Introduction
The use of enzyme complex in animal feeding improves the utilization of nutrients that would otherwise be unavailable (Leite et al., 2011), reflecting positively on the performance and reducing the pollution load of excreta (Opalinski et al., 2010). However, the inclusion of enzymes in formulations should be made carefully, because this additive may lose their biological activity during pelleting process, which involves high temperatures, moisture, and pressure.
The enzymes used in animal feed should resist and maintain considerable activity after the manufacturing and digestion processes to justify their inclusion in the diets (Dias et al., 2002; Shah and Madamwar, 2005). The factors that may influence their stability, among others, are the origin (organism), the type of enzyme, the diet composition, the processing conditions (temperature, pressure, and moisture), the storage, the conditions during the digestion process, and the action of endogenous enzymes (Ramos et al., 2012).
Today, most enzymes used in animal diets are produced by fungi; however, they can be readily denatured at temperatures above 60 °C. The enzyme stability is superior when incorporated with other ingredients of the formulation, which reduces the possibility of inactivation (Francesch et al., 1995). Therefore, an ideal enzyme must be able to withstand temperatures above 70 °C, since the processing of pelleted and extruded diets uses temperatures higher than this (Finnfeeds International, 1991). Moreover, enzymes must maintain their biological activity over time. According to Cowan (1993), the enzymes can be stable for more than three months at 25 °C when blended with other ingredients.
Considering the options of enzyme complexes, the complex solid-state fermentation (SSF) seems to stand out due to its qualitative features. Besides being naturally produced by fungi of the genus Aspergillus on solid matrix (Vats and Banerjee, 2004), SSF may have dozens of enzymes that work in synergy (carbohydrases, proteases, and others), providing more nutrients, which may improve performance of animals (Moura et al., 2015). However, studies on the stability of enzymes present in this complex should be performed, because this is subjected to various physical and chemical factors of the diet during the processing, storage, and digestive processes, which can reduce or inactivate its catalytic activity.
The effects of processing and storage time on the stability of enzymes of the enzyme complex SSF in pelleted diets for animals were evaluated.
Material and Methods
The trial was conducted in Viçosa, Minas Gerais, Brazil.
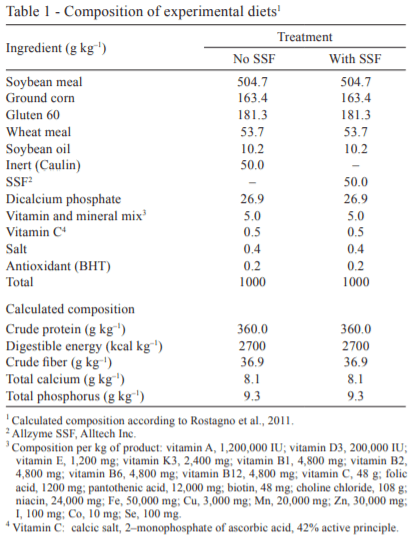
Two isonutritive diets were formulated containing 360 g kg–1 of crude protein (CP) and 2,700 kcal kg–1 of digestible energy (DE) in the diet (Table 1). This diet was formulated according to the requirements recommended for omnivore fish, but the type of processing and the enzyme activity can be applied to any animal species. The control diet was formulated without SSF and 50 g kg–1 of enzyme complex was added to the other experimental diet.
The ingredients were weighed and placed in a plastic bag for mixing. These bags were shaken for 5 min, providing a homogeneous mixture of ingredients. This mixture was placed in a bowl and water (at 55 °C) was added until the dough reached the point alloy. Soon after, the pellet machine received this dough and the wet pellets were produced. These pellets remained in forced ventilation oven at 55 °C for 14 h, which promoted its drying.
The trial was started at the moment of the processing of experimental diets and the samples were collected during the following steps: mixing, then pelleting, and then drying in an oven at 55 °C for 14 h. To evaluate the storage time, the diet ready after drying was regarded as day 1. On this day, two samples were taken, one kept at room temperature at 25 °C and one kept in a freezer at –18 °C.
At 15, 30, 45, and 60 days, sub-samples were taken to the two kinds of storage. All samples, the processing steps, and storage time were submitted to the laboratory and the activity of the following enzymes were measured: α-galactosidase, endoglucanase (carboxymethyl cellulase), xylanase, sucrase (invertase), α-amylase, lipase, and Trypsin.
To assess α-galactosidase, endoglucanase, xylanase, and sucrase, 200 mg of each sample of feed were macerated in 10 mL of buffer solution of 100 mM sodium acetate. This mixture was centrifuged at 13,000 rpm for 2 min and the supernatant (extract) was removed and stored in a freezer at –18 °C for enzyme analysis.
The activity of α-galactosidase was determined by measuring the amount of reducing sugar produced through the use of dinitrosalicylic acid (DNS) reagent according to Miller (1956). The reaction mixture was composed of 325 µL sodium acetate buffer (100 mM, pH 5), 125 µL of a solution of 10 mM sucrose, and 50 µL of enzyme extract. The trial was conducted in a water bath for 15 min at 40 o C. To stop the reaction, 1 mL of DNS reagent was added, followed by immersion of the test tube in boiling water bath for 5 min. The spectrophotometric measurements were taken at 540 nm and absorbance values converted into µmoles of reducing sugar, using a standard curve constructed from glucose amounts ranging from 0.2 to 2 µmoles.
For endoglucanase assay, 30 µL of the enzyme solution was mixed with 400 µL of carboxymethyl cellulose (0.625% w/v) and diluted in 70 µL of sodium acetate buffer (100 mM, pH 5). This solution was placed in a water bath at 50 °C for 30 min and the reaction stalled according to Miller (1956), using 500 µL of DNS and boiling water bath for 5 min. Afterwards, the spectrophotometric measurements were made at 540 nm.
The xylanase activity was determined using 70 µL sodium acetate buffer (100 mM, pH 5), 30 µL of enzyme extract, and 400 µL of birch wood xylan solution (1.25% w/v). The reaction was conducted for 20 min at 40 °C and paralyzed with 500 µL of DNS. This solution was incubated in boiling water bath for 5 min for color development. The activity was determined at 540 nm using a standard glucose curve.
For sucrase activity, 15 µL of enzyme extract was added to 125 µL of sucrase solution (2 g sucrose for analysis in 50 mL of sodium acetate buffer, 100 mM, pH 5) and 360 µL sodium acetate buffer solution (100 mM, pH 5). This solution was immediately placed in a water bath at 30 °C for 30 min and the reaction terminated with 500 µL DNS. Subsequently, the sample was taken to a bath in boiling water for 5 min and cooled in ambient temperature. Then, the activities were read in a spectrophotometer at 540 nm.
For α-amylase, trypsin, and lipase, 0.5 g of diet sample was macerated with 10 mL of 0.1M Tris-HCl buffer solution. This material was placed into polyethylene tubes and centrifuged at 12,000 rpm for 10 min. Thus, the supernatant was removed for determination of enzyme activity.
The α-amylase activity was based in the starch hydrolysis with release of dextrin and maltose molecules. By adding iodine, unhydrolysed starch acquires blue color. The amylase activity is inversely proportional to the intensity of blue color and is calculated by comparison with a control substrate. The α-amylase activity was determined in spectrophotometer at 660 nm wavelength, using the amylase of Bioclin colorimetric kit according to Caraway (1959).
The trypsin activity was obtained using N-BenzoylD-p-nitroanilide L-arginine (D, L-BApNA) as substrate according to the method described by Erlanger et al. (1961). Ten µL of enzyme extract was added, and immediately, the initial velocity was obtained by forming the p-nitroalinine. This reaction was determined in absorbance at 410 nm as a function of time. For calculations, the molar extinction coefficient of 8,800 M–1 cm–1 was used for the product.
For lipase activity, we used the Bioclin kit with the modified methodology of Cherry (1932). This methodology evaluated the lipase action present in the extract of the diet on a glycerol ester, releasing a chromogen, which was quantitatively determined in a spectrophotometer at 410 nm. The intensity of color formed was proportional to lipase activity.
All enzyme analyzes were performed with three replicates in duplicate. For statistical analysis we used the SAS software (Statistical Analysis System, version 9.0). The means were compared by SNK (Student Newman Kells) test at 0.05 to the assessment of processing and storage time. Through the F test at 0.05 we evaluated the means of two different forms of storage (25 °C and –18 °C). The results of the activities were also measured in terms of relative enzymatic activity, considering the initial phase of each time at 100%.
Results and Discussion
Enzyme activities were not observed in all the samples of the control treatment. Thus, only the means of enzyme activities of the treatment with SSF subjected to the processing and storage time at different temperatures were demonstrated.
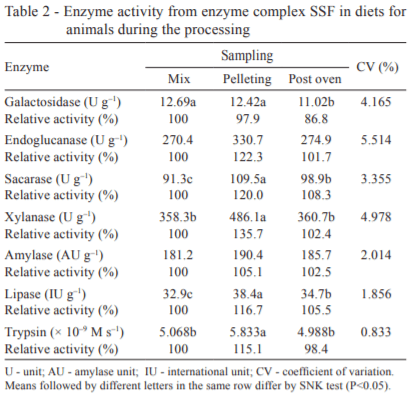
Over the processing (Table 2), the mixture was subjected to the pelletizing temperature of 55 °C and 14 h in forced ventilation oven at the same temperature. In general, the enzyme complex SSF was stable in relation to chemical and physical adversities of the trial. There were no observed differences (P>0.05) between the mixture and the post-oven step for activities of endoglucanase, xylanase, amylase, and trypsin. However, the α-galactosidase activity was significantly reduced (P<0.05) by 13.2%, unlike the sucrase and lipase activities, which increased (P<0.05) in 8.3% and 5.5%, respectively.
The activity results of the enzymes involved in this study were consistent with Spring et al. (1996), who tested the effect of the temperature of the pellet (60 °C to 100 °C) on the activity of cellulase, amylase, and pentosanase in diets with wheat and barley. The authors concluded that the enzyme activity was maintained at a pellet temperature of 80 °C. Close values were obtained by Silveira et al. (2010), who noted that pelleted diet at 75 °C did not cause deleterious effect on the enzyme activity of the complex SSF. Moreover, Colier and Hardy (1986) verified that after the pelleting process of microbial enzymes at 70 °C, the α-amylase activity decreased its original activity by 64.5%.
To all the enzymes, except for the α-galactosidase and α-amylase, the catalytic activity increased after the pelleting process. Probably, the moisture of the pellets favored the enzyme activity, because the presence of water is necessary for the actuation of the enzyme on a substrate, or hydrolysis.
Using the chromogenic substrate BAPNA, Marcushi (2010) also observed that the maximum trypsin activity of tambaqui (Colossoma macropomum) was verified in the pH range from 7.5 to 11.5 and at temperature of 70 °C. As for its heat stability over time, the enzyme retained more than 60% of its initial activity after 3 h at 60 °C. This supports the idea that the trypsin of the present study showed greater activity during pelleting (55 °C), with the values reduced after the drying stage (14 h at 55 °C).
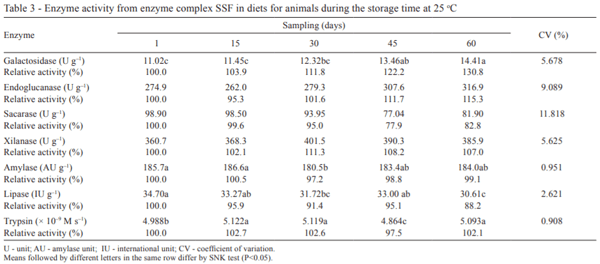
For the storage period, differences were not observed (P>0.05) for the activities of endoglucanase, sucrase, xylanase, amylase, and trypsin (Table 3). The experimental time did not influence the activities, suggesting that these enzymes remained stable during 60 days post oven. Similarly, Bedford (1996) also observed that an enzyme complex formed by amylase, protease, and xylanase withstood at 85 °C and 90 °C during 15 and 2 min, respectively, when subjected to the pelleting process. When stored at 22 °C, the enzyme activity was maintained for 12 months.
As for the other enzymes, differences were verified (P<0.05) during the storage time. The α-galactosidase and trypsin activities increased 30.8% and 2.1%, respectively, while for lipase activity, a reduction of 11.8% was observed. From 15 to 60 days of storage, the endoglucanase, sucrase, xylanase, and amylase remained stable at 25 °C, because no loss of activity was observed (P>0.05) over time.
Among the enzymes, the α-galactosidase presented a different standard of catalytic activity compared with other enzymes. After 14 h of diet drying in an oven at 55 °C, this enzyme may have suffered a partial denaturation, which reduced the activity. After a few days, the activity increased, evidencing a gradual enzyme renaturation. According to Nelson and Cox (2011), proteins (such as enzymes) can undergo a mild denaturation, with possible subsequent renaturation. Many proteins denatured by heat and other factors may regain their native structure and biological activity if returned to the previous conditions.
For trypsin, although similar activity among 15, 30, and 60 days, there was a reduction at 45 days, but still close to 100% of the initial relative activity. Under the conditions of this study, the trypsin remained near 100% relative activity throughout the trial despite some differences between treatments. In absolute values, the trypsin activity in the mixture and 60 days of storage were near (5.068 × 10–9 M s–1 and 5.093 × 10–9 M s–1, respectively).
Simulating the process of pelletizing, Ferreira et al. (2005) evaluated the lipase activity subjected to various temperatures and the presence of metallic and mineral ions in in vitro form. The authors concluded that there was loss of 16.13% activity when the lipase was subjected to 80 °C for 15 min. Regarding mineral solutions, all minerals and metal ions (Ca, Cu, Co, Mn, and Fe) strongly inhibited the lipase activity in less than 42%. The influence of these minerals on the lipase may explain the loss of activity in this study, because the diet was formulated with mineral supplementation, which contains all mineral and metal ions described above.
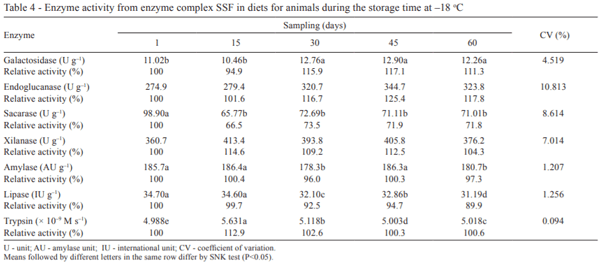
For the storage temperature of –18 °C (Table 4) only, no difference was observed (P>0.05) for endoglucanase and xylanase activities. The catalytic behavior of the enzymes during the storage time at 25 °C were similar to enzymes subjected to –18 °C, except for the sucrase. Normally, the enzyme activity is maintained when stored at freezing temperatures, which did not occur with this enzyme. In this case, the temperature negatively influenced (P<0.05) the sucrase activity with losses of 28.2% between the 1st and the 60th day post oven. Thus, for diets with enzyme complex SSF and high levels of sucrose, the freezing temperature should be prevented to a greater hydrolysis of the disaccharide.
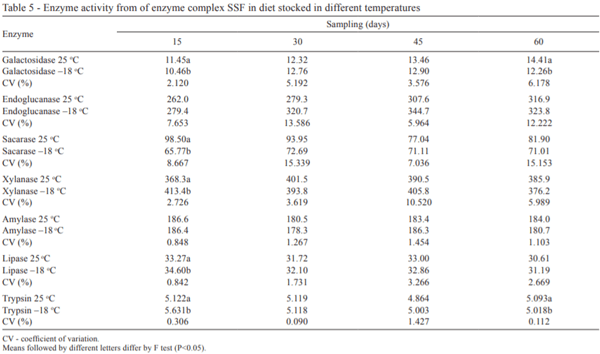
The two types of storage (25 °C and –18 °C) were compared and difference was only observed (P<0.05) for galactosidase and trypsin activities at 60 days (Table 5). At temperature of 25 °C, these enzymes showed higher activity than those subjected to temperature of –18 °C, however, both superior than the initial activity. For all enzymes from complex SSF, no significant difference (P>0.05) between 30 and 45 days was verified.
This result indicated that pelleted diet containing enzyme complex SSF does not need to be kept in negative temperatures up to 60 days, because the form of protection for this product provides stability to the processing and storage. This is advantageous for companies and producers, since it reduces the costs for obtaining cold storage and energy.
Conclusions
The enzymes evaluated from the enzyme complex SSF do not lose the stability when subjected to processing of pelleted diet at 55 °C, maintaining the activity for at least 60 days when stored in temperature at up to 25 °C.
Diets containing the enzyme complex SSF do not need to be maintained at negative temperatures down to –18 °C.
The enzyme complex SSF should be evaluated at higher processing temperatures and storage time greater than 60 days.
Acknowledgments
We would like to thank CNPq, CAPES, FAPEMIG, and Alltech Inc. for financial support.
This article was originally published in Revista Brasileira de Zootecnia, 45(12):731-736, 2016. http://dx.doi.org/10.1590/S1806-92902016001200001. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/).










")