M.; Gao, X.; Ling, Z.; Zhu, L.; Khan, F.A.; Zhang, J.; Qi, D. Ameliorative e ects of grape seed proanthocyanidin extract on growth performance, immune function, antioxidant capacity, biochemical constituents, liver histopathology and aflatoxin residues in broilers exposed to aflatoxin b1. Toxins 2017, 9, 371. [CrossRef] [PubMed]
- Taranu, I.; Hermenean, A.; Bulgaru, C.; Pistol, G.C.; Ciceu, A.; Grosu, I.A.; Marin, D.E. Diet containing grape seed meal by-product counteracts afb1 toxicity in liver of pig after weaning. Ecotoxicol. Environ. Saf. 2020, 203, 110899. [CrossRef]
- Grosu, I.A.; Pistol, G.C.; Taranu, I.; Marin, D.E. The impact of dietary grape seed meal on healthy and aflatoxin b1 a icted microbiota of pigs after weaning. Toxins 2019, 11, 25. [CrossRef]
- Whitlow, L. Evaluation of mycotoxin binders. In Proceedings of the 4th Mid-Atlantic Nutrition Conference, Timonium, MD, USA, 29–30 March 2006.
- Avantaggiato, G.; Solfrizzo, M.; Visconti, A. Recent advances on the use of adsorbent materials for detoxification of fusarium mycotoxins. Food Addit. Contam. 2005, 22, 379–388. [CrossRef]
- Sabater-Vilar, M.; Malekinejad, H.; Selman, M.H.; van der Doelen, M.A.; Fink-Gremmels, J. In vitro assessment of adsorbents aiming to prevent deoxynivalenol and zearalenone mycotoxicoses. Mycopathologia 2007, 163, 81–90. [CrossRef]
- Bo?carov-Stan?ci´c, A.; Adamovi´c, M.; Salma, N.; Bodroža-Solarov, M.; Vu?ckovi´-Ðisalov, J.; Panti´c, V. In vitro e cacy of mycotoxins adsorption by natural mineral adsorbents. Biotechnol. Anim. Husb. 2011, 27, 1241–1251. [CrossRef]
- Huwig, A.; Freimund, S.; Käppeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by di erent adsorbents. Toxicol. Lett. 2001, 122, 179–188. [CrossRef]
- Di Gregorio, M.C.; Nee , D.V.D.; Jager, A.V.; Corassin, C.H.; Carão, Á.C.D.P.; Albuquerque, R.D.; Azevedo, A.C.D.; Oliveira, C.A.F. Mineral adsorbents for prevention of mycotoxins in animal feeds. Toxin Rev. 2014, 33, 125–135. [CrossRef]
- Teixeira, A.; Baenas, N.; Dominguez-Perles, R.; Barros, A.; Rosa, E.; Moreno, D.A.; Garcia-Viguera, C. Natural bioactive compounds from winery by-products as health promoters: A review. Int. J. Mol. Sci. 2014, 15, 15638–15678. [CrossRef] [PubMed]
- Tang, G.-Y.; Zhao, C.-N.; Liu, Q.; Feng, X.-L.; Xu, X.-Y.; Cao, S.-Y.; Meng, X.; Li, S.; Gan, R.-Y.; Li, H.-B. Potential of grape wastes as a natural source of bioactive compounds. Molecules 2018, 23, 2598. [CrossRef] [PubMed]
- Chedea, V.S.; Pelmus, R.S.; Lazar, C.; Pistol, G.C.; Calin, L.G.; Toma, S.M.; Dragomir, C.; Taranu, I. E ects of a diet containing dried grape pomace on blood metabolites and milk composition of dairy cows. J. Sci. Food Agric. 2017, 97, 2516–2523. [CrossRef] [PubMed]
- Avantaggiato, G.; Greco, D.; Damascelli, A.; Solfrizzo, M.; Visconti, A. Assessment of multi-mycotoxin adsorption e cacy of grape pomace. J. Agric. Food Chem. 2014, 62, 497–507. [CrossRef] [PubMed]
- Gambacorta, L.; Pinton, P.; Avantaggiato, G.; Oswald, I.P.; Solfrizzo, M. Grape pomace, an agricultural byproduct reducing mycotoxin absorption: In vivo assessment in pig using urinary biomarkers. J. Agric. Food Chem. 2016, 64, 6762–6771. [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I. Overview on aflatoxins and oxidative stress. Toxin Rev. 2012, 31, 32–43. [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative stress and inflammation: What polyphenols can do for us? Oxid. Med. Cell Longev. 2016, 2016, 7432797. [CrossRef]
- Marin, D.E.; Pistol, G.C.; Gras, M.; Palade, M.; Taranu, I. A comparison between the e ects of ochratoxin a and aristolochic acid on the inflammation and oxidative stress in the liver and kidney of weanling piglets. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 1147–1156. [CrossRef]
- Marin, D.E.; Pistol, G.C.; Gras, M.A.; Palade, M.L.; Taranu, I. Comparative e ect of ochratoxin a on inflammation and oxidative stress parameters in gut and kidney of piglets. Regul. Toxicol. Pharmacol. RTP 2017, 89, 224–231. [CrossRef]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin b(1) in a ecting broiler’s performance, immunity, and gastrointestinal tract: A review of history and contemporary issues. Toxins 2011, 3, 566–590. [CrossRef]
- Benkerroum, N. Chronic and acute toxicities of aflatoxins: Mechanisms of action. Int. J. Environ. Res. Public Health 2020, 17, 423. [CrossRef]
- Mehrzad, J.; Malvandi, A.M.; Alipour, M.; Hosseinkhani, S. Environmentally relevant level of aflatoxin b(1) elicits toxic pro-inflammatory response in murine cns-derived cells. Toxicol. Lett. 2017, 279, 96–106. [CrossRef]
- Sangiovanni, E.; Dell’Agli, M. Special issue: Anti-inflammatory activity of plant polyphenols. Biomedicines 2020, 8, 64. [CrossRef]
- Calder, P.C. Immunoregulatory and anti-inflammatory e ects of n-3 polyunsaturated fatty acids. Braz. J. Med. Biol. Res. 1998, 31, 467–490. [CrossRef] [PubMed]
- Holanda, D.M.; Kim, S.W. E cacy of mycotoxin detoxifiers on health and growth of newly-weaned pigs under chronic dietary challenge of deoxynivalenol. Toxins 2020, 12, 311. [CrossRef] [PubMed]
- Van Le Thanh, B.; Lessard, M.; Chorfi, Y.; Guay, F. The e cacy of anti-mycotoxin feed additives in preventing the adverse e ects of wheat naturally contaminated with fusarium mycotoxins on performance, intestinal barrier function and nutrient digestibility and retention in weanling pigs. Can. J. Anim. Sci. 2015, 95, 197–209. [CrossRef]
- Solis-Cruz, B.; Hernandez-Patlan, D.; Petrone, V.M.; Pontin, K.P.; Latorre, J.D.; Beyssac, E.; Hernandez-Velasco, X.; Merino-Guzman, R.; Owens, C.; Hargis, B.M.; et al. Evaluation of cellulosic polymers and curcumin to reduce aflatoxin b1 toxic e ects on performance, biochemical, and immunological parameters of broiler chickens. Toxins 2019, 11, 121. [CrossRef]
- Xu, F.; Wang, P.; Yao, Q.; Shao, B.; Yu, H.; Yu, K.; Li, Y. Lycopene alleviates afb(1)-induced immunosuppression by inhibiting oxidative stress and apoptosis in the spleen of mice. Food Funct. 2019, 10, 3868–3879. [CrossRef]
- Taranu, I.; Marin, D.E.; Palade, M.; Pistol, G.C.; Chedea, V.S.; Gras, M.A.; Rotar, C. Assessment of the e cacy of a grape seed waste in counteracting the changes induced by aflatoxin b1 contaminated diet on performance, plasma, liver and intestinal tissues of pigs after weaning. Toxicon O . J. Int. Soc. Toxinol. 2019, 162, 24–31. [CrossRef]
- Taranu, I.; Gras, M.; Pistol, G.C.; Motiu, M.; Marin, D.E.; Lefter, N.; Ropota, M.; Habeanu, M. Omega-3 pufa rich camelina oil by-products improve the systemic metabolism and spleen cell functions in fattening pigs. PLoS ONE 2014, 9, e110186. [CrossRef]
- Palade, L.M.; Habeanu, M.; Marin, D.E.; Chedea, V.S.; Pistol, G.C.; Grosu, I.A.; Gheorghe, A.; Ropota, M.; Taranu, I. E ect of dietary hemp seed on oxidative status in sows during late gestation and lactation and their o spring. Anim. Open Access J. 2019, 9, 194. [CrossRef]
- Pistol, G.C.; Braicu, C.; Motiu, M.; Gras, M.A.; Marin, D.E.; Stancu, M.; Calin, L.; Israel-Roming, F.; Berindan-Neagoe, I.; Taranu, I. Zearalenone mycotoxin a ects immune mediators, mapk signalling molecules, nuclear receptors and genome-wide gene expression in pig spleen. PLoS ONE 2015, 10, e0127503. [CrossRef] [PubMed]
- Marin, D.E.; Pistol, G.C.; Neagoe, I.V.; Calin, L.; Taranu, I. E ects of zearalenone on oxidative stress and inflammation in weanling piglets. Food Chem. Toxicol. 2013, 58, 408–415. [CrossRef] [PubMed]
- Pistol, G.C.; Gras, M.A.; Marin, D.E.; Israel-Roming, F.; Stancu, M.; Taranu, I. Natural feed contaminant zearalenone decreases the expressions of important pro- and anti-inflammatory mediators and mitogen-activated protein kinase/nf-kappab signalling molecules in pigs. Br. J. Nutr. 2014, 111, 452–464. [CrossRef] [PubMed]
- Marin, D.E.; Motiu, M.; Pistol, G.C.; Gras, M.A.; Israel-Roming, F.; Calin, L.; Stancu, M.; Taranu, I. Diet contaminated with ochratoxin a at the highest level allowed by eu recommendation disturbs liver metabolism in weaned piglets. World Mycotoxin J. 2016, 9, 587–596. [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [CrossRef]